Reforestation Based on Mono-Plantation of Fast-Growing Tree Species Make It Difficult to Maintain (High) Soil Water Content in Tropics, a Case Study in Hainan Island, China


{kind=link}
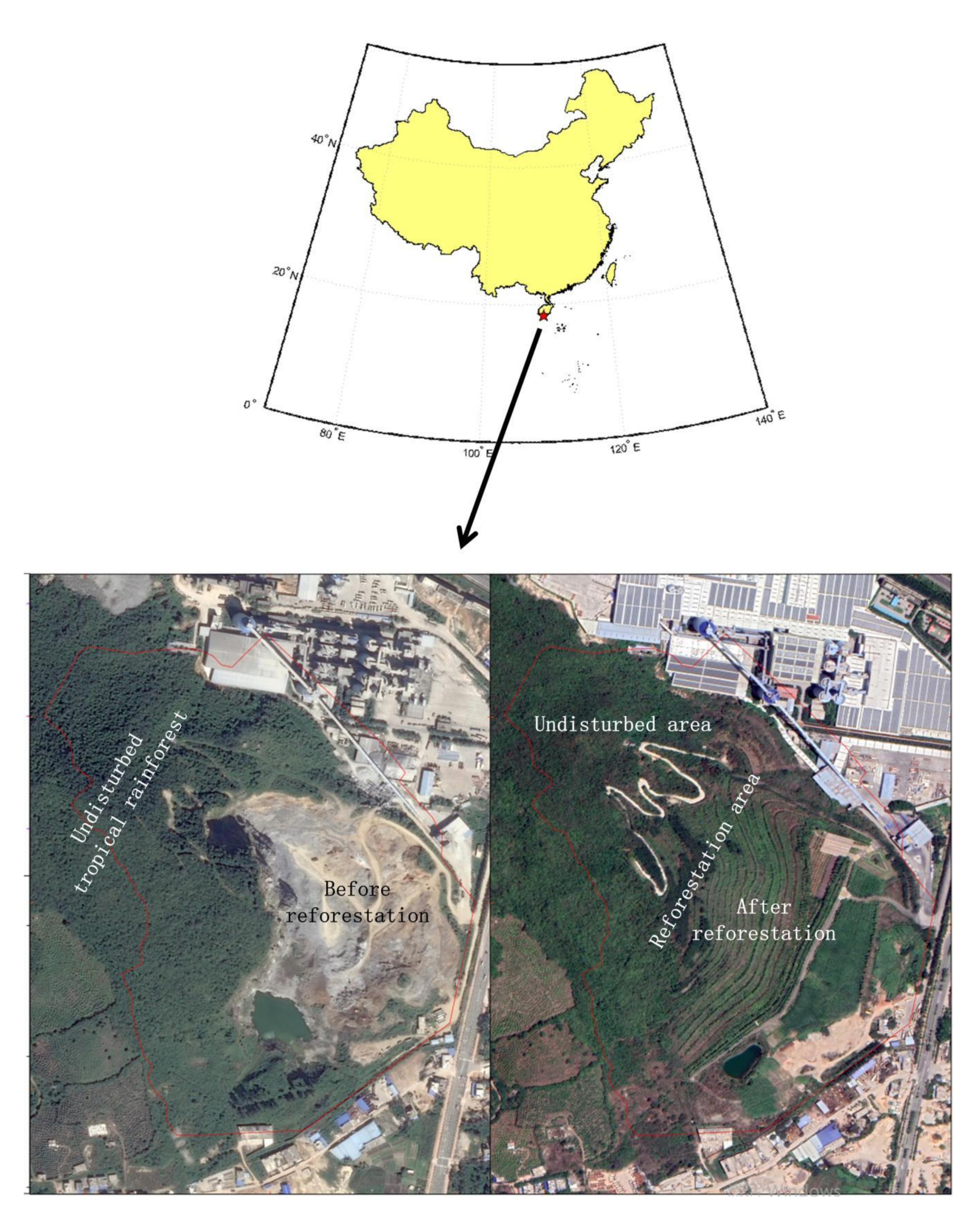{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Sampling
2.3. Statistics Methods
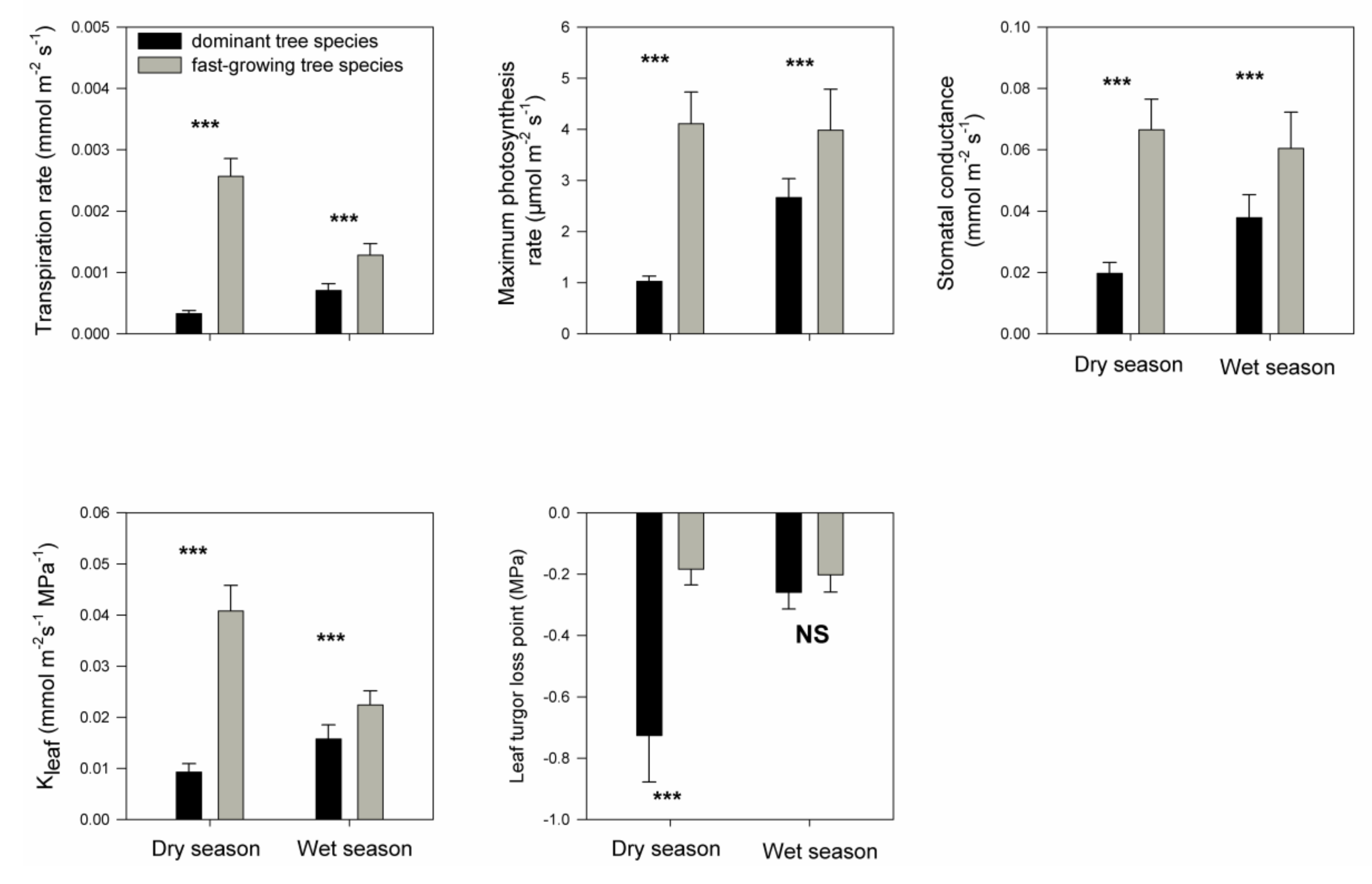
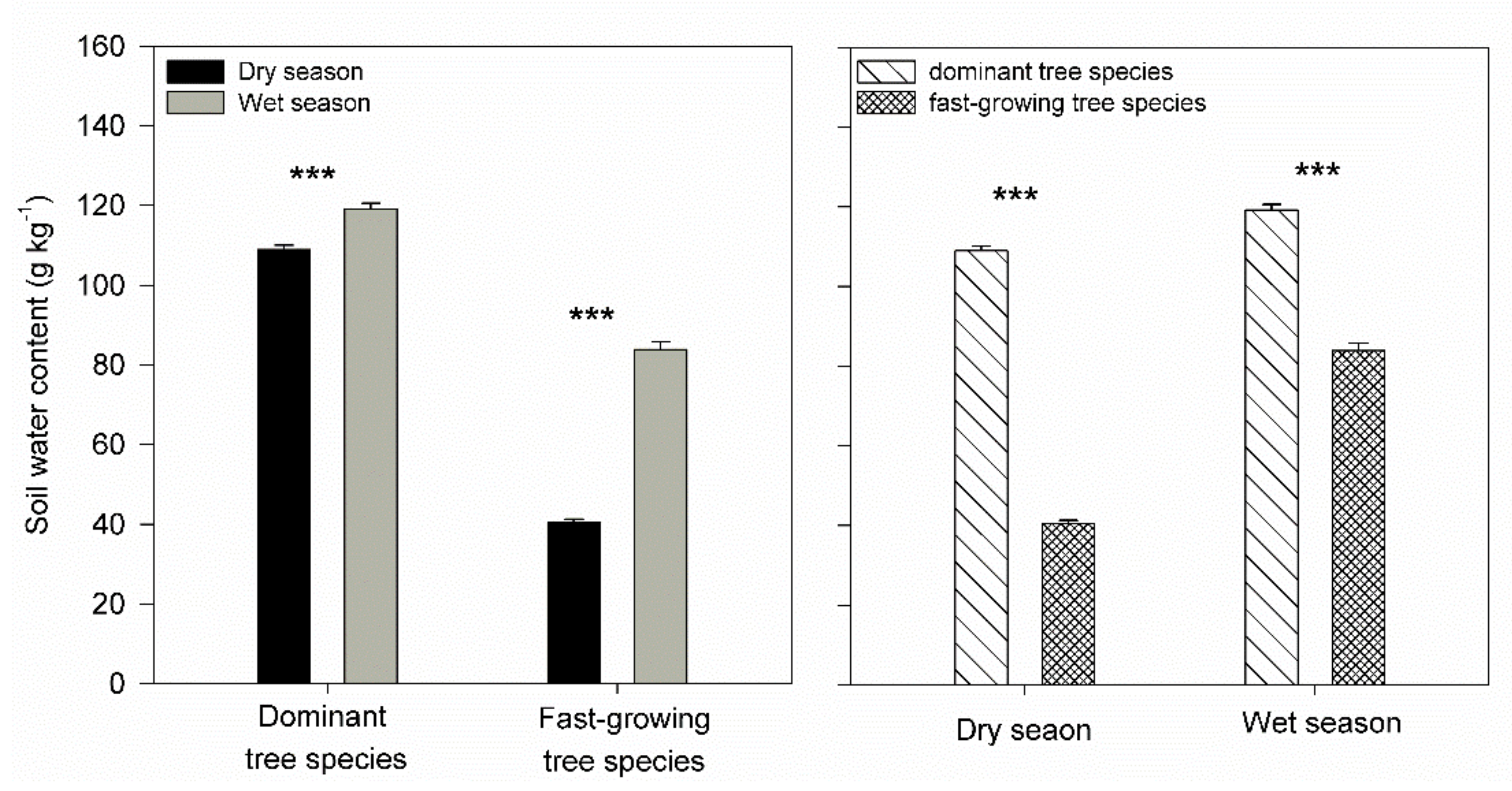
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mekonnen, M.M.; Hoekstra, A.Y. Four billion people facing severe water scarcity. Sci. Adv. 2016, 2, e1500323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Famiglietti, J.S.; Rodell, M. Water in the Balance. Science 2013, 340, 1300. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Wei, X.; Chen, X.; Zhou, P.; Liu, X.; Xiao, Y.; Sun, G.; Scott, D.F.; Zhou, S.; Han, L.; et al. Global pattern for the effect of climate and land cover on water yield. Nat. Commun. 2015, 6, 5918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, V.; Hall, M.J. The effects of afforestation and deforestation on water yields. J. Hydrol. 1996, 178, 293–309. [Google Scholar] [CrossRef]
- Lambin, E.F.; Meyfroidt, P. Global land use change, economic globalization, and the looming land scarcity. Proc. Natl. Acad. Sci. USA 2011, 108, 3465–3472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vörösmarty, C.J.; Hoekstra, A.Y.; Bunn, S.E.; Conway, D.; Gupta, J. Fresh water goes global. Science 2015, 349, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castello, L.; Macedo, M.N. Large-scale degradation of Amazonian freshwater ecosystems. Glob. Chang. Biol. 2016, 22, 990–1007. [Google Scholar] [CrossRef]
- Solange, F.; Ometto, B.M.; Weiss, K.C.B.; Palmer, M.A.; Silva, L.C.R. Impacts of forest restoration on water yield: A systematic review. PLoS ONE 2017, 12, e0183210. [Google Scholar]
- Peña-Arancibia, J.L.; Bruijnzeel, L.A.; Mulligan, M.; van Dijk, A.I.J.M. Forests as ‘sponges’ and ‘pumps’: Assessing the impact of deforestation on dry-season flows across the tropics. J. Hydrol. 2019, 574, 946–963. [Google Scholar] [CrossRef]
- Bruijnzeel, L.A. Hydrological functions of tropical forests: Not seeing the soil for the trees? Agric. Ecosyst. Environ. 2004, 104, 185–228. [Google Scholar] [CrossRef]
- Tardieu, F.; Parent, B. Predictable ‘meta-mechanisms’ emerge from feedbacks between transpiration and plant growth and cannot be simply deduced from short-term mechanisms. Plant Cell Environ. 2017, 40, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, J.; Bonell, M.; Venkatesh, B.; Purandara, B.K.; Rakesh, K.N.; Lele, S.; Kiran, M.C.; Reddy, V.; Badiger, S. The groundwater recharge response and hydrologic services of tropical humid forest ecosystems to use and reforestation: Support for the “infiltration-evapotranspiration trade-off hypothesis”. J. Hydrol. 2013, 498, 191–209. [Google Scholar] [CrossRef]
- Tuzet, A.; Perrier, A.; Leuning, R. A coupled model of stomatal conductance, photosynthesis and transpiration. Plant Cell Environ. 2003, 26, 1097–1116. [Google Scholar] [CrossRef]
- Gebrehiwot, S.G.; Ellison, D.; Bewket, W.; Seleshi, Y.; Inogwabini, B.-I.; Bishop, K. The Nile Basin waters and the West African rainforest: Rethinking the boundaries. Wiley Interdiscip. Rev. Water 2019, 6, e1317. [Google Scholar] [CrossRef] [Green Version]
- Mapulanga, A.M.; Naito, H. Effect of deforestation on access to clean drinking water. Proc. Natl. Acad. Sci. USA 2019, 116, 8249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guidicini, G.; Iwasa, O.Y. Tentative correlation between rainfall and landslides in a humid tropical environment. Bull. Eng. Geol. Environ. 1977, 16, 13–20. [Google Scholar] [CrossRef]
- Chang, K.T.; Chiang, S.H.; Lei, F. Analysing the Relationship between Typhoon-Triggered Landslides and Critical Rainfall Conditions. Earth Surf. Proc. Land. 2008, 33, 1261–1271. [Google Scholar] [CrossRef]
- Yumul, J.G.P.; Servando, N.T.; Suerte, L.O.; Magarzo, M.Y.; Juguan, L.V.V.; Dimalanta, C.B. Tropical cyclone–southwest monsoon interaction and the 2008 floods and landslides in Panay island, central Philippines: Meteorological and geological factors. Nat. Hazards 2012, 62, 827–840. [Google Scholar] [CrossRef]
- Acosta, L.A.; Eugenio, E.A.; Macandog, P.B.M.; Macandog, D.B.M.; Lin, E.K.H.; Abucay, E.R.; Cura, A.L.; Primavera, M.G. Loss and damage from typhoon-induced floods and landslides in the Philippines: Community Perceptions on climate impacts and adaptation options. Int. J. Glob. Warm. 2016, 9, 33–65. [Google Scholar] [CrossRef]
- Gao, Q.; Yu, M. Reforestation-induced changes of landscape composition and configuration modulate freshwater supply and flooding risk of tropical watersheds. PLoS ONE 2017, 12, e0181315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villamayor, B.M.R.; Rollon, R.N.; Samson, M.S.; Albano, G.M.G.; Primavera, J.H. Impact of Haiyan on Philippine mangroves: Implications to the fate of the widespread monospecific Rhizophora plantations against strong typhoons. Ocean Coast. Manag. 2016, 132, 1–14. [Google Scholar] [CrossRef]
- Stokes, A.; Atger, C.; Bengough, A.G.; Fourcaud, T.; Sidle, R.C. Desirable plant root traits for protecting natural and engineered slopes against landslides. Plant Soil 2009, 324, 1–30. [Google Scholar] [CrossRef]
- Walker, L.R.; Velázquez, E.; Shiels, A.B. Applying lessons from ecological succession to the restoration of landslides. Plant. Soil. 2009, 324, 157–168. [Google Scholar] [CrossRef]
- Pang, C.C.; Ma, X.K.-K.; Lo, J.P.-L.; Hung, T.T.-H.; Hau, B.C.-H. Vegetation succession on landslides in Hong Kong: Plant regeneration, survivorship and constraints to restoration. Glob. Ecol. Conserv. 2018, 15, e00428. [Google Scholar] [CrossRef]
- Miyashita, K.; Tanakamaru, S.; Maitani, T.; Kimura, K. Recovery responses of photosynthesis, transpiration, and stomatal conductance in kidney bean following drought stress. Environ. Exp. Bot. 2005, 53, 205–214. [Google Scholar] [CrossRef]
- Fisher, R.A.; Williams, M.; Da Costa, A.L.; Malhi, Y.; Da Costa, R.F.; Almeida, S.; Metr, P. The response of an Eastern Amazonian rain forest to drought stress: Results and modelling analyses from a throughfall exclusion experiment. Global Change Biol. 2007, 13, 2361–2378. [Google Scholar] [CrossRef]
- Maherali, H.; Sherrard, M.E.; Clifford, M.H.; Latta, R.G. Leaf hydraulic conductivity and photosynthesis are genetically correlated in an annual grass. New Phytol. 2008, 180, 240–247. [Google Scholar] [CrossRef]
- Santos, V.A.H.F.d.; Ferreira, M.J.; Rodrigues, J.V.F.C.; Garcia, M.N.; Ceron, J.V.B.; Nelson, B.W.; Saleska, S.R. Causes of reduced leaf-level photosynthesis during strong El Niño drought in a Central Amazon forest. Global Change Biol. 2018, 24, 4266–4279. [Google Scholar] [CrossRef]
- White, D.A.; McGrath, J.F.; Ryan, M.G.; Battaglia, M.; Mendham, D.S.; Kinal, J.; Downes, G.M.; Crombie, D.S.; Hunt, M.E. Managing for water-use efficient wood production in Eucalyptus globulus plantations. Forest Ecol. Manag. 2014, 331, 272–280. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, H.Y.H.; Lian, J.; John, R.; Li, R.; Liu, H.; Ye, W.; Berninger, F.; Ye, Q. Using functional trait diversity patterns to disentangle the scale-dependent ecological processes in a subtropical forest. Func. Ecol. 2018, 32, 1379–1389. [Google Scholar] [CrossRef]
- Zhang, H.; John, R.; Zhu, S.; Liu, H.; Xu, Q.; Qi, W.; Liu, K.; Chen, H.Y.H.; Ye, Q. Shifts in functional trait–species abundance relationships over secondary subalpine meadow succession in the Qinghai-Tibetan Plateau. Oecologia 2018, 188, 547–557. [Google Scholar] [CrossRef]
- Luo, J.H.; Cui, J.; Shree, P.P.; Jiang, K.; Tan, Z.Y.; He, Q.F.; Zhang, H.; Long, W.X. Seasonally distinctive growth and drought stress functional traits enable Leucaena Leucocephala to successfully invade a Chinese tropical forest. Trop. Conserv. Sci. 2020, 9, 1–7. [Google Scholar]
- Bartlett, M.K.; Scoffoni, C.; Sack, L. The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: A global meta-analysis. Ecology 2016, 97, 503–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, K.; Pan, M.; Li, H.; Wolf, A.; Wu, J.; Medvigy, D.; Caylor, K.K.; Sheffield, J.; Wood, E.F.; Liang, M.; et al. Photosynthetic seasonality of global tropical forests constrained by hydroclimate. Nature Geosci. 2015, 8, 284–289. [Google Scholar] [CrossRef]
- Wu, J.; Serbin, S.P.; Ely, K.; Wolfe, B. The response of stomatal conductance to seasonal drought in tropical forests. Global Change Biol. 2020, 26, 823–839. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Zhang, Y.; Yang, J.; Kreidler, N.; Sun, S.-W.; Lin, L.; Hu, Y.-H.; Cao, K.-F.; Sack, L. Drought tolerance as a driver of tropical forest assembly: Resolving spatial signatures for multiple processes. Ecology 2016, 97, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Kirschbaum, M.U. Does enhanced photosynthesis enhance growth? Lessons learned from CO2 enrichment studies. Plant Physiol. 2011, 155, 117–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Z.H.; Zhang, Y.P.; Song, Q.H.; Liu, W.J.; Deng, X.B.; Tang, J.W.; Yun, D.; Zhou, W.J.; Yang, L.Y.; Yu, G.R.; et al. Rubber plantations act as water pumps in tropical China. Geophys. Res. Lett. 2011, 38, 24406. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Shao, M.A.; Zhu, Y.; Liu, Z. Impacts of land use and plant characteristics on dried soil layers in different climatic regions on the Loess Plateau of China. Agric. Forest Meteorol. 2011, 151, 437–448. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, H.; Jian, S.; Liu, N. Tree plantations influence the abundance of ammonia-oxidizing bacteria in the soils of a coral island. Appl. Soil Ecol. 2019, 138, 220–222. [Google Scholar] [CrossRef]
- Mason, N.W.; Richardson, S.J.; Peltzer, D.A.; de Bello, F.; Wardle, D.A.; Allen, R.B. Changes in coexistence mechanisms along a long-term soil chronosequence revealed by functional trait diversity. J. Ecol. 2012, 100, 678–689. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, W.; Yang, J.; Luo, J.; Jiang, K.; Xu, J.; Zhang, H. Reforestation Based on Mono-Plantation of Fast-Growing Tree Species Make It Difficult to Maintain (High) Soil Water Content in Tropics, a Case Study in Hainan Island, China. Water 2020, 12, 3077. https://doi.org/10.3390/w12113077
Hong W, Yang J, Luo J, Jiang K, Xu J, Zhang H. Reforestation Based on Mono-Plantation of Fast-Growing Tree Species Make It Difficult to Maintain (High) Soil Water Content in Tropics, a Case Study in Hainan Island, China. Water. 2020; 12(11):3077. https://doi.org/10.3390/w12113077
Chicago/Turabian StyleHong, Wenjun, Jindian Yang, Jinhuan Luo, Kai Jiang, Junze Xu, and Hui Zhang. 2020. "Reforestation Based on Mono-Plantation of Fast-Growing Tree Species Make It Difficult to Maintain (High) Soil Water Content in Tropics, a Case Study in Hainan Island, China" Water 12, no. 11: 3077. https://doi.org/10.3390/w12113077
APA StyleHong, W., Yang, J., Luo, J., Jiang, K., Xu, J., & Zhang, H. (2020). Reforestation Based on Mono-Plantation of Fast-Growing Tree Species Make It Difficult to Maintain (High) Soil Water Content in Tropics, a Case Study in Hainan Island, China. Water, 12(11), 3077. https://doi.org/10.3390/w12113077