Ecosystem Metabolism in Small Ponds: The Effects of Floating-Leaved Macrophytes



Abstract
:1. Introduction
2. Materials and Methods
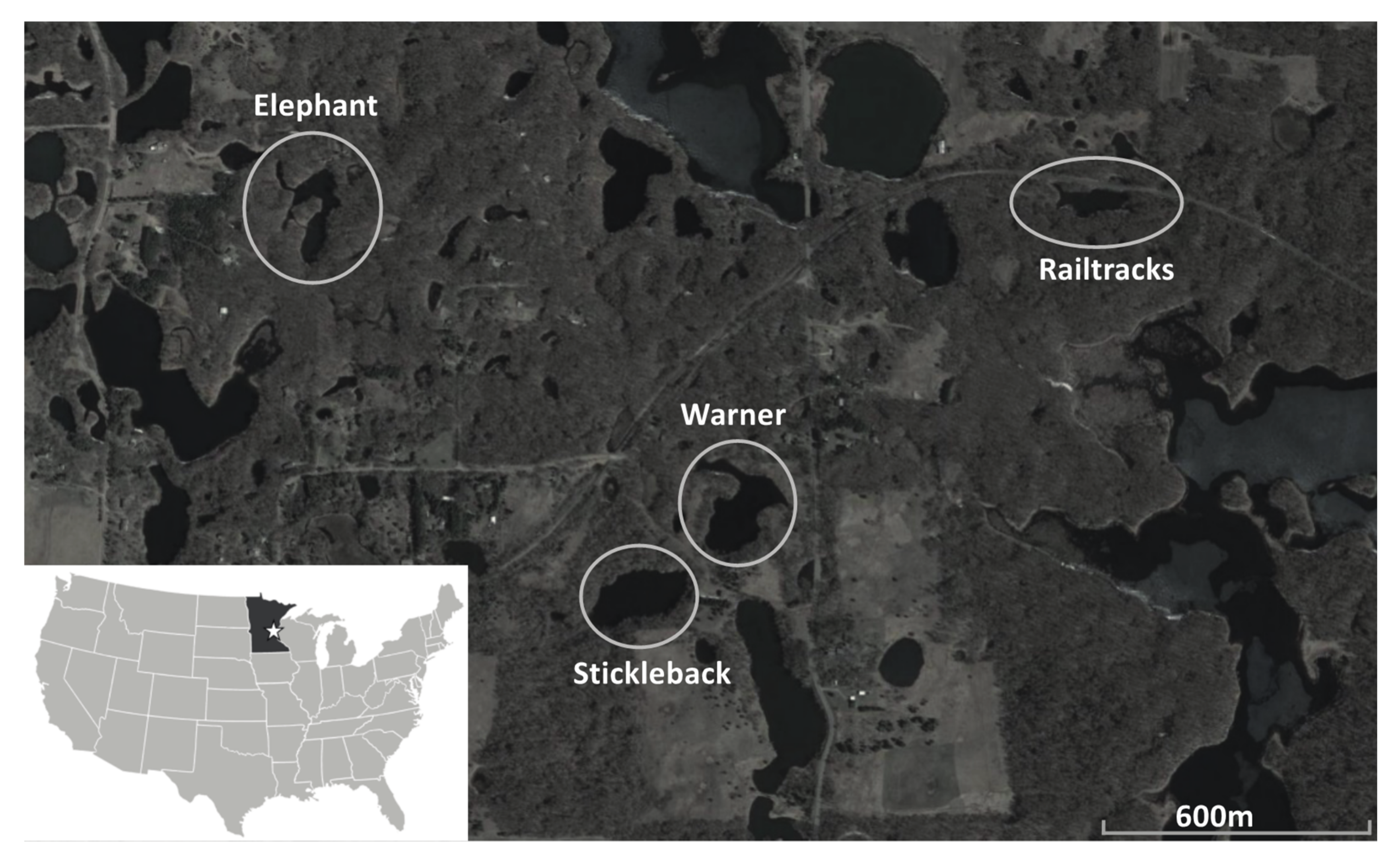
2.1. Pond Location
2.2. Physical and Chemical Parameters
2.3. Pond Primary Production
2.4. Pond Dissolved Oxygen Levels and Ecosystem Metabolism
2.5. Statistics
3. Results
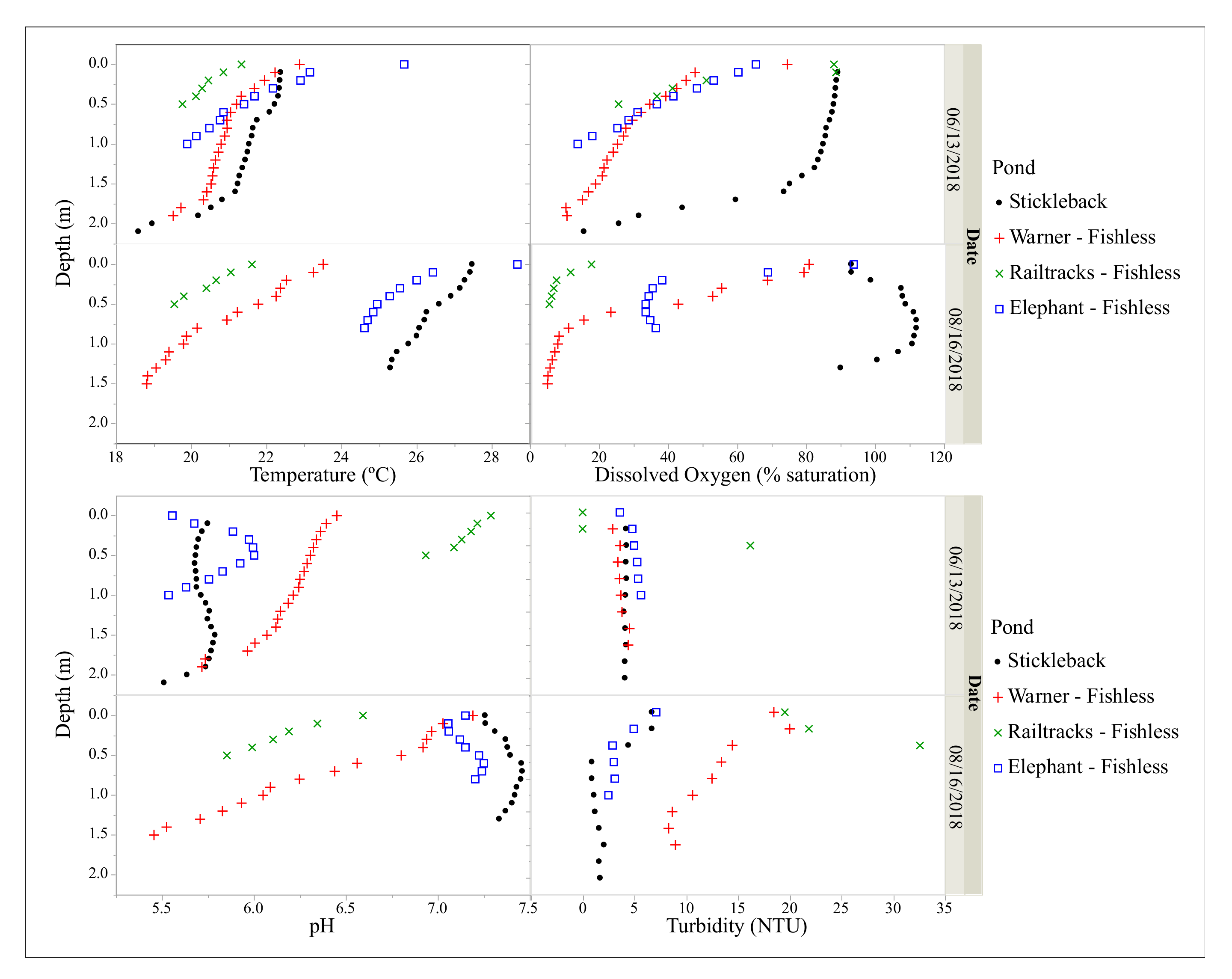
3.1. Physical and Chemical Parameters
3.2. Primary Production
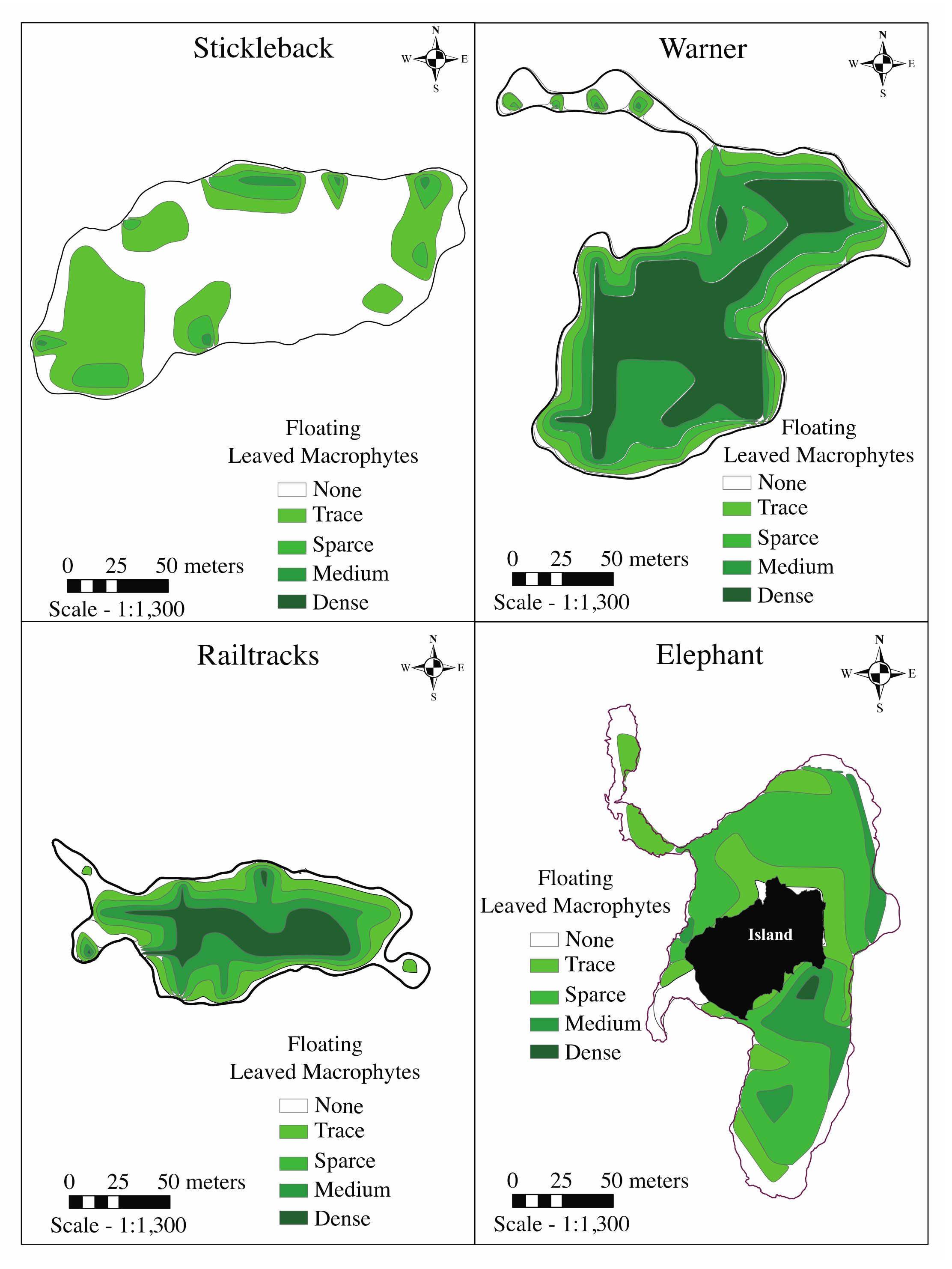
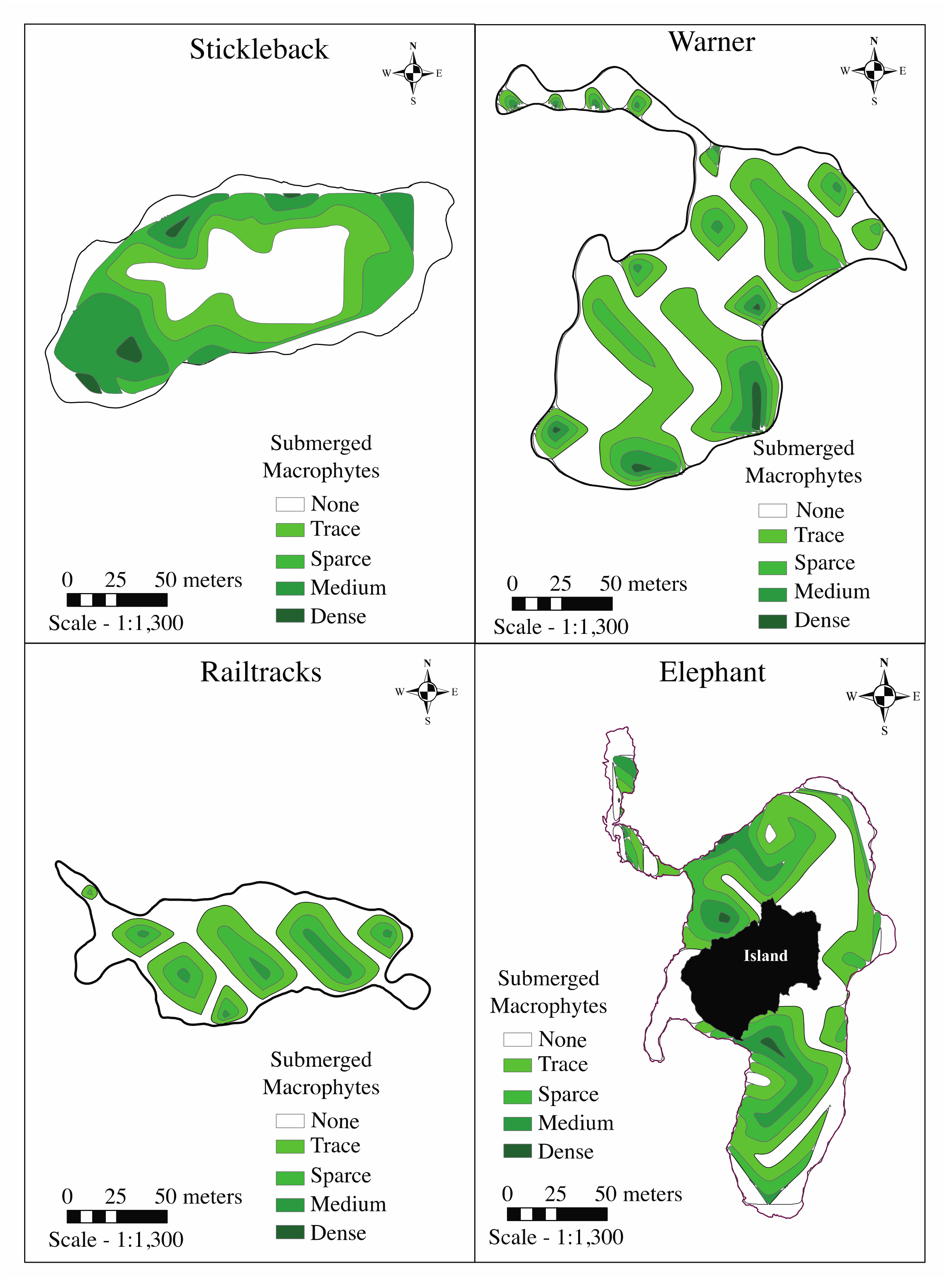
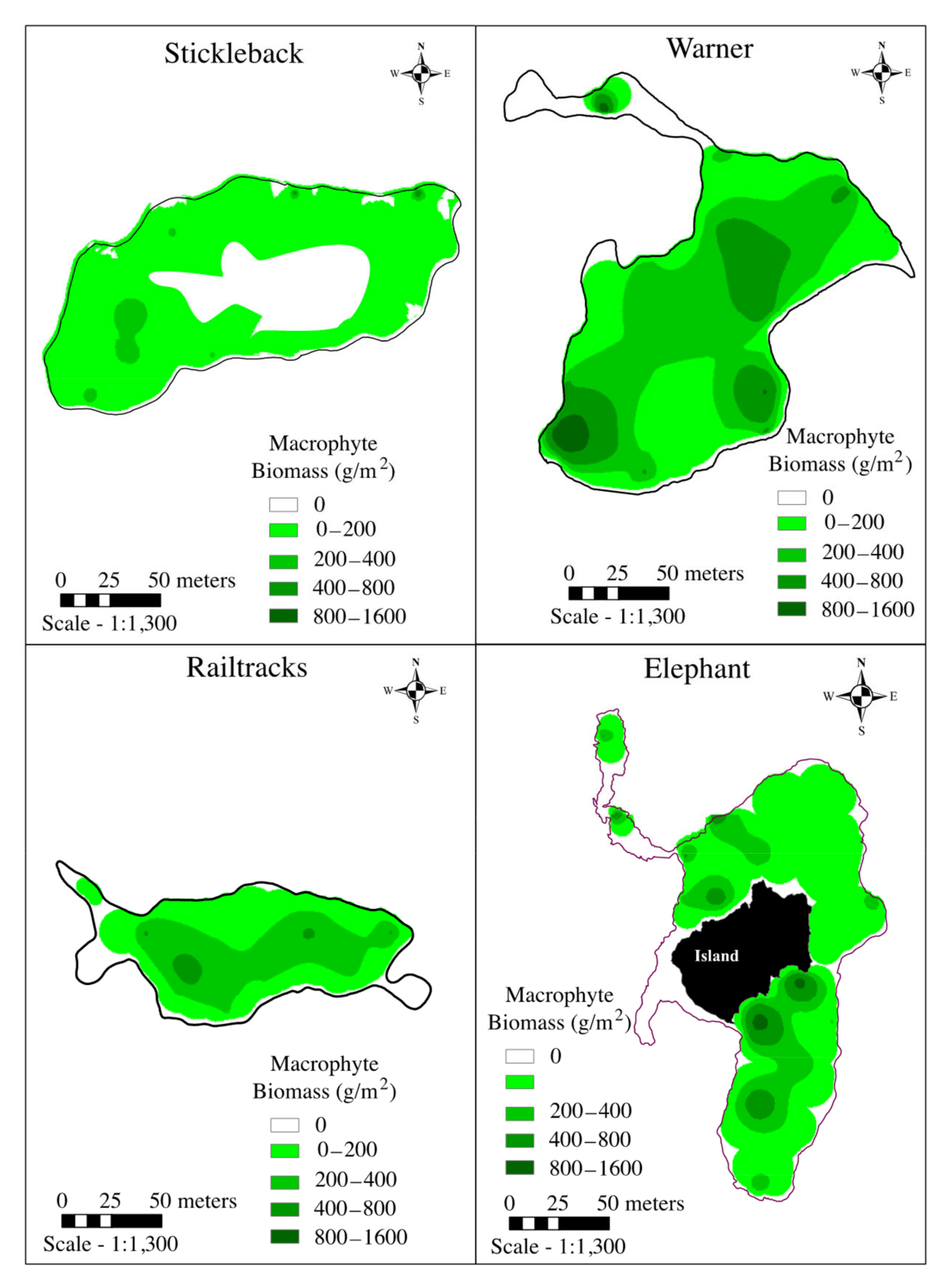
3.2.1. Macrophyte Abundance
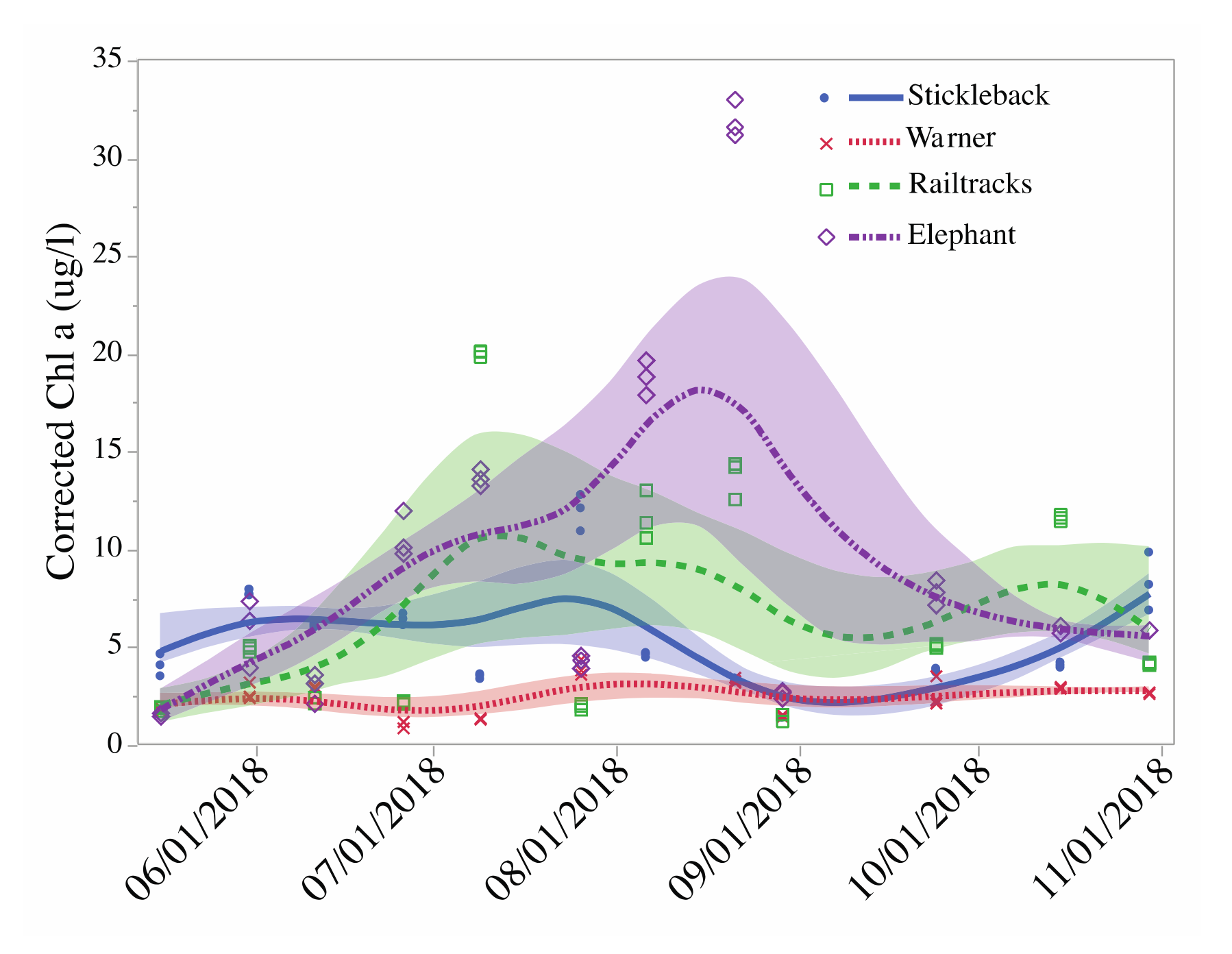
3.2.2. Phytoplankton
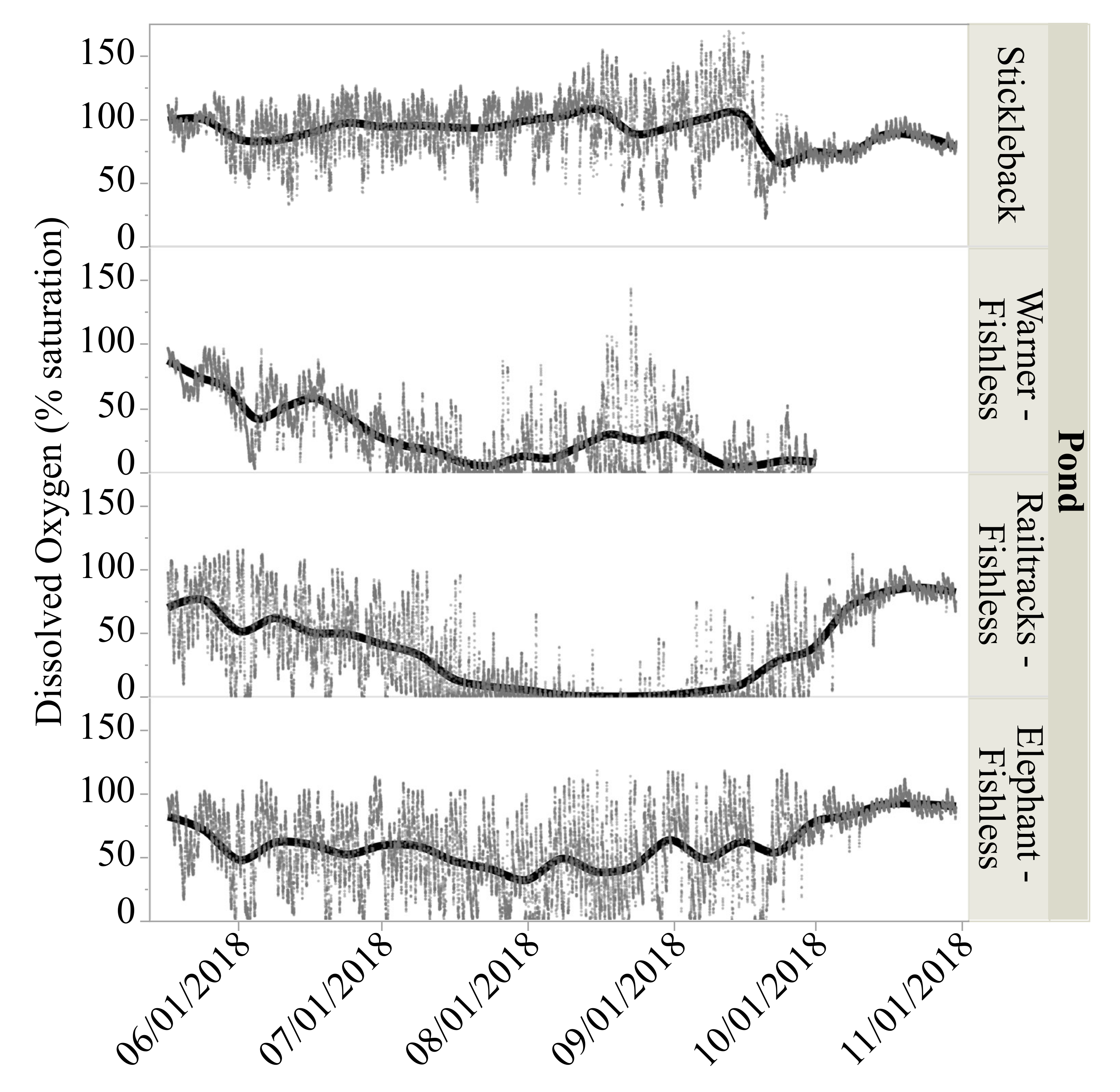
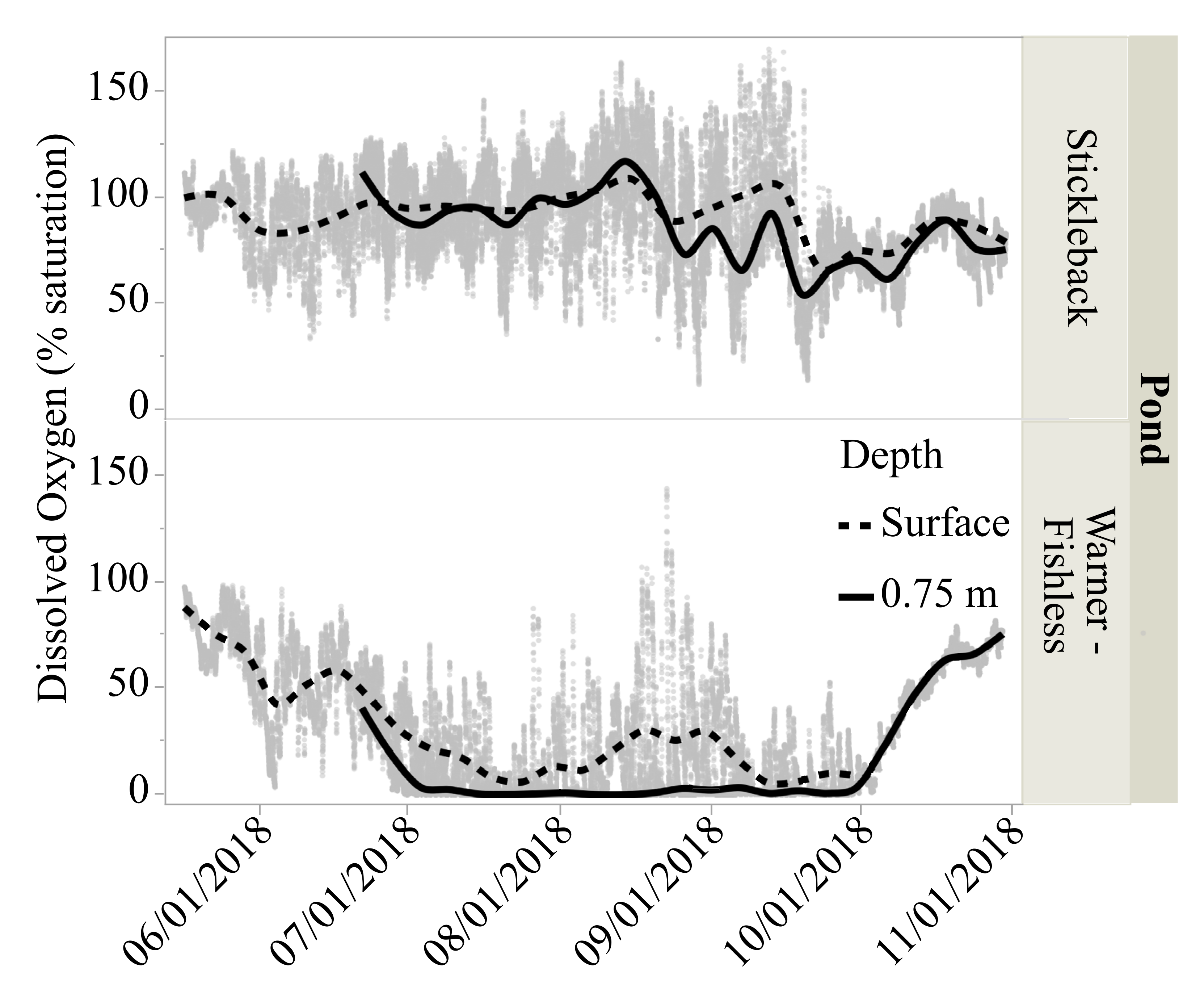
3.3. Dissolved Oxygen Levels
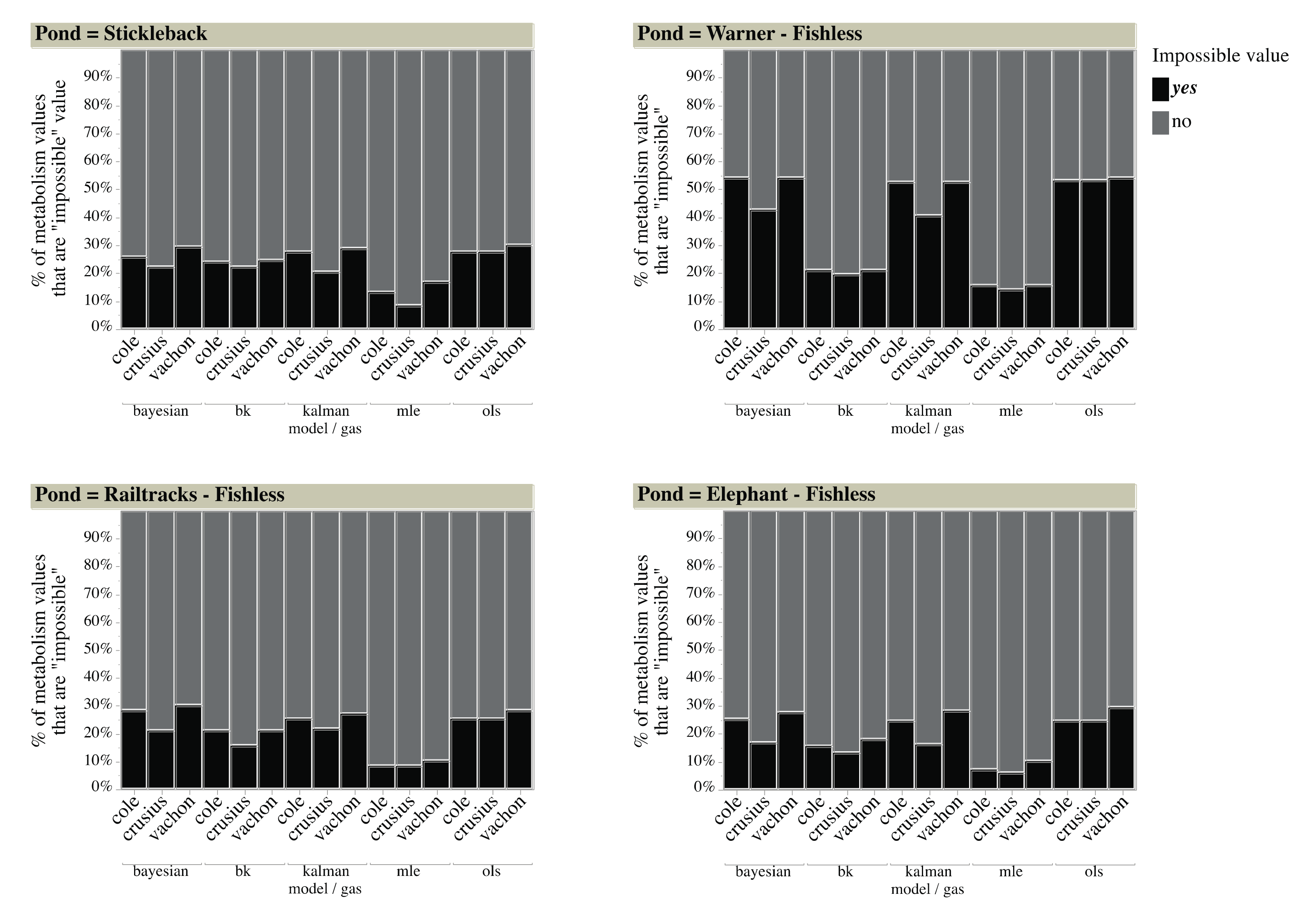
3.4. Pond Metabolism—Model Selection
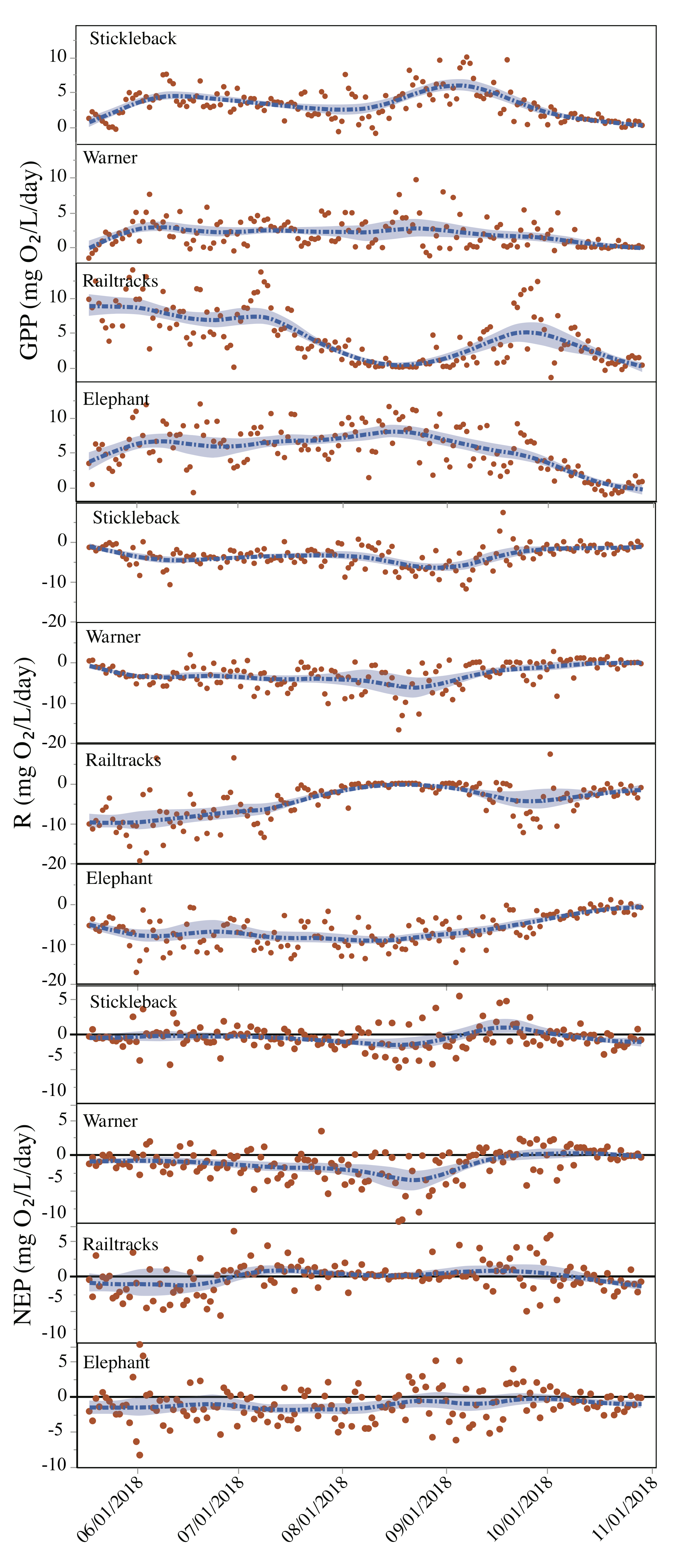
3.5. Pond Metabolism—Variation between Ponds and Depths
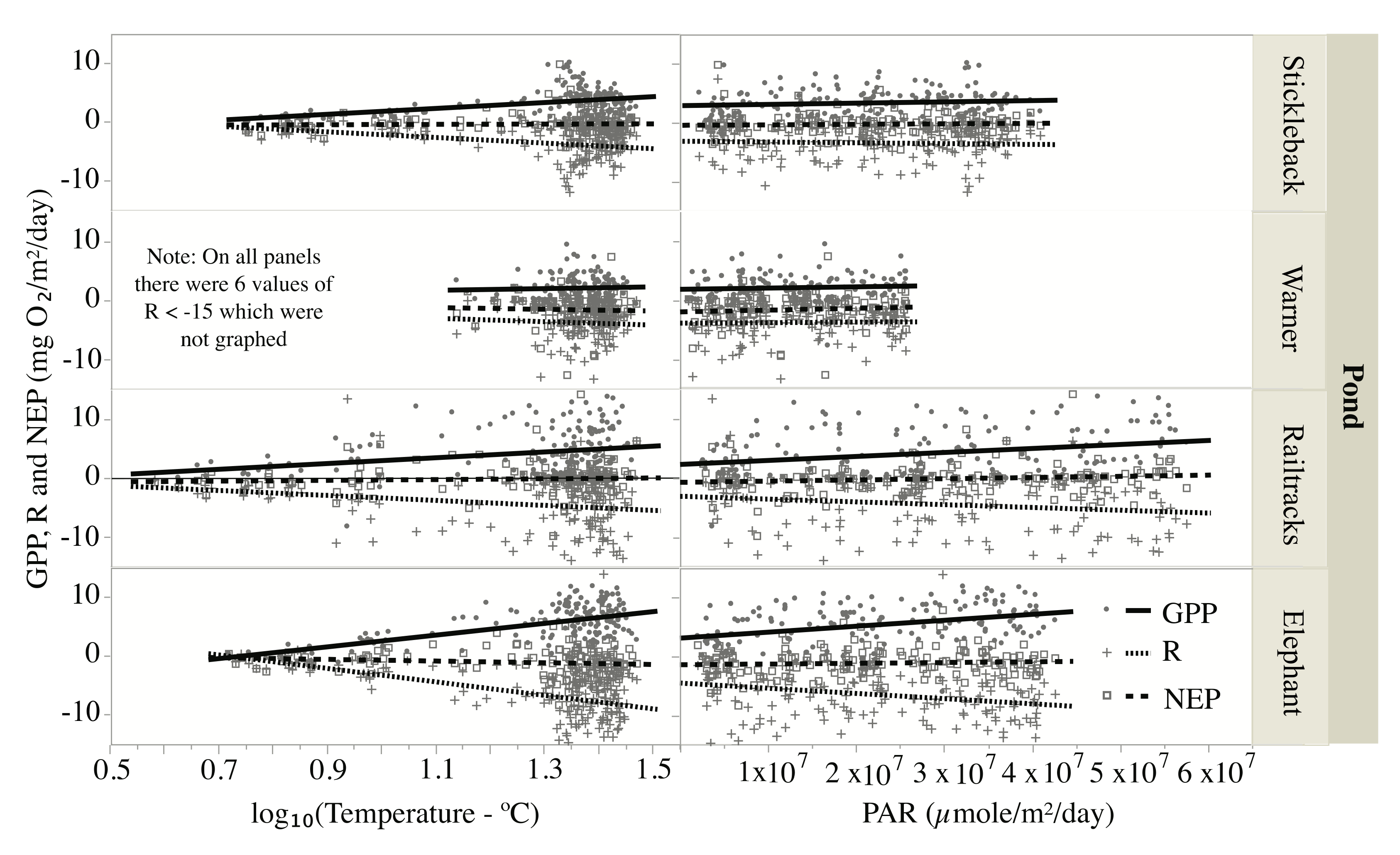
3.6. Pond Metabolism—Effects of Temperature and Light
4. Discussion
4.1. Physical/Chemical/Biologic Conditions of the Ponds
4.2. Ecosystem Metabolism
4.3. Issues with Measuring Ecosystem Metabolism
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Downing, J.A.; Prairie, Y.T.; Cole, J.J.; Duarte, C.M.; Tranvik, L.J.; Striegl, R.G.; McDowell, W.H.; Kortelainen, P.; Caraco, N.F.; Melack, J.M.; et al. The global abundance and size distribution of lakes, ponds, and impoundments. Limnol. Oceanogr. 2006, 51, 2388–2397. [Google Scholar] [CrossRef] [Green Version]
- Downing, J.A.; Duarte, C.M. Abundance and size distribution of lakes, ponds, and impoundments. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Elsevier: Oxford, UK, 2009; pp. 469–478. [Google Scholar]
- Downing, J.A. Emerging global role of small lakes and ponds: Little things mean a lot. Limnetica 2010, 29, 9–24. [Google Scholar] [CrossRef]
- Riley, W.D.; Potter, E.C.E.; Biggs, J.; Collins, A.L.; Jarvie, H.P.; Jones, J.I.; Kelly-Quinn, M.; Ormerod, S.J.; Sear, D.A.; Wilby, R.L.; et al. Small water bodies in UK and Ireland: Ecosystem function, human-generated degradation, and options for restorative action. Sci. Total Environ. 2018, 645, 1598–1616. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Xu, P.; Wang, Y.; Yan, K.; Chaudhary, S. Assessment of the ecosystem services provided by ponds in hilly areas. Sci. Total Environ. 2018, 642, 979–987. [Google Scholar] [CrossRef]
- Lougheed, V.L.; McIntosh, M.D.; Parker, C.A.; Stevenson, R.J. Wetland degradation leads to homogenization of the biota at local and landscape scales. Freshwater Biol. 2008, 53, 2402–2413. [Google Scholar] [CrossRef]
- Cole, J.J.; Prairie, Y.T.; Caraco, N.F.; McDowell, W.H.; Tranvik, L.J.; Striegl, R.G.; Duarte, C.M.; Kortelainen, P.; Downing, J.A.; Middelburg, J.J.; et al. Plumbing the global carbon cycle: Integrating inland waters into the terrestrial carbon budget. Ecosystems 2007, 10, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, P.J.; Cooke, D.A.; Deary, M.; Taylor, S.; Jeffries, M.J. Quantifying rapid spatial and temporal variations of CO2 fluxes from small, lowland freshwater ponds. Hydrobiologia 2017, 793, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.; Gilbert, P.J.; Cooke, D.A.; Deary, M.E.; Jeffries, M.J. High carbon burial rates by small ponds in the landscape. Front. Ecol. Environ. 2019, 17, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Dean, D.C.; He, F. Loss of only the smallest patches will reduce species diversity in most discrete habitat networks. Glob. Change Biol. 2018, 24, 5802–5814. [Google Scholar] [CrossRef]
- Bolpagni, R.; Poikane, S.; Laini, A.; Bagella, S.; Bartoli, M.; Cantonati, M. Ecological and conservation value of small standing-water ecosystems: A systematic review of current knowledge and future challenge. Water 2019, 11, 402. [Google Scholar] [CrossRef] [Green Version]
- Gee, J.H.R.; Smith, B.D.; Lee, K.M.; Griffiths, S.W. The ecological basis of freshwater pond management for biodiversity. Aquat. Conserv. 1997, 7, 91–104. [Google Scholar] [CrossRef]
- Thornhill, I.; Batty, L.; Death, R.G.; Friberg, N.R.; Ledger, M.E. Local and landscape scale determinants of macroinvertebrate assemblages and their conservation value in ponds across an urban land-use gradient. Biodivers. Conserv. 2017, 26, 1065–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wang, D. Responses of decomposition rate and nutrient release of floating-leaved and submerged aquatic macrophytes to vertical locations in an urban lake (Nanhu Lake, China). Chem. Ecol. 2019, 35, 431–444. [Google Scholar] [CrossRef]
- Parr, L.B.; Perkins, R.G.; Mason, C.F. Reduction in photosynthetic efficiency of Cladophora glomerata, induced by overlying canopies of Lemna spp. Water Res. 2002, 36, 1735–1742. [Google Scholar] [CrossRef]
- Klančnik, K.; Iskra, I.; Gradinjan, D.; Gaberščik, A. The quality and quantity of light in the water column are altered by the optical properties of natant plant species. Hydrobiologia 2018, 812, 203–212. [Google Scholar] [CrossRef]
- Smolders, A.J.P.; Vergeer, L.H.T.; van der Velde, G.; Roelofs, J.G.M. Phenolic contents of submerged, emergent and floating leaves of aquatic and semi-aquatic macrophyte species: Why do they differ? Oikos 2000, 91, 307–310. [Google Scholar] [CrossRef]
- Cronk, J.K.; Fennessy, M.S. Wetland Plants—Biology and Ecology; Lewis Publisher: London, UK, 2001. [Google Scholar]
- Ribaudo, C.; Bartoli, M.; Longhi, D.; Castaldi, S.; Neubauer, S.C.; Viaroli, P. CO2 and CH4 fluxes across a Nuphar lutea (L.) Sm. Stand. J. Limnol. 2012, 71, 200–210. [Google Scholar] [CrossRef] [Green Version]
- Grasset, C.; Abril, G.; Guillard, L.; Delolme, C.; Bornette, G. Carbon emission along a eutrophication gradient in temperate riverine wetlands: Effect of primary productivity and plant community composition. Freshwater Biol. 2016, 61, 1405–1420. [Google Scholar] [CrossRef]
- Schlacher, T.A.; Cronin, G. A trophic cascade in a macrophyte-based food web at the land-water ecotone. Ecol. Res. 2007, 5, 749–755. [Google Scholar] [CrossRef]
- Apolinarska, K.; Obremska, M.; Aunina, L.; Gałka, M. Response of the aquatic plants and mollusc communities in Lake Kojle (central Europe) to climatic changes between 250 BCE and 1550 CE. Aquat. Bot. 2018, 148, 35–45. [Google Scholar] [CrossRef]
- Bornette, G.; Puijalon, S. Response of aquatic plants to abiotic factors: A review. Aquat. Sci. 2011, 73, 1–14. [Google Scholar] [CrossRef]
- Scheffer, M.; Hosper, S.H.; Meijer, M.L.; Moss, B.; Jeppesen, E. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef]
- Braig, E.C.; Johnson, D.L. Impact of black bullhead (Ameiurus melas) on turbidity in a diked wetland. Hydrobiologia 2003, 490, 11–21. [Google Scholar] [CrossRef]
- Peretyatko, A.; Teissier, S.; Symoens, J.J.; Triest, L. Phytoplankton biomass and environmental factors over a gradient of clear to turbid peri-urban ponds. Aquatic Conser. 2007, 17, 584–601. [Google Scholar] [CrossRef]
- Potthoff, A.J.; Herwig, B.R.; Hanson, M.A.; Zimmer, K.D.; Butler, M.G.; Reed, J.R.; Parsons, B.G.; Ward, M.C. Cascading food-web effects of piscivore introductions in shallow lakes. J. Appl. Ecol. 2008, 45, 1170–1179. [Google Scholar] [CrossRef]
- Feuchtmayr, H.; Moran, R.; Hatton, K.; Connor, L.; Heyes, T.; Moss, B.; Harvey, I.; Atkinson, D. Global warming and eutrophication: Effects on water chemistry and autotrophic communities in experimental hypertrophic shallow lake mesocosms. J. Appl. Ecol. 2009, 46, 713–723. [Google Scholar] [CrossRef]
- Saad, J.F.; Porcel, S.; Lancelotti, J.; O’Farrell, I.; Izaguirre, I. Both lake regime and fish introduction shape autotrophic planktonic communities of lakes from the Patagonian Plateau (Argentina). Hydrobiologia 2019, 831, 133–145. [Google Scholar] [CrossRef]
- Staehr, P.; Sand-Jensen, K. Temporal dynamics and regulation of lake metabolism. Limnol. Oceanogr. 2007, 52, 108–120. [Google Scholar] [CrossRef] [Green Version]
- Stefanidis, K.; Dimitriou, E. Differentiation in aquatic metabolism between littoral habitats with floating-leaved and submerged macrophyte growth forms in a shallow eutrophic lake. Water 2019, 11, 287. [Google Scholar] [CrossRef] [Green Version]
- Van de Bogert, M.C.; Bade, D.L.; Carpenter, S.R.; Cole, J.J.; Pace, M.L.; Hanson, P.C.; Langman, O.C. Spatial heterogeneity strongly affects estimates of ecosystem metabolism in two north temperate lakes. Limnol. Oceanogr. 2012, 57, 1689–1700. [Google Scholar] [CrossRef] [Green Version]
- Klotz, R.L. Factors driving the metabolism of two north temperate ponds. Hydrobiologia 2013, 711, 9–17. [Google Scholar] [CrossRef]
- Hornbach, D.J.; Hove, M.C.; Ensley-Field, M.W.; Glasenapp, M.R.; Goodbar, I.A.; Harman, J.D.; Huber, B.D.; Kangas, E.A.; Liu, K.X.; Stark-Ragsdale, M.; et al. Comparison of ecosystem processes in a woodland and prairie pond with different hydroperiods. J. Freshwater Ecol. 2017, 32, 675–695. [Google Scholar] [CrossRef] [Green Version]
- Kazanjian, G.; Flury, S.; Attermeyer, K.; Kalettka, T.; Andreas Kleeberg, A.; Premke, K.; Köhler, J.; Hilt, S. Primary production in nutrient-rich kettle holes and consequences for nutrient and carbon cycling. Hydrobiologia 2018, 806, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Caraco, N.; Cole, J.; Findlay, S.; Wigand, C. Vascular plants as engineers of oxygen in aquatic systems. BioScience 2006, 56, 219–225. [Google Scholar] [CrossRef]
- Zimmer, K.D.; Hobbs, W.O.; Domin, L.M.; Herwig, B.R.; Hanson, M.A.; Cotner, J.B. Uniform carbon fluxes in shallow lakes in alternative stable states. Limnol. Oceanogr. 2016, 61, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Ameel, J.J.; Axler, R.P.; Owen, C.J. Persulfate digestion for determination of total nitrogen and phosphorus in low-nutrient waters. Am. Environ. Lab. 1993, 10, 7–11. [Google Scholar]
- Madsen, J.D.; Wersal, R.M. A review of aquatic plant monitoring and assessment methods. J. Aquat. Plant Manage. 2017, 55, 1–12. [Google Scholar]
- Arar, E.J.; Collins, G.B. In vitro Determination of Chlorophyll a and Pheophytin a in Marine and Freshwater Algae by Fluorescence; National Exposure Research Laboratory Office of Research and Development U.S. Environmental Protection Agency (EPA Publication; Method No. 445.0): Cincinnati, OH, USA, 1997. Available online: http://permanent.access.gpo.gov/lps68140/m445-0.pdf (accessed on 15 January 2020).
- Hagerthey, S.E.; Cole, J.J.; Kilbanea, D. Aquatic metabolism in the Everglades: Dominance of water column heterotrophy. Limnol. Oceanogr. 2010, 55, 653–666. [Google Scholar] [CrossRef]
- Odum, H.T. Primary production in flowing waters. Limnol. Oceanogr. 1956, 1, 102–117. [Google Scholar] [CrossRef]
- Winslow, L.A.; Zwart, J.A.; Batt, R.D.; Dugan, H.A.; Woolway, R.I.; Corman, J.R.; Hanson, P.C.; Read, J.S. LakeMetabolizer: An R package for estimating lake metabolism from free-water oxygen using diverse statistical models. Inland Waters 2016, 6, 622–636. [Google Scholar] [CrossRef]
- Cole, J.J.; Caraco, N.F. Atmospheric exchange of carbon dioxide in a low-wind oligotrophic lake measured by the addition of SF6. Limnol. Oceanogr. 1998, 43, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Crusius, J.; Wanninkhof, R. Gas transfer velocities measured at low wind speed over a lake. Limnol. Oceanogr. 2003, 48, 1010–1017. [Google Scholar] [CrossRef]
- Vachon, D.; Prairie, Y. The ecosystem size and shape dependence of gas transfer velocity versus wind speed relationships in lakes. Can. J. Fish. Aquat. Sci. 2013, 70, 1757–1764. [Google Scholar] [CrossRef]
- Heiskary, S.; Wilson, B. Minnesota’s approach to lake nutrient criteria development. Lake Reserv. Manage. 2008, 24, 282–297. [Google Scholar] [CrossRef]
- Søndergaard, M.; Jeppesen, E.; Jensen, J.P. Pond or lake: Does it make any difference? Arch. Hydrobiol. 2005, 162, 143–165. [Google Scholar] [CrossRef]
- Zimmer, K.D.; Hanson, M.A.; Herwig, B.R.; Konsti, M.L. Threshold and stability of alternative regimes in shallow Prairie-Parkland lakes of central North America. Ecosystems 2009, 12, 843–852. [Google Scholar] [CrossRef]
- Daldorph, P.W.G.; Thomas, J.D. Factors influencing the stability of nutrient-enriched fresh-water macrophyte communities—The role of sticklebacks Pungitius pungitius and fresh-water snails. Freshwater Biol. 1995, 33, 271–289. [Google Scholar] [CrossRef]
- Stephen, D.; Moss, B.; Phillips, G. The relative importance of top-down and bottom-up control of phytoplankton in a shallow macrophyte-dominated lake. Freshwater Biol. 1998, 39, 699–713. [Google Scholar] [CrossRef]
- Horppila, J.; Nurminen, L. Effects of different macrophyte growth forms on sediment and P resuspension in a shallow lake. Hydrobiologia 2005, 545, 167–175. [Google Scholar] [CrossRef]
- Hilt, S.; Brothers, S.; Jeppesen, E.; Veraart, A.J.; Kosten, S. Translating regime shifts in shallow lakes to changes in ecosystem functions and services. Bioscience 2017, 67, 928–936. [Google Scholar] [CrossRef]
- Gross, E.M. Allelopathy of aquatic autotrophs. Crit. Rev. Plant Sci. 2003, 22, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Schriver, P.; Bogestrand, J.; Jeppesen, E.; Sondergaard, M. Impact of submerged macrophytes on fish-zooplankton-phytoplankton interactions—Large-scale enclosure experiments in a shallow eutrophic lake. Freshwater Biol. 1995, 33, 255–270. [Google Scholar] [CrossRef]
- Burks, R.L.; Lodge, D.M.; Jeppesen, E.; Lauridsen, T. Diel horizontal migration of zooplankton: Costs and benefits of inhabiting littoral zones. Freshwater Biol. 2002, 47, 343–365. [Google Scholar] [CrossRef]
- Muylaert, K.; Pérez-Martínez, C.; Sánchez-Castillo, P.; Lauridsen, T.L.; Vanderstukken, M.; Declerck, S.A.J.; Van der Gucht, K.; Conde-Porcuna, J.; Jeppesen, E.; De Meester, L.; et al. Influence of nutrients, submerged macrophytes and zooplankton grazing on phytoplankton biomass and diversity along a latitudinal gradient in Europe. Hydrobiologia 2010, 653, 79–90. [Google Scholar] [CrossRef]
- López-Archilla, A.I.; Mollá, S.; Coleto, M.C.; Guerrero, M.C.; Montes, C. Ecosystem metabolism in a Mediterranean shallow lake (Laguna de Santa Olalla, Doñana National Park, SW Spain. Wetlands 2004, 24, 848–858. [Google Scholar] [CrossRef]
- Geertz-Hansen, O.; Montes, C.; Duarte, C.M.; Sand-Jensen, K.; Marbá, N.; Grillas, P. Ecosystem metabolism in a temporary Mediterranean marsh (Doñana National Park, SW Spain). Biogeosciences 2011, 8, 963–971. [Google Scholar] [CrossRef] [Green Version]
- Christensen, J.P.A.; Sand-Jensen, K.; Staehr, P.A. Fluctuating water levels control water chemistry and metabolism of a charophyte-dominated pond. Freshwater Biol. 2013, 58, 1353–1365. [Google Scholar] [CrossRef]
- Solomon, C.T.; Bruesewitz, D.A.; Richardson, D.C.; Rose, K.C.; Van de Bogert, M.C.; Hanson, P.C.; Kratz, T.K.; Larget, B.; Adrian, R.; Babin, B.L.; et al. Ecosystem respiration: Drivers of daily variability and background respiration in lakes around the globe. Limnol. Oceanogr. 2013, 58, 849–866. [Google Scholar] [CrossRef] [Green Version]
- Lauster, G.H.; Hanson, P.C.; Kratz, T.K. Gross primary production and respiration differences among littoral and pelagic habitats in northern Wisconsin lakes. Can. J. Fish. Aquat. Sci. 2006, 63, 1130–1141. [Google Scholar] [CrossRef]
- Coloso, J.J.; Cole, J.J.; Pace, M.L. Difficulty in discerning drivers of lake ecosystem metabolism with high-frequency data. Ecosystems 2011, 14, 935–948. [Google Scholar] [CrossRef]
- Hoellein, T.J.; Bruesewitz, D.A.; Richardson, D.C. Revisiting Odum (1956): A synthesis of aquatic ecosystem metabolism. Limnol. Oceanogr. 2013, 58, 2089–2100. [Google Scholar] [CrossRef]
- Hanson, P.C.; Bade, D.L.; Carpenter, S.R. Lake metabolism: Relationships with dissolved organic carbon and phosphorus. Limnol. Oceanogr. 2003, 48, 1112–1119. [Google Scholar] [CrossRef] [Green Version]
- Staehr, P.A.; Baastrup-Spohr, L.; Sand-Jensen, K.; Stedmon, C. Lake metabolism scales with lake morphometry and catchment conditions. Aquat. Sci. 2011, 74, 155–169. [Google Scholar] [CrossRef]
- Laas, A.; Nõges, P.; Kõiv, T.; Nõges, T. High-frequency metabolism study in a large and shallow temperate lake reveals seasonal switching between net autotrophy and net heterotrophy. Hydrobiologia 2012, 694, 57–74. [Google Scholar] [CrossRef]
- Camacho, A.; Murueta, N.; Blasco, E.; Santamans, A.C.; Picazo, A. Hydrology-driven macrophyte dynamics determines the ecological functioning of a model Mediterranean temporary lake. Hydrobiologia 2016, 774, 93–107. [Google Scholar] [CrossRef]
- Sand-Jensen, K.; Staehr, P.A. Scaling of pelagic metabolism to size, trophy and forest cover in small Danish lakes. Ecosystems 2007, 10, 127–141. [Google Scholar] [CrossRef]
- Sand-Jensen, K.; Staehr, P.A. Net heterotrophy in lakes: A widespread over gradients in trophic and land cover. Ecosystems 2009, 12, 336–348. [Google Scholar] [CrossRef]
- Àvila, N.; López-Flore, R.; Boix, D.; Gascón, S.; Quintana, X.D. Environmental factors affecting the balance of autotrophs versus heterotrophs in the microbial food web of temporary ponds. Hydrobiologia 2016, 782, 127–143. [Google Scholar] [CrossRef]
- Martinsen, K.T.; Kragh, T.; Sand-Jensen, K. Carbon dioxide efflux and ecosystem metabolism of small forest lakes. Aquatic Sci. 2020, 82, 9. [Google Scholar] [CrossRef]
- Martinsen, K.T.; Andersen, M.R.; Kragh, T.; Sand-Jensen, K. High rates and close diel coupling of primary production and ecosystem respiration in small, oligotrophic lakes. Aquat. Sci. 2017, 79, 995–1007. [Google Scholar] [CrossRef]
- Yvon-Durocher, G.; Jones, J.I.; Trimmer, M.; Woodward, G.; Montoya, J.M. Warming alters the metabolic balance of ecosystems. Phil. Trans. R. Soc. B 2010, 365, 2117–2126. [Google Scholar] [CrossRef] [Green Version]
- Dunalska, J.A.; Staehr, P.A.; Jaworska, B.; Górnaik, D. Ecosystem metabolism in a lake restored by hypolimnetic withdrawal. Ecol. Eng. 2014, 73, 616–623. [Google Scholar] [CrossRef]
- Peeters, F.; Atamanchuk, D.; Tengberg, A.; Encinas-Fernandez, J.; Hofmann, H. Lake Metabolism: Comparison of Lake Metabolic Rates Estimated from a Diel CO2- and the Common Diel O2-Technique. PLoS ONE 2016, 11, e0168393. [Google Scholar] [CrossRef]
- Żbikowski, J.; Simčič, T.; Pajk, F.; Poznańska-Kakareko, M.; Kakareko, T.; Kobak, J. Respiration rates in shallow lakes of different types: Contribution of benthic microorganisms, macrophytes, plankton and macrozoobenthos. Hydrobiologia 2019, 828, 117–136. [Google Scholar] [CrossRef] [Green Version]
- Dugan, H.A.; Woolway, R.I.; Santoso, A.B.; Corman, J.R.; Jamines, A.; Nodine, E.R.; Patil, V.P.; Zwart, J.A.; Brentrup, J.A.; Hetherington, A.L.; et al. Consequences of gas flux model choice on the interpretation of metabolic balance across 15 lakes. Inland Waters 2016, 6, 581–592. [Google Scholar] [CrossRef]
- Holgerson, M.A.; Farr, E.R.; Raymond, P.A. Gas transfer velocities in small forested ponds. J. Geophys. Res. Biogeo. 2017, 122, 1011–1021. [Google Scholar] [CrossRef]
- Istvánovics, V.; Honti, M. Coupled simulation of high-frequency dynamics of dissolved oxygen and chlorophyll widens the scope of lake metabolism studies. Limnol. Oceanogr. 2018, 63, 72–90. [Google Scholar] [CrossRef] [Green Version]
- McEnroe, N.A.; Buttle, J.M.; Marsalek, J.; Pick, F.R.; Xenopoulos, M.A.; Frost, P.C. Thermal and chemical stratification of urban ponds: Are they ‘completely mixed reactors’? Urban Ecosyst. 2013, 16, 327–339. [Google Scholar] [CrossRef]
- Martinsen, K.T.; Andersen, M.R.; Sand-Jensen, K. Water temperature dynamics and the prevalence of daytime stratification in small temperate shallow lakes. Hydrobiologia 2019, 826, 247–262. [Google Scholar] [CrossRef]
- Andersen, M.R.; Kragh, T.; Sand-Jensen, K.K. Extreme diel dissolved oxygen and carbon cycles in shallow vegetated lakes. Proc. Roy. Soc. B 2017, 284, 1427. [Google Scholar] [CrossRef] [PubMed]
- Brothers, S.M.; Hilt, S.; Meyer, S.; Köhler, J. Plant community structure determines primary productivity in shallow, eutrophic lakes. Freshwater Biol. 2013, 58, 2264–2276. [Google Scholar] [CrossRef] [Green Version]
- Obrador, B.; Staehr, P.A.; Christensen, J.P.C. Vertical patterns of metabolism in three contrasting stratified lakes. Limnol. Oceanogr. 2014, 59, 1228–1240. [Google Scholar] [CrossRef]
- Rose, K.C.; Winslow, L.A.; Read, J.S.; Read, E.K.; Solomon, C.T.; Adrian, R.; Hanson, P.C. Improving the precision of lake ecosystem metabolism estimates by identifying predictors of model uncertainty. Limnol. Oceanogr. Meth. 2014, 12, 303–312. [Google Scholar] [CrossRef]
- Giling, D.P.; Staehr, P.A.; Grossart, H.P.; Andersen, M.R.; Boehrer, B.; Escot, C.; Evrendilek, F.; Gomez-Gener, L.; Honti, M.; Jones, I.D.; et al. Delving deeper: Metabolic processes in the metalimnion of stratified lakes. Limnol. Oceanogr. 2017, 62, 1288–1306. [Google Scholar] [CrossRef]
- Tonetta, D.; Staehr, P.A.; Schmitt, R.; Petrucio, M.M. Physical conditions driving the spatial and temporal variability in aquatic metabolism of a subtropical coastal lake. Limnologica 2016, 58, 30–40. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Pond | |||
|---|---|---|---|---|
| Stickleback | Warner | Railtracks | Elephant | |
| Average depth (m) | 1.5 | 1.2 | 0.47 | 0.57 |
| Pond surface area (m2) | 15,121 | 16,812 | 6995 | 7077 |
| Pond volume (m3) | 22,733 | 19,806 | 3301 | 7077 |
| Conductivity (µS/cm) 1 | 40.1 a (4.0) | 70.0 (7.7) b | 23.3 (11.2)c | 30.4 a,c (3.7) |
| Extinction coefficient—June 13, 2018 (r²) | 2.33 (0.91) | 2.72 (0.95) | 6.96 (0.98) | 2.96 (0.97) |
| Extinction coefficient—July 16, 2018 (r²) | 2.14 (0.93) | 3.99 (0.98) | 6.39 (0.94) | 2.56 (0.96) |
| Total phosphorus (µg/L) 2,4 | 89.4 (9.3) a | 46.5 (9.3) b | 44.7 (9.3)b | 56.5 (9.3) a,b |
| Total nitrogen (mg/L) 2,4 | 1.6 (0.2) a | 0.7 (0.2)b | 0.7 (0.2)b | 0.8 (0.2) b |
| Dissolved organic carbon (mg/L) 2,4 | 9.8 (0.5) a | 6.2 (0.5)b | 6.7 (0.5)b | 7.6 (0.5) c |
| Water temperature (°C) 2,3 | 21.3 (0.05) a | 20.3 (0.05) b | 20.1 (0.05)c | 20.7 (0.05) d |
| Average daily water temperature range (°C) 2,3 | 5.1 (0.23)a | 7.0 (0.23) b | 8.0 (0.23) c | 6.8 (0.23) b |
| Mean dissolved oxygen saturation (%) 2,3 | 90.6 (0.19) a | 31.0 (0.20)b | 35.2 (0.19) c | 58.9 (0.19) d |
| Independent Variables | PAR 1 (µmoles/m2/sec) 2 | Temperature (°C) 2 | Dissolved Oxygen 3 (% Saturation) 2 |
|---|---|---|---|
| Pond | 64.1; 3, 50; < 0.0001 | 320; 3, 102; < 0.0001 | 108.8; 3, 92; < 0.0001 |
| Date | 22.1; 1, 50; < 0.0001 | 87.1; 1, 102; < 0.0001 | 0.1; 1, 92; 0.8 |
| Depth | 471.9; 1, 50; < 0.0001 | 232.3; 1, 102; < 0.0001 | 104.9; 1, 92; < 0.0001 |
| Pond × Date | 24.0; 3, 50; < 0.0001 | 195.5; 3, 102; <0.0001 | 9.0; 3, 92; < 0.0001 |
| Date × Depth | 0.1; 1, 50; 0.7 | 3.6; 1, 102; 0.06 | 4.6; 1, 92; 0.04 |
| Pond × Date × Depth | 3.4; 3, 50; 0.03 | 9.2; 3, 102; < 0.0001 | 12.7; 3, 92; < 0.0001 |
| Parameter | Pond | |||
|---|---|---|---|---|
| Stickleback | Warner | Railtracks | Elephant | |
| Submerged macrophytes (% medium or dense coverage) | 20.7 | 6.1 | 3.3 | 8.4 |
| Floating-leaved macrophytes (% medium or dense coverage) | 0.9 | 61.2 | 43.7 | 12.4 |
| Total macrophyte biomass (kg) | 376.5 | 4129.2 | 1166.5 | 1487.5 |
| Average macrophyte biomass (g/m2) 1 | 24.9 (36.4)a | 229.3 (3186.9) b | 151.6 (117.3) c | 98.6 (141.7) d |
| Phytoplankton biomass (µg chl a/L) 2 | 5.34 (1.17) a | 2.6 (1.2)a | 6.65 (1.3) a,b | 9.3 (1.2) b |
| Mean GPP (mgO2/L/day) 1 | 3.2 (2.3) a | 2.2 (2.2)b | 4.3 (4.0) c | 5.5 (4.0) d |
| Mean R (mgO2/L/day) 1 | −3.6 (2.7)a | −3.7 (3.4) a | −4.4 (4.9) a | −6.8 (6.1) b |
| Mean NEP (mgO2/L/day) 1 | −0.4 (1.8) a | −1.5 (2.4)b | −0.1 (2.6) a | −1.3 (3.3) b |
| Category of Comparison | Independent Variables | Dependent Variable F; df; p | ||
|---|---|---|---|---|
| GPP | R | NEP | ||
| Temperature | Pond | 30.9; 3626; < 0.0001 | 21.6; 3, 626; < 0.0001 | 8.9; 3626; < 0.0001 |
| log10 (temperature °C) | 23.0; 1626; < 0.0001 | 15.5; 1626; < 0.0001 | 0.3; 1626; 0.58 | |
| Pond × Temperature | 5.4; 3626; 0.001 | 5.4; 3626; 0.001 | 1.1; 3626; 0.34 | |
| Light | Pond | 20.5; 3626; < 0.0001 | 17.6; 3626; < 0.0001 | 5.7; 3, 626; 0.0008 |
| PAR (µmoles/m2/sec) | 19.7; 1626; < 0.0001 | 4.9; 1626; 0.027 | 3.7; 1626; 0.055 | |
| Pond × Temperature | 3.1; 3626; 0.026 | 1.8; 3626; 0.14 | 0.2; 3626; 0.88 | |
| Study [Reference] | GPP (mgO2/L/day) | R (mgO2/L/day) | NEP (mgO2/L/day) |
|---|---|---|---|
| López-Archilla et al. [58] | 2.9 | −3.2 | −0.3 |
| Geertz-Hansen et al. [59] 1 | 0.9–9.1 | −4.1–−0.4 | 0.5–5.0 |
| Christensen et al. [60] | 3.1 | −2.9 | 0.3 |
| Klotz [33] 2 | 3.6–7.1 | −5.3–−4.2 | −0.6–1.62 |
| Solomon et al. [61] 3 | 0–15 | 0–−15 | −0.38 |
| Hornbach et al. [34] 4 | 5.0–8.8 | −48.0–−38.9 | −39.2–−33.9 |
| This Study 5 | 2.2–5.5 | −6.8–−3.6 | −1.5–−0.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hornbach, D.J.; Schilling, E.G.; Kundel, H. Ecosystem Metabolism in Small Ponds: The Effects of Floating-Leaved Macrophytes. Water 2020, 12, 1458. https://doi.org/10.3390/w12051458
Hornbach DJ, Schilling EG, Kundel H. Ecosystem Metabolism in Small Ponds: The Effects of Floating-Leaved Macrophytes. Water. 2020; 12(5):1458. https://doi.org/10.3390/w12051458
Chicago/Turabian StyleHornbach, Daniel J., Emily G. Schilling, and Holly Kundel. 2020. "Ecosystem Metabolism in Small Ponds: The Effects of Floating-Leaved Macrophytes" Water 12, no. 5: 1458. https://doi.org/10.3390/w12051458
APA StyleHornbach, D. J., Schilling, E. G., & Kundel, H. (2020). Ecosystem Metabolism in Small Ponds: The Effects of Floating-Leaved Macrophytes. Water, 12(5), 1458. https://doi.org/10.3390/w12051458