An Extended Ecosystem Model for Understanding EE2 Indirect Effects on a Freshwater Food Web and its Ecosystem Function Resilience



Abstract
:
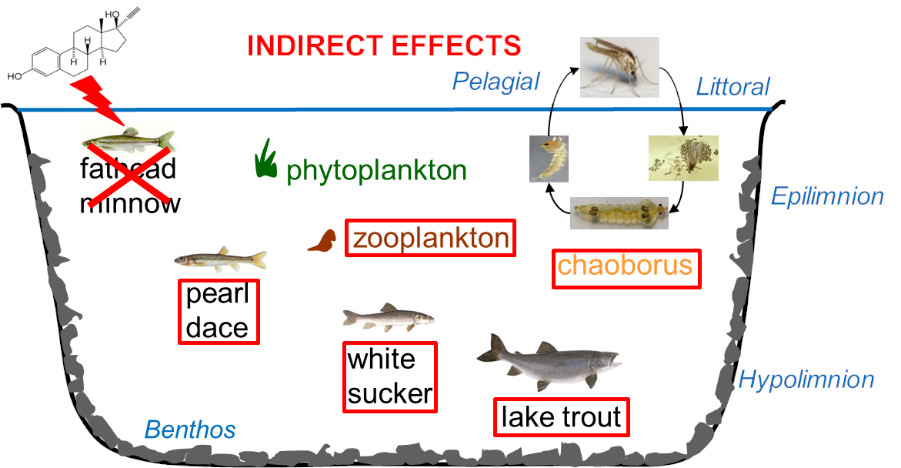{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Step 1: Collecting Experimental Data
2.2. Step 2: Building and Calibrating an Ecosystem Model
2.3. Step 3: Simulating Multitrophic Interactions
2.4. Step 4: Predicting EE2 Indirect Effects
3. Results
3.1. Extended Ecosystem Model
3.2. Seasonal Cycles, Multitrophic Interactions and Biodiversity
3.2.1. Primary Producers—Phytoplankton
Seasonal Cycles and Multitrophic Interactions
Horizontal Diversity
Vertical Diversity
3.2.2. Primary Consumers—Zooplankton
Seasonal Cycles and Multitrophic Interactions
Horizontal Diversity
Vertical Diversity
3.2.3. Secondary and Tertiary Consumers—Fish
Seasonal Cycles and Multitrophic Interactions
Horizontal Diversity
Vertical Diversity
3.2.4. Omnivorous, Mid-Trophic Level Group—Chaoborus
Seasonal Cycles and Multitrophic Interactions
Vertical Diversity
3.3. Indirect Effects of EE2
4. Discussion
4.1. Chaoborus Contribution to Ecosystem Functioning
4.2. EE2 Effects on Ecosystem Functioning
4.3. In the Context of Ecosystem-Based Management (EBM)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Decision, E.U. Watch List of Decision 495/2015/EU. Watch list of substances for Union-wide monitoring in the field of water policy pursuant to Directive 2008/105/EC of the European Parliament and of the Council. Off. J. Eur. Union 2015, 78, 40–42. [Google Scholar]
- Sousa, J.C.G.; Ribeiro, A.R.; Barbosa, M.O.; Pereira, M.F.R.; Silva, A.M.T. A review on environmental monitoring of water organic pollutants identified by EU guidelines. J. Hazard. Mater. 2018, 344, 146–162. [Google Scholar] [CrossRef] [PubMed]
- Han Tran, N.; Reinhard, M.; Yew-Hoong, K. Occurrence and fate of emerging contaminants in municipal wastewater treatment plants from different geographical regions—A Review. Water Res. 2018, 133, 182–207. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, D.J.; Mastrocco, F.; Anderson, P.D.; Lange, R.; Sumpter, J.P. Predicted–no–effect concentrations for the steroid estrogens estrone, 17b-estradiol, estriol and 17a-ethinylestradiol. Environ. Toxicol. Chem. 2012, 31, 1396–1406. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.R.; Gunnarsson, L.; Kristiansson, E.; Tyler, C.R. Assessing variation in the potential susceptibility of fish to pharmaceuticals, considering evolutionary differences in their physiology and ecology. Phil. Trans. R. Soc. B 2014, 369, 20130576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleeger, J.W.; Carmana, K.R.; Nisbet, R.M. Indirect effects of contaminants in aquatic ecosystems. Sci. Total Environ. 2003, 317, 207–233. [Google Scholar] [CrossRef]
- Peters, R.E.M.; Courtenay, S.C.; Hewitt, L.M.; Maclatchy, D.L. Effects of 17α-ethynylestradiol on early-life development, sex differentiation and vitellogenin induction in mummichog (Fundulus heteroclitus). Mar. Environ. Res. 2010, 69, 178–186. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, B.M.; Lazorchak, J.M.; Jensen, K.M.; Haring, H.J.; Smith, M.E.; Flick, R.W.; Bencic, D.C.; Biales, A.D. Reproductive effects in fathead minnows (Pimphales promelas) following a 21-d exposure to 17α-ethinylestradiol. Chemosphere 2016, 144, 366–373. [Google Scholar] [CrossRef]
- Young, B.J.; Lopez, G.C.; Cristos, D.S.; Crespo, D.C.; Somoza, G.M.; Carriquiriborde, P. Intersex and liver alterations induced by long-term sublethal exposure to 17alpha-ethinylestradiol in adult male cnesterodon decemmaculatus (Pisces: Poeciliidae). Environ. Toxicol. Chem. 2017, 36, 1738–1745. [Google Scholar] [CrossRef] [Green Version]
- Porseryd, T.; Larsson, J.; Kellner, M.; Bollner, T.; Dinnétz, P.; Hällström, I.P. Altered non-reproductive behavior and feminization caused by developmental exposure to 17α–ethinylestradiol persist to adulthood in three-spined stickleback (Gasterosteus aculeatus). Aquat. Toxicol. 2019, 207, 142–152. [Google Scholar] [CrossRef]
- Kidd, K.A.; Blanchfield, P.J.; Mills, K.H.; Palace, V.P.; Evans, R.E.; Lazorchak, J.M.; Flick, R.W. Collapse of a fish population following exposure to a synthetic estrogen. Proc. Natl. Acad. Sci. USA 2007, 104, 8897–8901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clouzot, L.; Vanrolleghem, P.A. Endocrine Disruption: From a whole-lake experiment to a calibrated ecosystem model. Environ. Model. Softw. 2019, 115, 6–18. [Google Scholar] [CrossRef]
- Kidd, K.A.; Paterson, M.J.; Rennie, M.D.; Podemski, C.L.; Findlay, D.L.; Blanchfield, P.J.; Liber, K. Direct and indirect responses of a freshwater food web to a potent synthetic oestrogen. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudgeon, D. Prospects for sustaining freshwater biodiversity in the 21st century: Linking ecosystem structure and function. Curr. Opin. Environ. Sustain. 2010, 2, 422–430. [Google Scholar] [CrossRef]
- Teixeira, H.; Lillebø, A.I.; Culhane, F.; Robinson, L.; Trauner, D.; Borgwardt, F.; Kuemmerlen, M.; Barbosa, A.; McDonald, H.; Funk, A.; et al. Linking biodiversity to ecosystem services supply: Patterns across aquatic ecosystems. Sci. Total Environ. 2019, 657, 517–534. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Schielzeth, H.; Barnes, A.D.; Barry, K.E.; Bonn, A.; Brose, U.; Bruelheid, H.; Buchmann, N.; Buscot, F.; Ebeling, A.; et al. A multitrophic perspective on biodiversity–ecosystem functioning research. Adv. Ecol. Res. 2019, 61. [Google Scholar] [CrossRef]
- Hines, J.; Ebeling, A.; Barnes, A.D.; Brose, U.; Scherber, C.; Scheu, S.; Tscharntke, T.; Weisser, W.W.; Giling, D.P.; Klein, A.M.; et al. Mapping change in biodiversity and ecosystem function research: Food webs foster integration of experiments and science policy. Adv. Ecol. Res. 2019, 61. [Google Scholar] [CrossRef]
- Langhans, S.D.; Jähnig, S.C.; Lago, M.; Schmidt–Kloiber, A.; Hein, T. The potential of ecosystem–based management to integrate biodiversity conservation and ecosystem service provision in aquatic ecosystems. Sci. Total Environ. 2019, 672, 1017–1020. [Google Scholar] [CrossRef]
- Duffy, J.E.; Cardinale, B.J.; France, K.E.; McIntyre, P.B.; Thébault, E.; Loreau, M. The functional role of biodiversity in ecosystems: Incorporating trophic complexity. Ecol. Lett. 2007, 10, 522–538. [Google Scholar] [CrossRef] [Green Version]
- Daam, M.A.; Teixeira, H.; Lillebø, A.I.; Nogueira, A.J.A. Establishing causal links between aquatic biodiversity and ecosystem functioning: Status and research needs. Sci. Total Environ. 2019, 656, 1145–1156. [Google Scholar] [CrossRef]
- Queirós, A.M.; Bruggeman, J.; Stephens, N.; Artioli, Y.; Butenschön, M.; Blackford, J.C.; Widdicombe, S.; Allen, J.I.; Somerfield, P.J. Placing biodiversity in ecosystem models without getting lost in translation. J. Sea Res. 2015, 98, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Barnes, A.D.; Jochum, M.; Lefcheck, J.S.; Eisenhauer, N.; Scherber, C.; O’Connor, M.I.; de Ruiter, P.; Brose, U. Energy flux: The link between multitrophic biodiversity and ecosystem functioning. Trends Ecol. Evol. 2018, 33, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Vanhooren, H.; Meirlaen, J.; Amerlinck, Y.; Claeys, F.; Vangheluwe, H.; Vanrolleghem, P.A. WEST: Modelling biological wastewater treatment. J. Hydroinform. 2003, 5, 27–50. [Google Scholar] [CrossRef] [Green Version]
- Chair, S.N.; Chapin, F.S., III; Costanza, R.; Ehrlich, P.R.; Golley, F.B.; Hooper, D.U.; Lawton, J.H.; O.Neill, R.V.; Mooney, H.A.; Sala, O.A.; et al. Biodiversity and Ecosystem Functioning: Maintaining Natural Life Support Processes. Ecol.–ESA 1999, 4. [Google Scholar] [CrossRef]
- Vaneeckhaute, C.; Remigi, E.; Tack, F.M.G.; Meers, E.; Belia, E.; Vanrolleghem, P.A. Optimizing the configuration of integrated nutrient and energy recovery treatment trains: A new application of global sensitivity analysis to the generic nutrient recovery model (NRM) library. Bioresour. Technol. 2018, 269, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Hanazato, T. Effects of repeated application of carbaryl on zooplankton communities in experimental ponds with or without the predator Chaoborus. Environ. Pollut. 1991, 74, 309–324. [Google Scholar] [CrossRef]
- Tang, K.W.; Flury, S.; Grossart, H.P.; McGinnis, D.F. The Chaoborus pump: Migrating phantom midge larvae sustain hypolimnetic oxygen deficiency and nutrient internal loading in lakes. Water Res. 2017, 122, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Le Guen, C.; Tecchio, S.; Dauvin, J.C.; De Roton, G.; Lobry, J.; Lepagef, M.; Morin, J.; Lassalle, G.; Raoux, A.; Niquil, N. Assessing the ecological status of an estuarine ecosystem: Linking biodiversity and food–web indicators. Estuar. Coast. Shelf Sci. 2019, 228, 106339. [Google Scholar] [CrossRef]
- Jabiol, J.; McKie, B.G.; Bruder, A.; Bernadet, C.; Gessner, M.O.; Chauvet, E. Trophic complexity enhances ecosystem functioning in an aquatic detritus–based model system. J. Anim. Ecol. 2013, 82, 1042–1051. [Google Scholar] [CrossRef] [Green Version]
- Duncan, C.; Thompson, J.R.; Pettorelli, N. The quest for a mechanistic understanding of biodiversity–ecosystem services relationships. Proc. R. Soc. B 2015, 282, 20151348. [Google Scholar] [CrossRef] [Green Version]
- Bennett, E.M.; Cramer, W.; Begossi, A.; Cundill, G.; Dıaz, S.; Egoh, B.N.; Geijzendorffer, I.R.; Krug, C.B.; Lavorel, S.; Woodward, G.; et al. Linking biodiversity, ecosystem services, and human well–being: Three challenges for designing research for sustainability. Curr. Opin. Environ. Sustain. 2015, 14, 76–85. [Google Scholar] [CrossRef]




















© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clouzot, L.; Haguenauer, C.; Vanrolleghem, P.A. An Extended Ecosystem Model for Understanding EE2 Indirect Effects on a Freshwater Food Web and its Ecosystem Function Resilience. Water 2020, 12, 1736. https://doi.org/10.3390/w12061736
Clouzot L, Haguenauer C, Vanrolleghem PA. An Extended Ecosystem Model for Understanding EE2 Indirect Effects on a Freshwater Food Web and its Ecosystem Function Resilience. Water. 2020; 12(6):1736. https://doi.org/10.3390/w12061736
Chicago/Turabian StyleClouzot, Ludiwine, Charlotte Haguenauer, and Peter A. Vanrolleghem. 2020. "An Extended Ecosystem Model for Understanding EE2 Indirect Effects on a Freshwater Food Web and its Ecosystem Function Resilience" Water 12, no. 6: 1736. https://doi.org/10.3390/w12061736
APA StyleClouzot, L., Haguenauer, C., & Vanrolleghem, P. A. (2020). An Extended Ecosystem Model for Understanding EE2 Indirect Effects on a Freshwater Food Web and its Ecosystem Function Resilience. Water, 12(6), 1736. https://doi.org/10.3390/w12061736