Abundance and Spatial Distribution of Aerobic Anoxygenic Phototrophic Bacteria in Tama River, Japan


Abstract
:1. Introduction
2. Materials and Methods
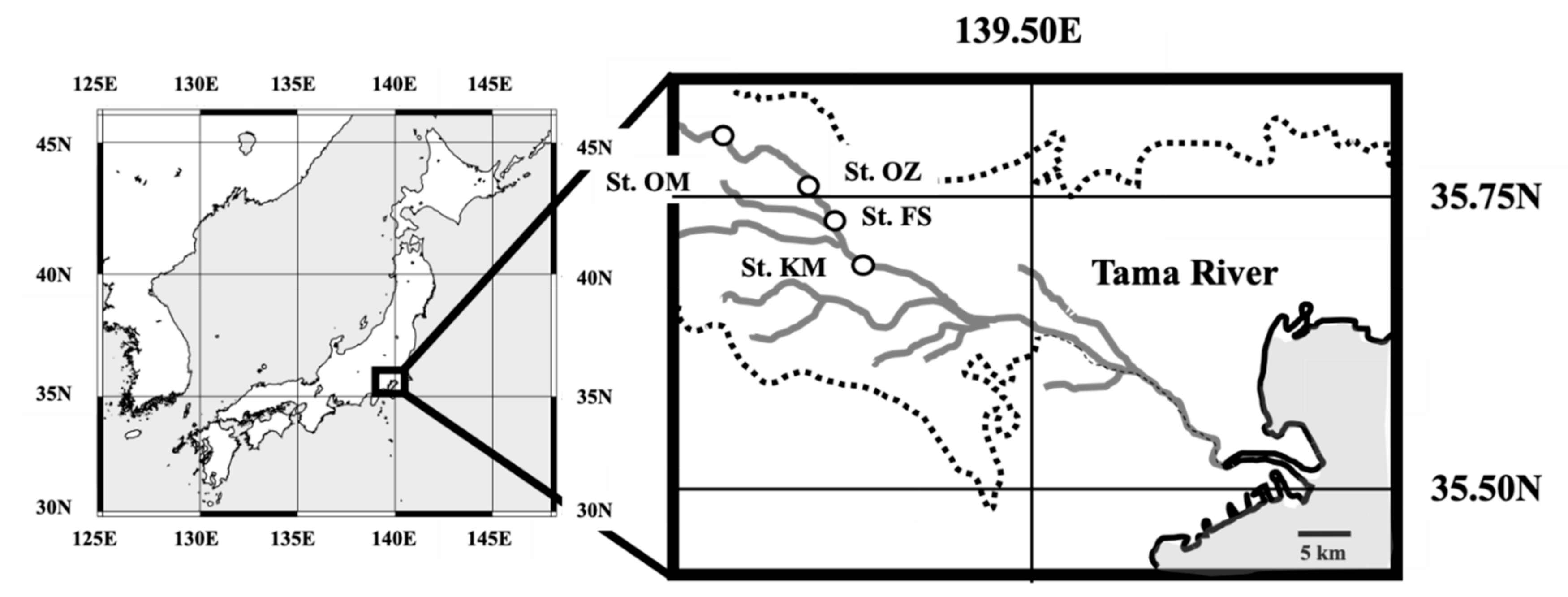
2.1. Sampling Sites and the Reported Geochemical Parameters
2.2. Infra-Red Epifluorescence Microscopy
cyanobacterial cells appearing on the Chl a filter set)/DAPI-stained bacterial cells per unit area.
3. Results and Discussion
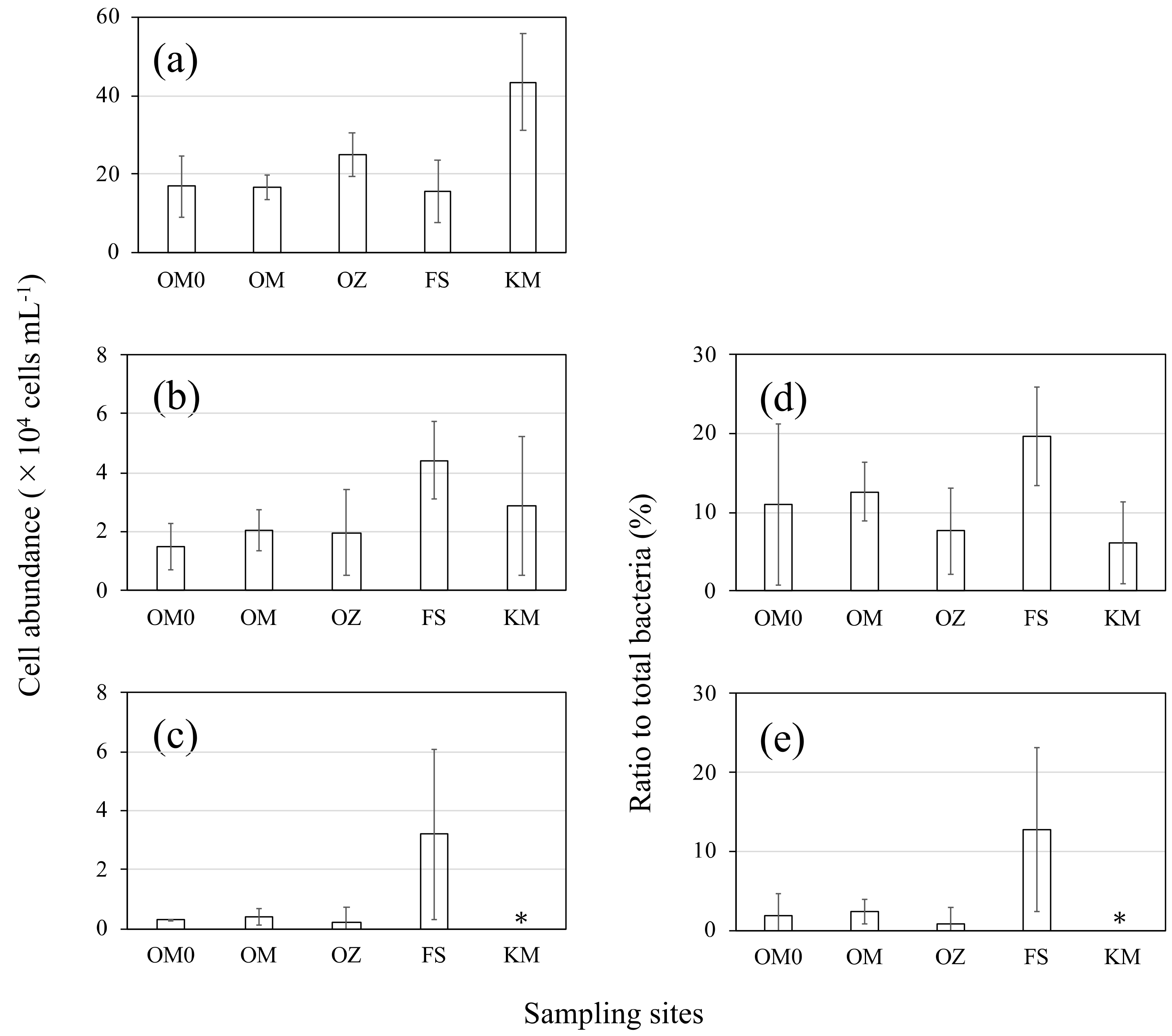
3.1. Determination of Absolute Cell Abundances of the Total Bacteria, Cyanobacteria and AAnPB
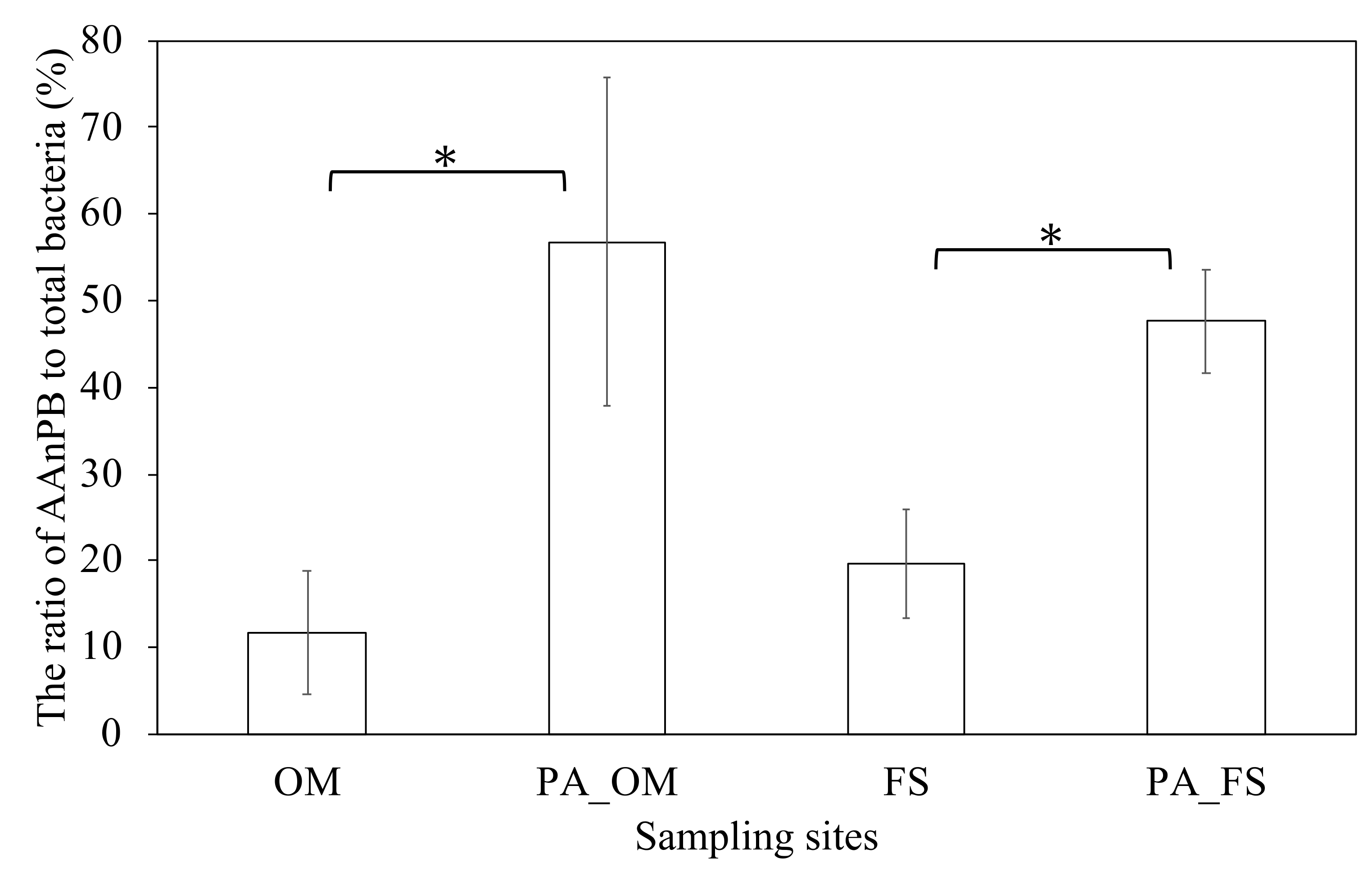
3.2. Observation of the Particle-Attached AAnPB and Their Distribution Comparison with Free-Living AAnPB
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beatty, J.T. On the natural selection and evolution of the aerobic phototrophic bacteria. Photosynth. Res. 2002, 73, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Kolber, Z.S.; Van Dover, C.L.; Niderman, R.A.; Falkowski, P.G. Bacterial photosynthesis in surface waters of the open ocean. Nature 2000, 407, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Kolber, Z.S.; Plumley, F.G.; ALang, A.S.; Beatty, J.T.; Blankenship, R.E.; Van Dover, C.L.; Vetriani, C.; Koblížek, M.; Rathgeber, C.; Falkowski, P.G. Contribution of Aerobic Photoheterotrophic Bacteria to the Carbon Cycle in the Ocean. Science 2001, 292, 2492–2495. [Google Scholar] [CrossRef] [PubMed]
- Koblížek, M. Ecology of aerobic anoxygenic phototrophs in aquatic environments. FEMS Microbiol. Rev. 2015, 39, 854–870. [Google Scholar] [CrossRef] [Green Version]
- Lami, R.; Cottrell, M.T.; Ras, J.; Ulloa, O.; Obernosterer, I.; Claustre, H.; Kirchman, D.L.; LeBaron, P. High Abundances of Aerobic Anoxygenic Photosynthetic Bacteria in the South Pacific Ocean. Appl. Environ. Microbiol. 2007, 73, 4198–4205. [Google Scholar] [CrossRef] [Green Version]
- Cottrell, M.T.; Kirchman, D.L. Photoheterotrophic Microbes in the Arctic Ocean in Summer and Winter. Appl. Environ. Microbiol. 2009, 75, 4958–4966. [Google Scholar] [CrossRef] [Green Version]
- Sato-Takabe, Y.; Nakao, H.; Kataoka, T.; Yokokawa, T.; Hamasaki, K.; Ohta, K.; Suzuki, S. Abundance of Common Aerobic Anoxygenic Phototrophic Bacteria in a Coastal Aquaculture Area. Front. Microbiol. 2016, 7, 1996. [Google Scholar] [CrossRef] [Green Version]
- Mašín, M.; Nedoma, J.; Pechar, L.; Koblížek, M. Distribution of aerobic anoxygenic phototrophs in temperate freshwater systems. Environ. Microbiol. 2008, 10, 1988–1996. [Google Scholar] [CrossRef]
- Medová, H.; Boldareva, E.N.; Hrouzek, P.; Borzenko, S.V.; Namsaraev, Z.B.; Gorlenko, V.M.; Namsaraev, B.B.; Koblížek, M. High abundances of aerobic anoxygenic phototrophs in saline steppe lakes. FEMS Microbiol. Ecol. 2011, 76, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Čuperová, Z.; Holzer, E.; Salka, I.; Sommaruga, R.; Koblížek, M. Temporal Changes and Altitudinal Distribution of Aerobic Anoxygenic Phototrophs in Mountain Lakes. Appl. Environ. Microbiol. 2013, 79, 6439–6446. [Google Scholar] [CrossRef] [Green Version]
- Fauteux, L.; Cottrell, M.T.; Kirchman, D.L.; Borrego, C.M.; Garcia-Chaves, M.C.; Del Giorgio, P.A. Patterns in Abundance, Cell Size and Pigment Content of Aerobic Anoxygenic Phototrophic Bacteria along Environmental Gradients in Northern Lakes. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Kasalický, V.; Zeng, Y.; Piwosz, K.; Šimek, K.; Kratochvilová, H.; Koblížek, M. Aerobic Anoxygenic Photosynthesis Is Commonly Present within the Genus Limnohabitans. Appl. Environ. Microbiol. 2017, 84, e02116-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Song, A.; Peng, W.; Jin, Z.; Müller, W.E.G.; Wang, X. Contribution of aerobic anoxygenic phototrophic bacteria to total organic carbon pool in aquatic system of subtropical karst catchments, Southwest China: Evidence from hydrochemical and microbiological study. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Ferrera, I.; Sarmento, H.; Priscu, J.C.; Chiuchiolo, A.; González, J.M.; Grossart, H.-P. Diversity and Distribution of Freshwater Aerobic Anoxygenic Phototrophic Bacteria across a Wide Latitudinal Gradient. Front. Microbiol. 2017, 8, 175. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Wu, X.; Zhou, Q.; Donde, O.; Tian, C.; Wang, C.; Feng, B.; Xiao, B. Distribution of Aerobic Anoxygenic Phototrophs in Freshwater Plateau Lakes. Pol. J. Environ. Stud. 2018, 27, 871–879. [Google Scholar] [CrossRef]
- Lami, R.; Âuperová, Z.; Ras, J.; LeBaron, P.; Koblíïek, M. Distribution of free-living and particle-attached aerobic anoxygenic phototrophic bacteria in marine environments. Aquat. Microb. Ecol. 2009, 55, 31–38. [Google Scholar] [CrossRef]
- Waidner, L.A.; Kirchman, D.L. Aerobic anoxygenic phototrophic bacteria attached to particles in tubid waters of the Delaware and Chesapeake estuaries. Appl. Environ. Microbiol. 2007, 73, 3936–3944. [Google Scholar] [CrossRef] [Green Version]
- Okai, M.; Aoki, H.; Ishida, M.; Urano, N. Antibiotic-resistance of fecao coliforms at the bottom of the Tama River, Tokyo. Biocontrol Sci. 2019, 24, 173–178. [Google Scholar] [CrossRef]
- Kirchman, D.L. Introduction and overview. In Microbial Ecology of the Ocean, 2nd ed.; Kirchman, D.L., Ed.; Wiley: Hoboken, NJ, USA, 2008; Chapter 1; pp. 1–6. [Google Scholar]
- Sieracki, M.E.; Gilg, I.C.; Thier, E.C.; Poulton, N.J.; Goericke, R. Distribution of planktonic aerobic anoxygenic photoheterotrophic bacteria in the northwest Atlantic. Limnol. Oceanogr. 2006, 51, 38–46. [Google Scholar] [CrossRef]
- Jiao, N.; Zhang, Y.; Zeng, Y.; Hong, N.; Liu, R.; Chen, F.; Wang, P. Distinct distribution pattern of abundance and diversity of aerobic anoxygenic phototrophic bacteria in the global ocean. Environ. Microbiol. 2007, 9, 3091–3099. [Google Scholar] [CrossRef]
- Hojerová, E.; Mašín, M.; Brunet, C.; Ferrera, I.; Gasol, J.M.; Koblížek, M. Distribution and Growth of Aerobic Anoxygenic Phototrophs in the Mediterranean Sea. Environ. Microbiol. 2011, 13, 2717–2725. [Google Scholar] [CrossRef] [PubMed]
- Mašín, M.; Cepáková, Z.; Hojerová, E.; Salka, I.; Grossart, H.P.; Koblížek, M. Distribution of aerobic anoxygenic phototrophic bacteria in glacial lakes of northern Europe. Aquat. Microb. Ecol. 2012, 66, 77–86. [Google Scholar] [CrossRef]
- Hirose, S.; Matsuura, K.; Haruta, S. Phylogenetically Diverse Aerobic Anoxygenic Phototrophic Bacteria Isolated from Epilithic Biofilms in Tama River, Japan. Microbes Environ. 2016, 31, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiba, T.; Simidu, U.; Taga, N. Distribution of Aerobic Bacteria Which Contain Bacteriochlorophyll a. Appl. Environ. Microbiol. 1979, 38, 43–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-González, C.; Proia, L.; Ferrera, I.; Gasol, J.M.; Sabater, S. Effects of large river dam regulation on bacterioplankton community structure. FEMS Microbiol. Ecol. 2013, 84, 316–331. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organisms | DAPI | Chl a | BChl a + Chl a |
|---|---|---|---|
| Heterotrophs | + | - | - |
| Cyanobacteria | + | + | + |
| AAnPB | + | - | + |
| Site | WT (°C) | pH | DO (mg L−1) | TN (mg L−1) | TP (mg L−1) |
|---|---|---|---|---|---|
| OM0 | 16.8 | 8.32 | 11.0 | 0.48 | 0.009 |
| OM | 14.9 | 9.00 | 10.8 | 0.48 | 0.011 |
| OZ | 17.6 | 9.10 | N.D. | N.D. | N.D. |
| FS | 22.6 | 8.00 | 10.2 | 0.53 | 0.009 |
| KM | 22.9 | 8.50 | N.D. | N.D. | N.D. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato-Takabe, Y.; Hirose, S.; Hori, T.; Hanada, S. Abundance and Spatial Distribution of Aerobic Anoxygenic Phototrophic Bacteria in Tama River, Japan. Water 2020, 12, 150. https://doi.org/10.3390/w12010150
Sato-Takabe Y, Hirose S, Hori T, Hanada S. Abundance and Spatial Distribution of Aerobic Anoxygenic Phototrophic Bacteria in Tama River, Japan. Water. 2020; 12(1):150. https://doi.org/10.3390/w12010150
Chicago/Turabian StyleSato-Takabe, Yuki, Setsuko Hirose, Tomoyuki Hori, and Satoshi Hanada. 2020. "Abundance and Spatial Distribution of Aerobic Anoxygenic Phototrophic Bacteria in Tama River, Japan" Water 12, no. 1: 150. https://doi.org/10.3390/w12010150
APA StyleSato-Takabe, Y., Hirose, S., Hori, T., & Hanada, S. (2020). Abundance and Spatial Distribution of Aerobic Anoxygenic Phototrophic Bacteria in Tama River, Japan. Water, 12(1), 150. https://doi.org/10.3390/w12010150