Effect of Cadmium and Nickel Exposure on Early Development in Zebrafish (Danio rerio) Embryos

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry and Embryo Collection
2.2. Heavy Metal Exposure
2.3. Survival and Developmental Assessments
2.4. Morphometric Measurements
2.5. Immunohistochemical Analysis of Myosin Expression
2.6. Statistical Analyses
3. Results
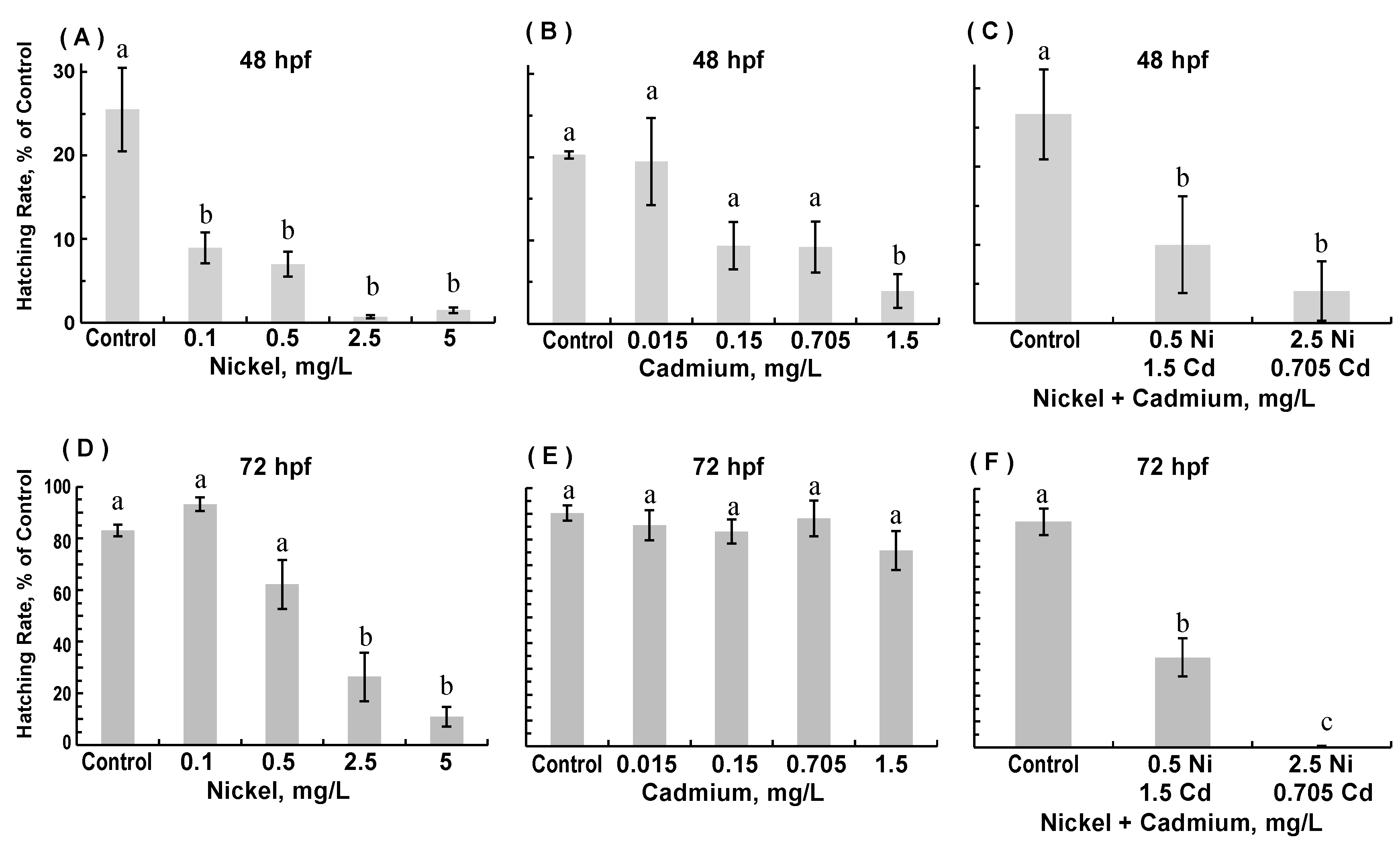
3.1. Survival and Hatching Success
3.2. Spontaneous and Elicited Movements
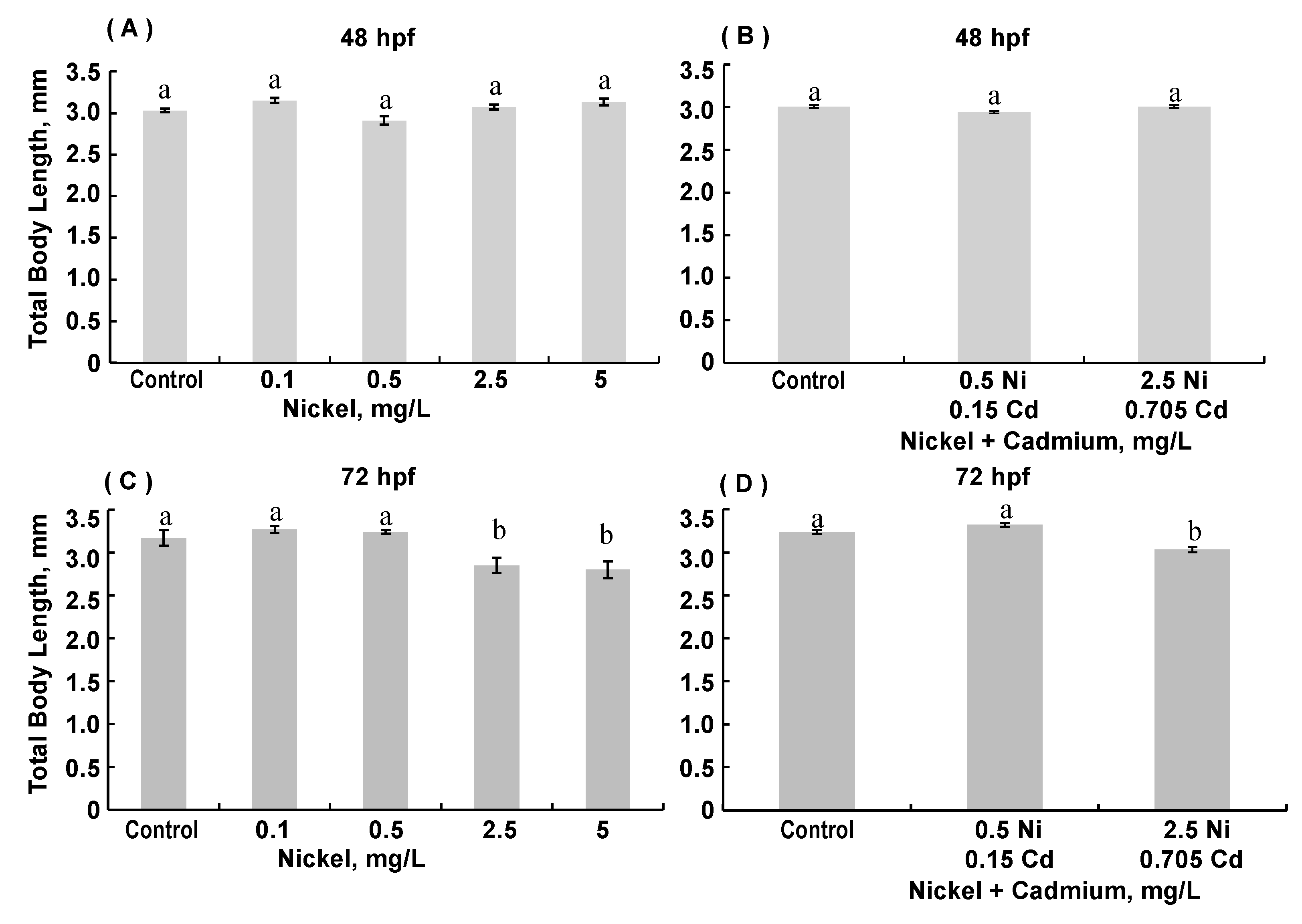
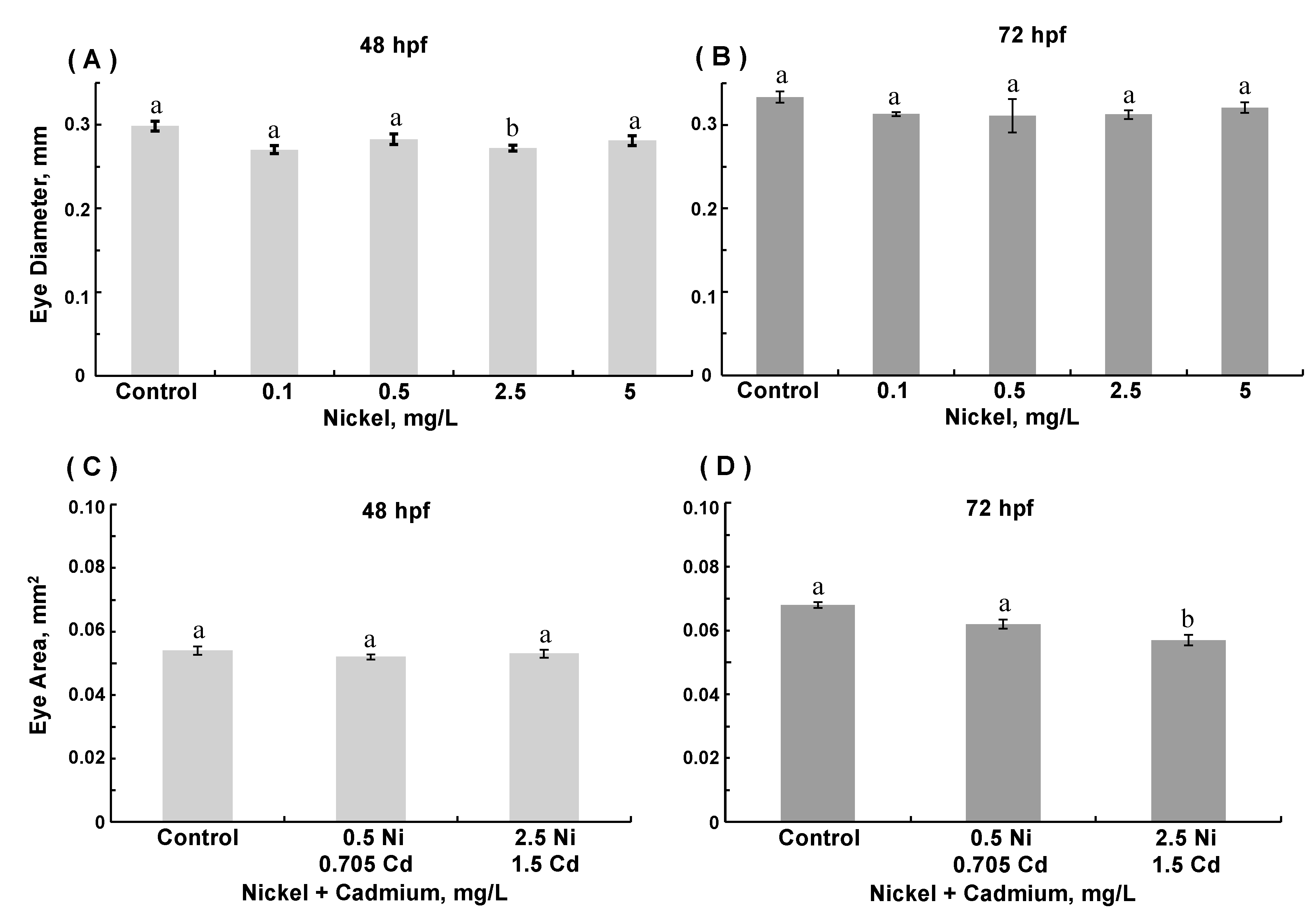
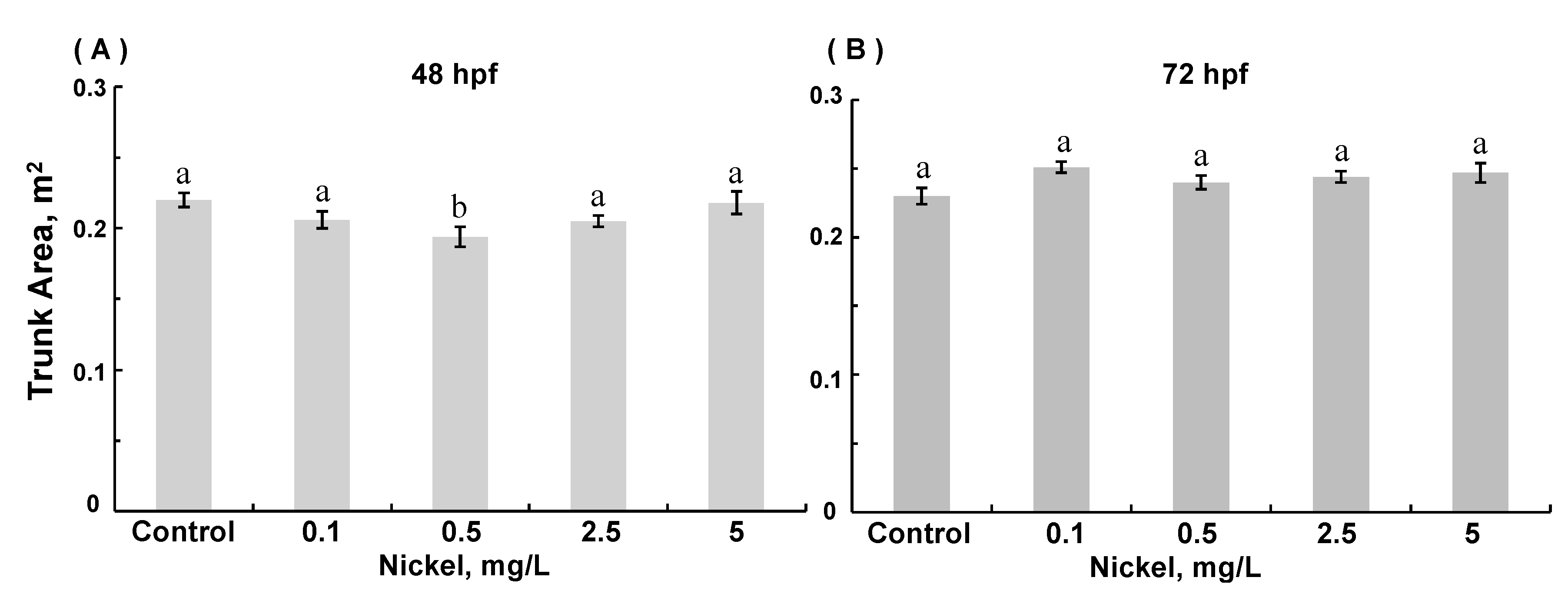
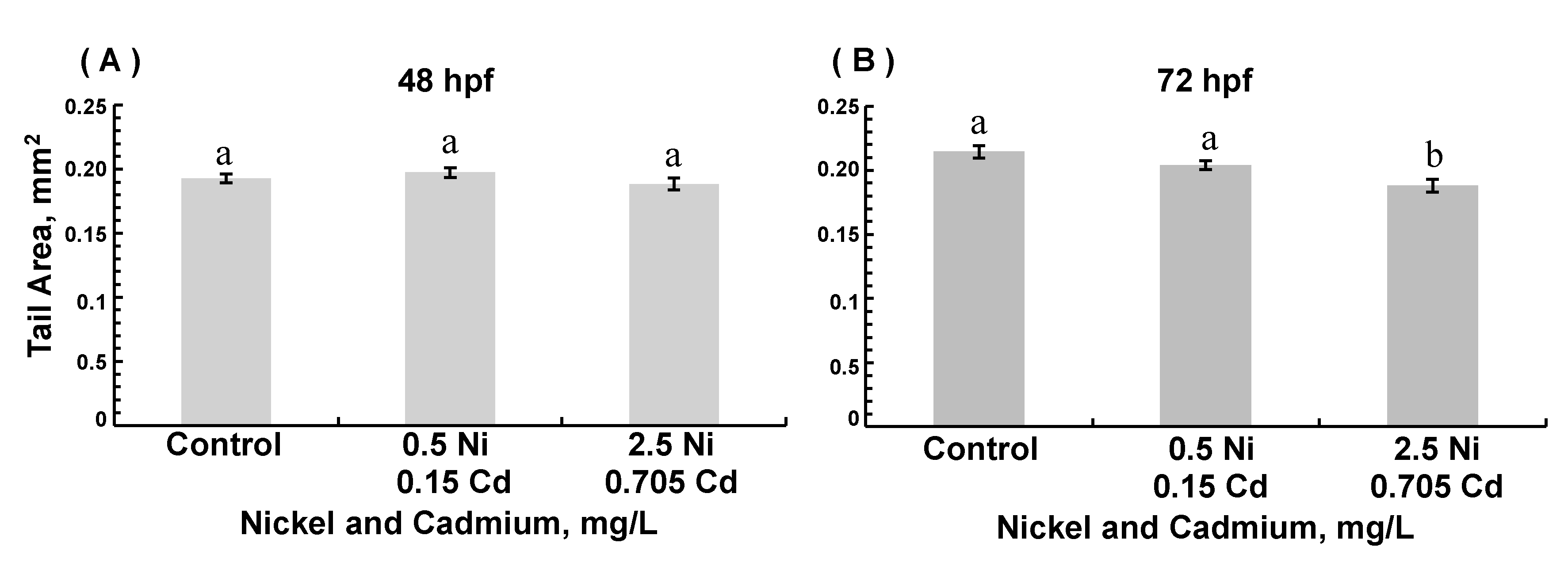
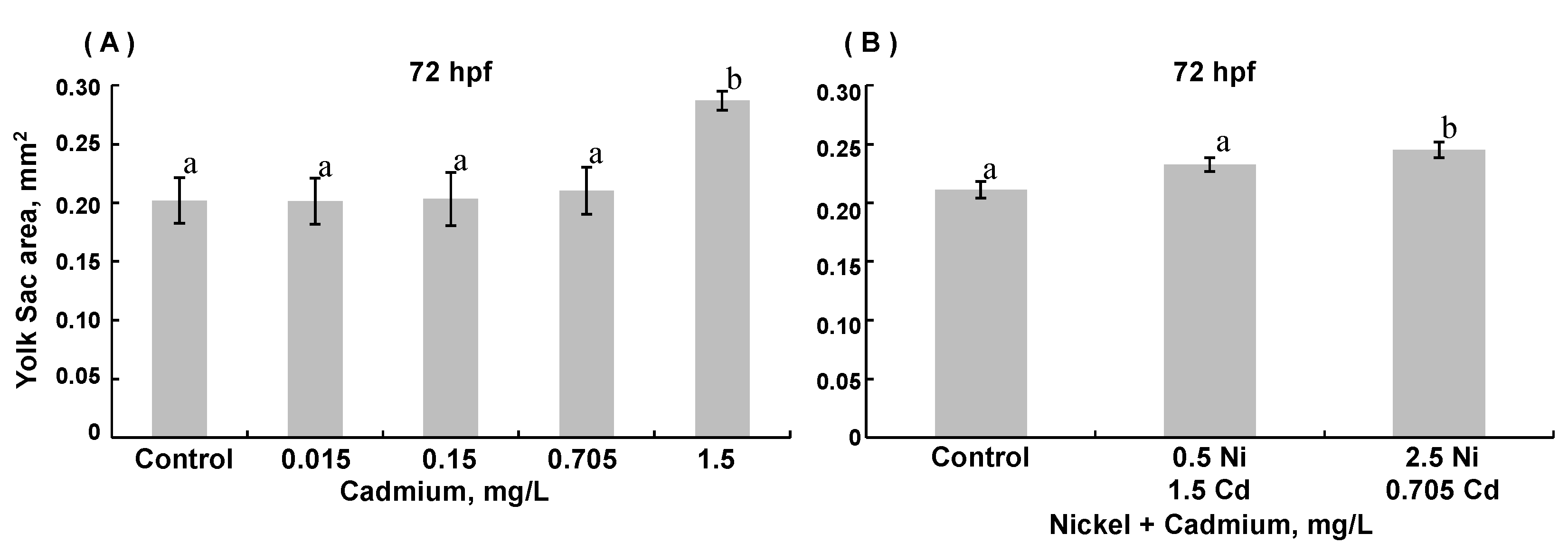
3.3. Morphological Assessments
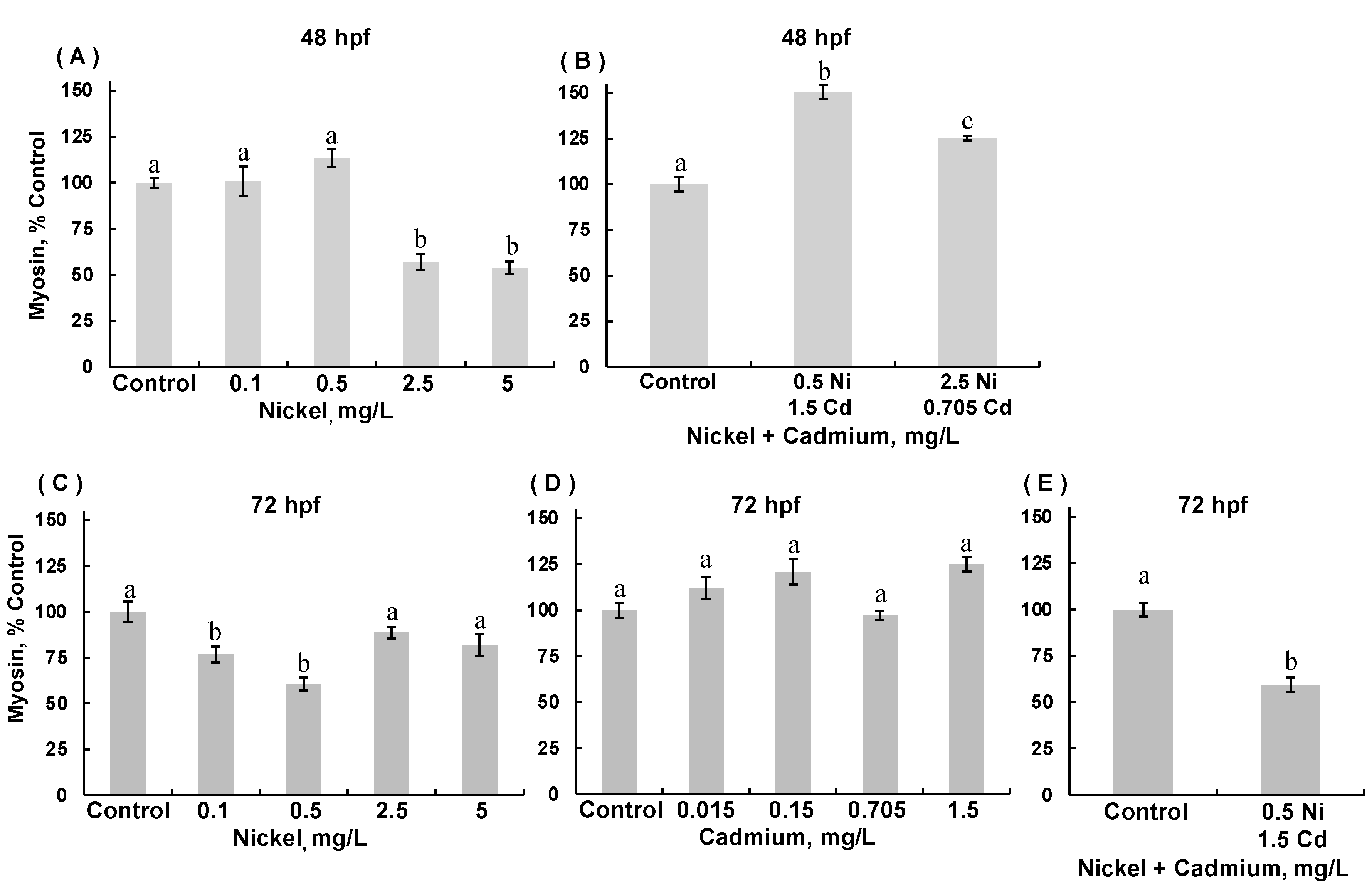
3.4. Myosin Expression in Skeletal Muscle
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Health Risks of Heavy Metals from Long-Range Transboundary Air Pollution. Copenhagen, World Health Organization Regional Office for Europe 2007. Available online: http://www.euro.who.int/document/E91044.pdf (accessed on 30 June 2019).
- Rathor, G.; Chopra, N.; Adhikari, T. Nickel as a pollutant and its management. Int. Res. J. Environ. Sci. 2014, 3, 94–98. [Google Scholar]
- WHO. Exposure to Cadmium: A Major Public Health Concern. Geneva, World Health Organization Regional Office for Europe 2010. Available online: http://www.who.int/ipcs/features/cadmium.pdf (accessed on 30 June 2019).
- Boluda, R.; Errecalde, F.; Lagarda, M.J. Environmental cadmium, lead and nickel contamination: Possible relationship between soil and vegetable content. Anal. Bioanal. Chem. 1991, 339, 654–657. [Google Scholar] [CrossRef]
- Gil, C.; Boluda, R.; Miras, J.J.R.; Gil, C. Determination and evaluation of cadmium, lead and nickel in greenhouse soils of Almería (Spain). Chemosphere 2004, 55, 1027–1034. [Google Scholar] [CrossRef]
- Olsson, I.-M.; Bensryd, I.; Lundh, T.; Ottosson, H.; Skerfving, S.; Oskarsson, A. Cadmium in blood and urine—Impact of sex, age, dietary intake, iron status, and former smoking—Association of renal effects. Environ. Health Perspect. 2002, 110, 1185–1190. [Google Scholar] [CrossRef]
- Yu, C.; Ling, Q.; Yan, S.; Li, J.; Chen, Z.; Peng, Z. Cadmium contamination in various environmental materials in an industrial area, Hangzhou, China. Chem. Speciat. Bioavailab. 2010, 22, 35–42. [Google Scholar] [CrossRef]
- Abudayyak, M.; Guzel, E.; Özhan, G. Nickel oxide nanoparticles are highly toxic to SH-SY5Y neuronal cells. Neurochem. Int. 2017, 108, 7–14. [Google Scholar] [CrossRef]
- Robinson, B.H. E-waste: An assessment of global production and environmental impacts. Sci. Total. Environ. 2009, 408, 183–191. [Google Scholar] [CrossRef]
- Redelstein, R.; Zielke, H.; Spira, D.; Feiler, U.; Erdinger, L.; Zimmer, H.; Wiseman, S.; Hecker, M.; Giesy, J.P.; Seiler, T.-B.; et al. Bioaccumulation and molecular effects of sediment-bound metals in zebrafish embryos. Environ. Sci. Pollut. Res. 2015, 22, 16290–16304. [Google Scholar] [CrossRef]
- Das, K.K.; Das, S.N.; Dhundasi, S.A. Nickel: Molecular Diversity, Application, Essentiality and Toxicity in Human Health. Biometals: Molecular Structures, Binding Properties and Applications; Nova Science Publishers: New York, NY, USA, 2010; pp. 33–58. [Google Scholar]
- Jacobo-Estrada, T.; Santoyo-Sánchez, M.; Thévenod, F.; Barbier, O. Cadmium Handling, Toxicity and Molecular Targets Involved during Pregnancy: Lessons from Experimental Models. Int. J. Mol. Sci. 2017, 18, 1590. [Google Scholar] [CrossRef] [Green Version]
- Jijie, R.; Solcan, G.; Nicoara, M.; Micu, D.; Strungaru, S. Antagonistic effects in zebrafish (Danio rerio) behavior and oxidative stress induced by toxic metals and deltamethrin acute exposure. Sci. Total. Environ. 2020, 698, 134299. [Google Scholar] [CrossRef] [PubMed]
- Sonnack, L.; Klawonn, T.; Kriehuber, R.; Hollert, H.; Schäfers, C.; Fenske, M. Comparative analysis of the transcriptome responses of zebrafish embryos after exposure to low concentrations of cadmium, cobalt and copper. Comp. Biochem. Physiol. Part D Genom. Proteom. 2018, 25, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Lahnsteiner, F. The sensitivity and reproducibility of the zebrafish (Danio rerio) embryo test for the screening of waste water quality and for testing the toxicity of chemicals. Altern. Lab. Anim. 2008, 36, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.-L.; Ru, Y.-F.; Liu, M.; Tang, J.-N.; Zheng, J.-F.; Wu, B.; Gu, Y.-H.; Shi, H.-J. Reproductive effects of cadmium on sperm function and early embryonic development in vitro. PLoS ONE 2017, 12, e0186727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.; Bannigan, J. Cadmium: Toxic effects on the reproductive system and the embryo. Reprod. Toxicol. 2008, 25, 304–315. [Google Scholar] [CrossRef] [PubMed]
- National Research Council; Safe Drinking Water Committee. Drinking Water and Health, Volume 9; National Academies Press: Washington, DC, USA, 1989.
- Nagel, R. DarT: The embryo test with the Zebrafish Danio rerio—A general model in ecotoxicology and toxicology. Altex 2002, 19, 38–48. [Google Scholar] [PubMed]
- Dubińska-Magiera, M.; Daczewska, M.; Lewicka, A.; Migocka-Patrzałek, M.; Niedbalska-Tarnowska, J.; Jagla, K. Zebrafish: A Model for the Study of Toxicants Affecting Muscle Development and Function. Int. J. Mol. Sci. 2016, 17, 1941. [Google Scholar] [CrossRef]
- Krzykwa, J.C.; Saeid, A.; Jeffries, M.K.S. Identifying sublethal endpoints for evaluating neurotoxic compounds utilizing the fish embryo toxicity test. Ecotoxicol. Environ. Saf. 2019, 170, 521–529. [Google Scholar] [CrossRef]
- Fraysse, B.; Mons, R.; Garric, J. Development of a zebrafish 4-day embryo-larval bioassay to assess toxicity of chemicals. Ecotoxicol. Environ. Saf. 2006, 63, 253–267. [Google Scholar] [CrossRef]
- Weil, M.; Meißner, T.; Busch, W.; Springer, A.; Kühnel, D.; Schulz, R.; Duis, K. The oxidized state of the nanocomposite Carbo-Iron® causes no adverse effects on growth, survival and differential gene expression in zebrafish. Sci. Total. Environ. 2015, 198–208. [Google Scholar] [CrossRef]
- Ribeiro, R.X.; Brito, R.D.S.; Pereira, A.C.; Monteiro, K.B.E.S.; Gonçalves, B.B.; Rocha, T.L. Ecotoxicological assessment of effluents from Brazilian wastewater treatment plants using zebrafish embryotoxicity test: A multi-biomarker approach. Sci. Total Environ. 2020, 735, 139036. [Google Scholar] [CrossRef]
- Zindler, F.; Beedgen, F.; Brandt, D.; Steiner, M.; Stengel, D.; Baumann, L.; Braunbeck, T. Analysis of tail coiling activity of zebrafish (Danio rerio) embryos allows for the differentiation of neurotoxicants with different modes of action. Ecotoxicol. Environ. Saf. 2019, 186, 109754. [Google Scholar] [CrossRef] [PubMed]
- Chow, E.S.H.; Hui, M.N.Y.; Lin, C.C.; Cheng, S.H. Cadmium inhibits neurogenesis in zebrafish embryonic brain development. Aquat. Toxicol. 2008, 87, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Jones, I.; Kille, P.; Sweeney, G. Cadmium delays growth hormone expression during rainbow trout development. J. Fish Biol. 2005, 59, 1015–1022. [Google Scholar] [CrossRef]
- Witeska, M.; Jezierska, B.; Chaber, J. The influence of cadmium on common carp embryos and larvae. Aquaculture 1995, 129, 129–132. [Google Scholar] [CrossRef]
- Wu, S.M.; Tsai, P.J.; Chou, M.Y.; Wang, W.-D. Effects of Maternal Cadmium Exposure on Female Reproductive Functions, Gamete Quality, and Offspring Development in Zebrafish (Danio rerio). Arch. Environ. Contam. Toxicol. 2013, 65, 521–536. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Liu, Z.; Liu, F.; Ye, Y.; Peng, T.; Ota, T. Embryonic exposure to cadmium (II) and chromium (VI) induce behavioral alterations, oxidative stress and immunotoxicity in zebrafish (Danio rerio). Neurotoxicology Teratol. 2015, 48, 9–17. [Google Scholar] [CrossRef]
- Nguyen, C.R.J.L.T.H.; Nguyen, L.T.H.; Janssen, C.R. Embryo-Larval Toxicity Tests with the African Catfish (Clarias gariepinus): Comparative Sensitivity of Endpoints. Arch. Environ. Contam. Toxicol. 2002, 42, 256–262. [Google Scholar] [CrossRef]
- Lefauve, M.K.; Connaughton, V.P. Developmental exposure to heavy metals alters visually-guided behaviors in zebrafish. Curr. Zool. 2017, 63, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Scheil, V.; Zürn, A.; Triebskorn, R.; Köhler, H.-R. Embryo development, stress protein (Hsp70) responses, and histopathology in zebrafish (Danio rerio) following exposure to nickel chloride, chlorpyrifos, and binary mixtures of them. Environ. Toxicol. 2009, 25, 83–93. [Google Scholar] [CrossRef]
- Kim, K.; Wang, C.-H.; Ok, Y.S.; Lee, S.-E. Heart developmental toxicity by carbon black waste generated from oil refinery on zebrafish embryos (Danio rerio): Combined toxicity on heart function by nickel and vanadium. J. Hazard. Mater. 2019, 363, 127–137. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio Rerio), 4th ed.; University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Khalid, S.; Shahid, M.; Shah, A.H.; Saeed, F.; Ali, M.; Qaisrani, S.A.; Dumat, C. Heavy metal contamination and exposure risk assessment via drinking groundwater in Vehari, Pakistan. Environ. Sci. Pollut. Res. 2020, 1–13. [Google Scholar] [CrossRef]
- Saravi, S.S.; Karami, B.; Karami, S.; Shokrzadeh, M. Evaluation of Metal Pollution in Fish and Water Collected from Gorgan Coast of the Caspian Sea, Iran. Bull. Environ. Contam. Toxicol. 2012, 89, 419–423. [Google Scholar] [CrossRef]
- Corales-Ultra, O.G.; Peja, R.P.; Casas, E.V. Baseline study on the levels of heavy metals in seawater and macroalgae near an abandoned mine in Manicani, Guiuan, Eastern Samar, Philippines. Mar. Pollut. Bull. 2019, 149, 110549. [Google Scholar] [CrossRef] [PubMed]
- Kline, T.L.; Sussman, C.R.; Irazabal, M.V.; Mishra, P.K.; Pearson, E.A.; Torres, V.E.; Macura, S.I. Three-dimensional NMR microscopy of zebrafish specimens. NMR Biomed. 2018, 32, e4031. [Google Scholar] [CrossRef] [PubMed]
- Abbott, L.; Jacobowitz, D. Development of calretinin-immunoreactive unipolar brush-like cells and an afferent pathway to the embryonic and early postnatal mouse cerebellum. Brain Struct. Funct. 1995, 191, 541–599. [Google Scholar] [CrossRef]
- Abramoff, M.D.; Magalhaes, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Saint-Amant, L.; Drapeau, P. Time course of the development of motor behaviors in the zebrafish embryo. J. Neurobiol. 1998, 37, 622–632. [Google Scholar] [CrossRef]
- Beattie, J.H.; Pascoe, D. Cadmium uptake by rainbow trout, Salmo gairdneri eggs and alevins. J. Fish Biol. 1978, 13, 631–637. [Google Scholar] [CrossRef]
- Micihibata, H. Uptake and distribution of cadmium in the egg of the teleost, Oryzias latipes. J. Fish Biol. 1981, 19, 691–696. [Google Scholar] [CrossRef]
- Jezierska, B.; Ługowska, K.; Witeska, M. The effects of heavy metals on embryonic development of fish (a review). Fish Physiol. Biochem. 2008, 35, 625–640. [Google Scholar] [CrossRef]
- Peterson, R.H.; Martin-Robichaud, D.J. Perivitelline and Vitelline Potentials in Teleost Eggs as Influenced by Ambient Ionic Strength, Natal Salinity, and Electrode Electrolyte; and the Influence of these Potentials on Cadmium Dynamics within the Egg. Can. J. Fish. Aquat. Sci. 1986, 43, 1445–1450. [Google Scholar] [CrossRef]
- Liu, L.; Yan, Y.; Wang, J.; Wu, W.; Xu, L. Generation ofmt:egfptransgenic zebrafish biosensor for the detection of aquatic zinc and cadmium. Environ. Toxicol. Chem. 2016, 35, 2066–2073. [Google Scholar] [CrossRef] [PubMed]
- Consigli, V.; Guarienti, M.; Bilo, F.; Benassi, L.; Depero, L.E.; Bontempi, E.; Presta, M. Evaluation of the Biotoxicity of Tree Wood Ashes in Zebrafish Embryos. Zebrafish 2016, 13, 449–455. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Kim, S.-J.; Bae, M.A.; Cho, K.-H.; Kim, J.-R. Cadmium exposure exacerbates severe hyperlipidemia and fatty liver changes in zebrafish via impairment of high-density lipoproteins functionality. Toxicol. Vitr. 2018, 47, 249–258. [Google Scholar] [CrossRef]
- Xiu, R. Toxicity of mercury, copper, nickel, lead, and cobalt to embryos and larvae of zebrafish, Brachydanio rerio. Arch. Environ. Contam. Toxicol. 1991, 21, 126–134. [Google Scholar] [CrossRef]
- Peng, G.; He, Y.; Zhao, M.; Yu, T.; Qin, Y.; Lin, S. Differential effects of metal oxide nanoparticles on zebrafish embryos and developing larvae. Environ. Sci. Nano 2018, 5, 1200–1207. [Google Scholar] [CrossRef]
- Kienle, C.; Kohler, H.-R.; Filser, J.; Gerhardt, A. Effects of nickel chloride and oxygen depletion on behaviour and vitality of zebrafish (Danio rerio, Hamilton, 1822) (Pisces, Cypriniformes) embryos and larvae. Environ. Pollut. 2008, 152, 612–620. [Google Scholar] [CrossRef]
- Hagenmaier, H.E. The hatching process in fish embryos. Dev. Genes Evol. 1974, 175, 157–162. [Google Scholar] [CrossRef]
- Nebeker, A.V.; Savonen, C.; Stevens, D.G. Sensitivity of rainbow trout early stages to nickel chloride. Environ. Toxicol. Chem. Int. J. 1985, 2, 233–239. [Google Scholar] [CrossRef]
- Novelli, E.; Hernandes, R.; Filho, J.N.; Barbosa, L. Differential/combined effect of water contamination with cadmium and nickel on tissues of rats. Environ. Pollut. 1998, 103, 295–300. [Google Scholar] [CrossRef]
- Dang, Y.; Wang, F.; Liu, C. Real-time PCR array to study the effects of chemicals on the growth hormone/insulin-like growth factors (GH/IGFs) axis of zebrafish embryos/larvae. Chemosphere 2018, 207, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cao, H.; Meng, Y.; Jin, G.; Zhu, M. The toxicity of cadmium (Cd2+) towards embryos and pro-larva of soldatov’s catfish (Silurus soldatovi). Ecotoxicol. Environ. Saf. 2012, 80, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Liu, L.; Li, D.-L.; Ling, F.; Wang, G. Developmental toxicity in rare minnow (Gobiocypris rarus) embryos exposed to Cu, Zn and Cd. Ecotoxicol. Environ. Saf. 2014, 104, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Si, J.; Wei, Y.; Li, S.; Jiang, Y.; Zhou, R.; Liu, B.; Zhang, H. Toxicity of porcelain-fused-to-metal substrate to zebrafish (Danio rerio) embryos and larvae. Life Sci. 2018, 203, 66–71. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Patterson, J.; Kimmel, R.O. The development and behavioral characteristics of the startle response in the zebra fish. Dev. Psychobiol. 1974, 7, 47–60. [Google Scholar] [CrossRef]
- Katayama, R.; Yamane, A.; Fukui, T. Changes in the Expression of Myosins During Postnatal Development of Masseter Muscle in the Microphthalmic Mouse. Open Dent. J. 2010, 4, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talmadge, R.J.; Acosta, W.; Garland, T. Myosin heavy chain isoform expression in adult and juvenile mini-muscle mice bred for high-voluntary wheel running. Mech. Dev. 2014, 134, 16–30. [Google Scholar] [CrossRef]
- He, M.-D.; Xu, S.-C.; Lu, Y.-H.; Li, L.; Zhong, M.; Zhang, Y.-W.; Wang, Y.; Li, M.; Yang, J.; Zhang, G.-B.; et al. L-carnitine protects against nickel-induced neurotoxicity by maintaining mitochondrial function in Neuro-2a cells. Toxicol. Appl. Pharmacol. 2011, 253, 38–44. [Google Scholar] [CrossRef]
- Xu, S.; He, M.; Zhong, M.; Li, L.; Lu, Y.; Zhang, Y.; Zhang, L.; Yu, Z.; Zhou, Z. The neuroprotective effects of taurine against nickel by reducing oxidative stress and maintaining mitochondrial function in cortical neurons. Neurosci. Lett. 2015, 590, 52–57. [Google Scholar] [CrossRef]
- Morrice, J.R.; Gregory-Evans, C.Y.; Shaw, C. Modeling Environmentally-Induced Motor Neuron Degeneration in Zebrafish. Sci. Rep. 2018, 8, 4890. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n) | 0.1 mg/L Nickel (n) | 0.5 mg/L Nickel (n) | 2.5 mg/L Nickel (n) | 5 mg/L Nickel (n) | |
|---|---|---|---|---|---|
| No Movement | 50 | 71 * | 67 * | 68 * | 67 * |
| Movement | 55 | 34 * | 38 * | 37 * | 38 * |
| Distance | Control (n) | Exposed (n) |
|---|---|---|
| 0–25 mm | 12 | 26 * |
| 26–50+ mm | 37 | 34 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aldavood, S.J.; Abbott, L.C.; Evans, Z.R.; Griffin, D.J.; Lee, M.D.; Quintero-Arevalo, N.M.; Villalobos, A.R. Effect of Cadmium and Nickel Exposure on Early Development in Zebrafish (Danio rerio) Embryos. Water 2020, 12, 3005. https://doi.org/10.3390/w12113005
Aldavood SJ, Abbott LC, Evans ZR, Griffin DJ, Lee MD, Quintero-Arevalo NM, Villalobos AR. Effect of Cadmium and Nickel Exposure on Early Development in Zebrafish (Danio rerio) Embryos. Water. 2020; 12(11):3005. https://doi.org/10.3390/w12113005
Chicago/Turabian StyleAldavood, Seyed Javid, Louise C. Abbott, Zachary R. Evans, Daniel J. Griffin, MaKenzie D. Lee, Natalia M. Quintero-Arevalo, and Alice R. Villalobos. 2020. "Effect of Cadmium and Nickel Exposure on Early Development in Zebrafish (Danio rerio) Embryos" Water 12, no. 11: 3005. https://doi.org/10.3390/w12113005
APA StyleAldavood, S. J., Abbott, L. C., Evans, Z. R., Griffin, D. J., Lee, M. D., Quintero-Arevalo, N. M., & Villalobos, A. R. (2020). Effect of Cadmium and Nickel Exposure on Early Development in Zebrafish (Danio rerio) Embryos. Water, 12(11), 3005. https://doi.org/10.3390/w12113005