Modelling PAHs Transfer from Polluted Soil to Herbaceous Species in Phytoremediation Attempts

Abstract
:
1. Introduction
2. Materials and Methods
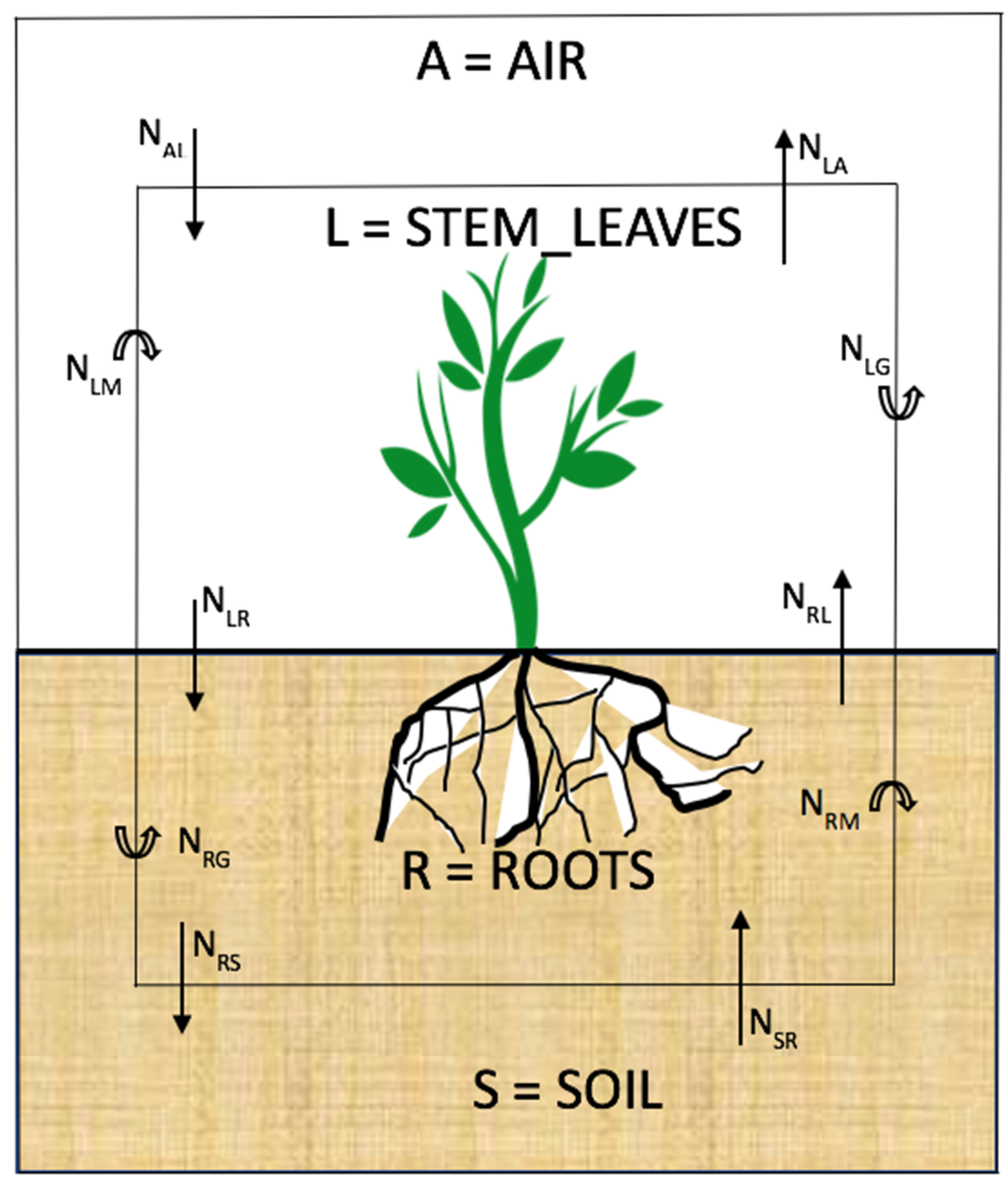
2.1. Conceptual Model
2.2. Model Equations
- -
- Mi = mass of PAH in compartment i;
- -
- Zi = fugacity capacity of compartment i;
- -
- fi and fj = fugacity of PAH in compartments i and j (Pa);
- -
- Vi = volume of compartment i (m3);
- -
- Dji = transport D values to compartment i from compartment j with fugacity fj, including the flow in xylem and phloem and the uptake from soil and air (mol·Pa−1·h−1);
- -
- Do = transport and transformation D values for processes by which pollutant is removed from a giving compartment (mol·Pa−1·h−1).
2.2.1. Fugacity Expression of the Plant Compartments
- -
- A = Air, R = Roots, S = Soil and L = Stems and Leaves
- -
- fA, fR, fS and fL are the fugacity of PAH in compartment A, R, S and L respectively;
- -
- DAL, DRL, DSR and DLR are the air to stem_leaves, roots to stem_leaves, soil to roots and stem_leaves to roots transport D values of PAH. With , with the PAH half-life.
- -
- DLM, DLG, are the PAH lost by metabolism and growth dilution. DLA, DLR,DRS and DRL are the other losses related to the first compartment. For example, DLA is the transport D value lost from stem_leaves to air (through transpiration for instance).
- -
- τAL,L, τRL,L, τLA,L and τLR,L are the air to stem_leaves, root to stem_leaves, stem_leaves to air and stem_leaves to root PAH clearance half-lives in the stem_leaves compartment;
- -
- τLR,R, τSR,R, τRL,R and τRS,R are the stem_leaves to root, soil to root, root to stem_leaves and root to soil PAH clearance half-lives in the root compartment;
- -
- kLG and kLM are stem_leaves growth dilution and metabolism rates constants, respectively; kRG and kRM the roots growth dilution and metabolism rates constants, respectively.
2.2.2. PAHs Concentrations in the Roots and Stem_Leaves Compartments
2.3. Model Input Parameters
- -
- Partition coefficients of PAHs between compartments;
- -
- Flowrates that control the transport processes;
- -
- Retention times of PAHs in each compartment of the plant according to the flow direction;
- -
- Half-lives of transport (τ), growth dilution (τG) and metabolism (τM).
- -
- Metabolism and growth dilution half-lives
- -
- Plant morphological and physiological parameters such as volumes of stem_leaves and roots compartments for each plant species; and the initial soil PAHs concentrations.
2.3.1. Calculated Input Parameters
Partition Coefficients and the Initial PAHs Concentration in Water
- -
- VC, VW, Vl, VF and VP are the volume fractions of cuticular membrane, water, carbohydrate, lipids and protein of the leaves.
- -
- KAW is the air to water partition coefficient and RT/H derived from the Henry’s law constant.
Flowrates and Retention Times
Transport Half-Lives
2.3.2. Existing Input Parameters
Metabolism and Growth Dilution Half-Lives
Plant Morphological and Physiological Parameters and Others
2.4. Overview of the Experimental Work
2.5. Model Resolution and Data Analysis
3. Results and Discussion
3.1. Sensitivity Analysis
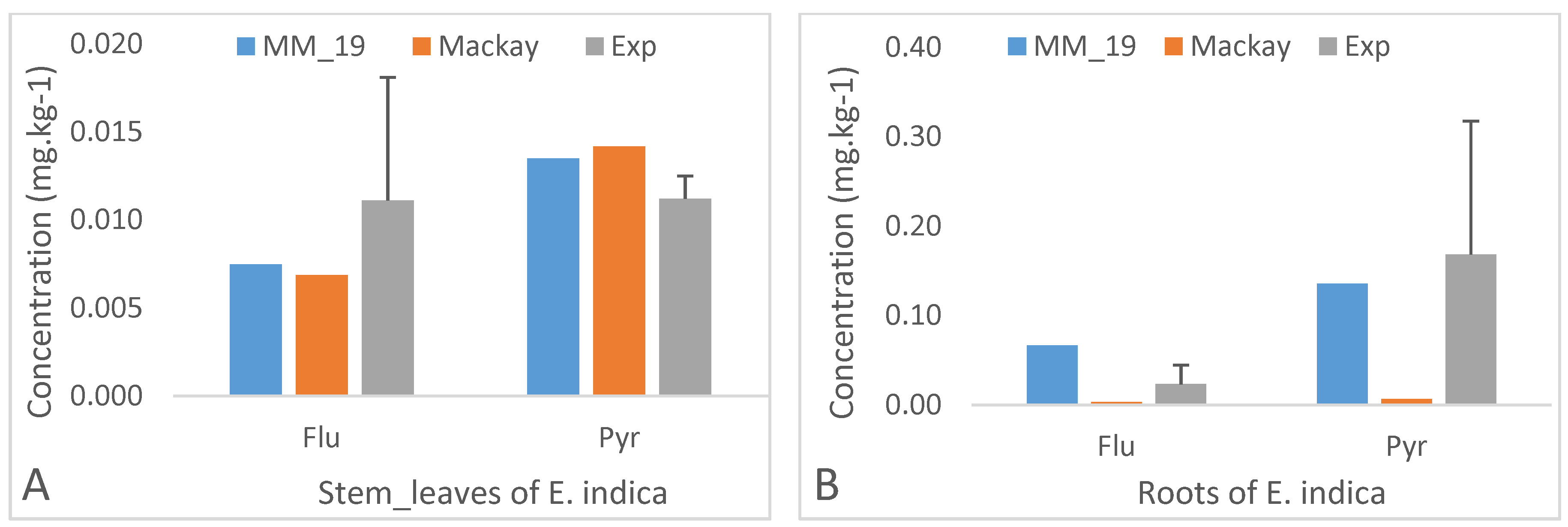
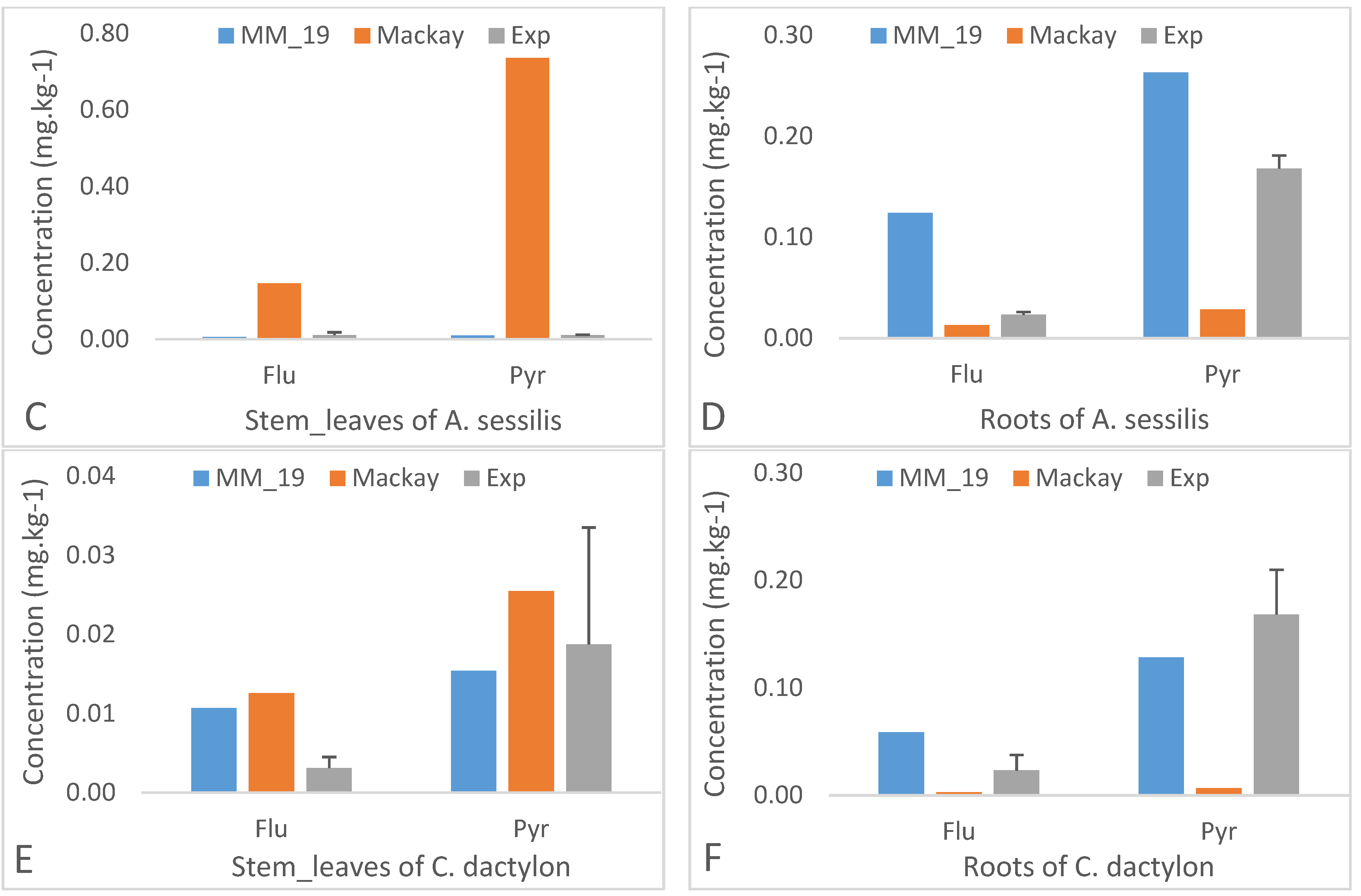
3.2. Impact of Light PAHs on Model Calibration
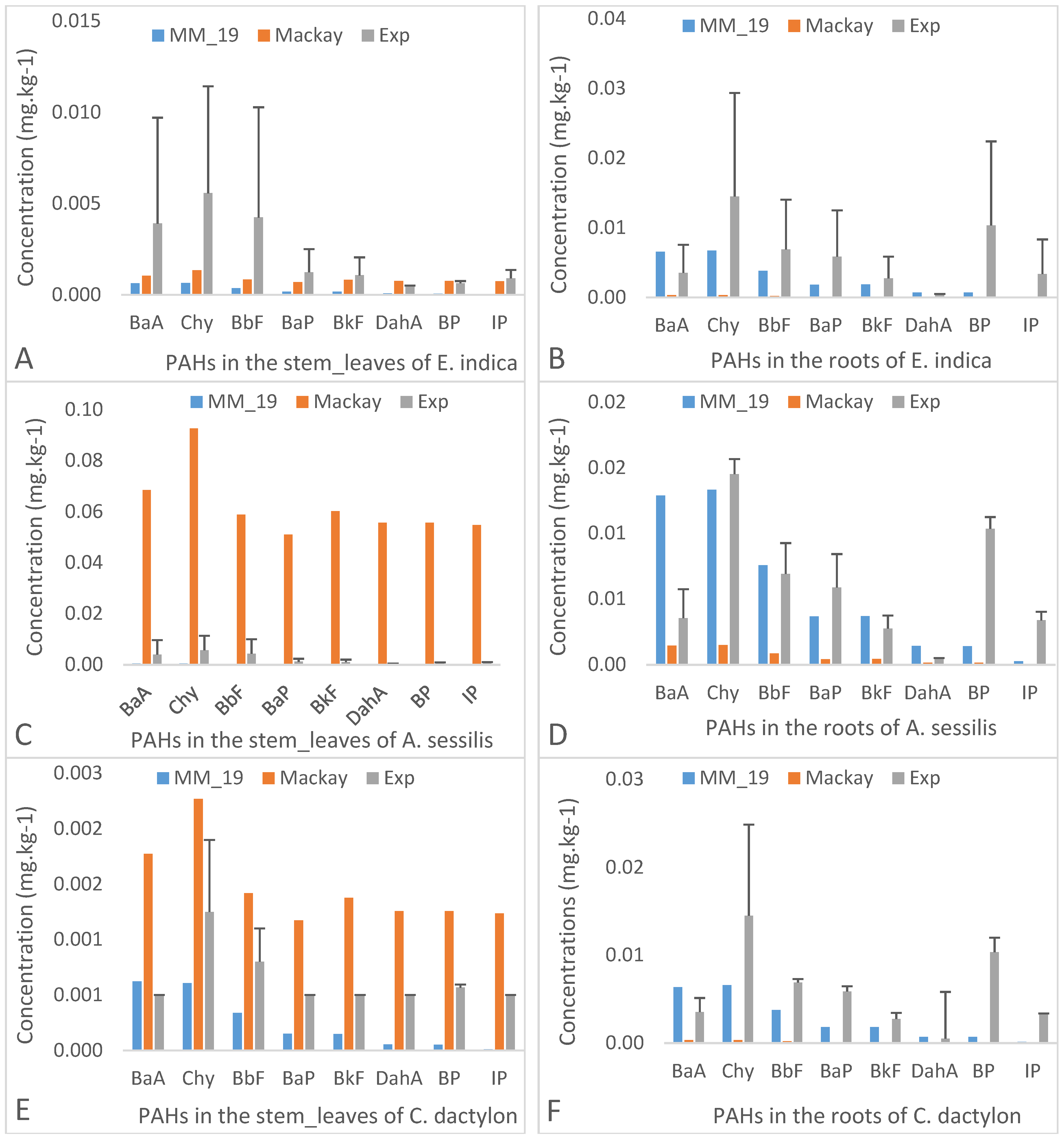
3.3. Impact of the Physico-Chemical Properties of PAHs on Their Transfer from Soil to the Plant Species
3.3.1. Intermediate Molecular Weights’ PAHs
3.3.2. High Molecular Weights’ PAHs
3.4. Comparison Between the MM_19 Model and the Mackay_97 Model
3.4.1. Significance of the Phloem Flow Multiplying Factor (α)
3.4.2. Models Efficiency
3.5. Modelling Results
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Collins, C.; Fryer, M.; Grosso, A. Plant uptake of non ionic organic chemicals. Environ. Sci. Technol. 2006, 40, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Hung, H.; Mackay, D. A novel and simple model of the uptake of organic chemicals by vegetation from air and soil. Chemosphere 1997, 35, 959–977. [Google Scholar] [CrossRef]
- Trapp, S.; Legind, C. Uptake of Organic Contaminants from Soil into Vegetables and Fruits, springerprofessional.de. 2011. Available online: https://www.springerprofessional.de/uptake-of-organic-contaminants-from-soil-into-vegetables-and-fru/4719244 (accessed on 22 May 2018).
- INERIS. Modèle de transfert sol-plante des polluants organiques. Tome 1: Revue bibliographique | Ineris. 2002. Available online: https://www.ineris.fr/fr/modele-transfert-sol-plante-polluants-organiques-tome-1-revue-bibliographique (accessed on 16 November 2018).
- GBriggs, G.; Bromilow, R.H.; Evans, A.A. Relationships between lipophilicity and root uptake and translocation of non-ionised chemicals by barley. Pestic. Sci. 1982, 13, 495–504. [Google Scholar] [CrossRef]
- Calamari, D.; Vighi, M.; Bacci, E.E. The use of terrestrial plant biomass as a parameter in the fugacity model. Chemosphere 1978, 16, 2359–2364. [Google Scholar] [CrossRef]
- Collins, C.D.; Finnegan, E. Modeling the Plant Uptake of Organic Chemicals, Including the Soil−Air−Plant Pathway. Environ. Sci. Technol. 2010, 44, 998–1003. [Google Scholar] [CrossRef] [PubMed]
- Empereur-Bissonnet, P. Modeling polycyclic aromatic hydrocarbon uptake in common vegetables grown in aged polluted soils. Environ. Risques St. 2013, 12, 29–40. [Google Scholar] [CrossRef]
- Nguemté, P.M.; Wafo, G.V.D.; Djocgoue, P.F.; Noumsi, I.M.K.; Ngnien, A.W. Potentialities of Six Plant Species on Phytoremediation Attempts of Fuel Oil-Contaminated Soils. Water Air Soil Pollut. 2018, 229, 88. [Google Scholar] [CrossRef]
- Danesh, A. (Ed.) 3-Phase Equilibria. In Developments in Petroleum Science; Elsevier: Amsterdam, The Netherlands, 1998; pp. 105–128. [Google Scholar]
- Karickhoff, S.W. Semi-empirical estimation of sorption of hydrophobic pollutants on natural sediments and soils. Chemosphere 1981, 10, 833–846. [Google Scholar] [CrossRef]
- Muller, J.F.; Hawker, D.W.; Connell, D.W. Calculation of bioconcentration factors of persistent hydrophobic compounds in the air/vegetation system. Chemosphere 1994, 29, 623–640. [Google Scholar] [CrossRef]
- Nguemté, P.; Wafo, G.V.; Djocgoue, P.; Noumsi, I.K.; Ngnien, A.W. Phytoremédiation de sols pollués par les hydrocarbures–évaluation des potentialités de six espèces végétales tropicales. Rev. Sci. L8217eau J. Water Sci. 2017, 30, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Lasdon, L.S.; Fox, R.L.; Ratner, M.W. Nonlinear optimization using the generalized reduced gradient method. RAIRO-Oper. Res.-Rech. Opérationnelle 1974, 8, 73–103. [Google Scholar] [CrossRef] [Green Version]
- Oleszczuk, P.; Baran, S. Polycyclic Aromatic Hydrocarbons Content in Shoots and Leaves of Willow (Salix). Water Air Soil Pollut. 2005, 168, 91–111. [Google Scholar] [CrossRef]
- Hajabbasi, M.A. Importance of soil physical characteristics for petroleum hydrocarbons phytoremediation: A review. Afr. J. Environ. Sci. Technol. 2016, 10, 394–405. [Google Scholar] [CrossRef] [Green Version]
- Binet, P.; Portal, J.M.; Leyval, C. Dissipation of 3–6-ring polycyclic aromatic hydrocarbons in the rhizosphere of ryegrass. Soil Biol. Biochem. 2000, 32, 2011–2017. [Google Scholar] [CrossRef]
- Rodrigues, R.; Lacerda, C.A.; Lanças, F.M. Coating of solid-phase microextraction fibers with chemically bonded silica particles: Selective extraction of polycyclic aromatic hydrocarbons from drinking water samples. J. Chromatogr. Sci. 2002, 40, 489–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramaniyam, A. The Influence of Plants in the Remediation of Petroleum Hydrocarbon- Contaminated Sites. Pharm. Anal. Chem. Open Access 2015, 1, 1–11. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
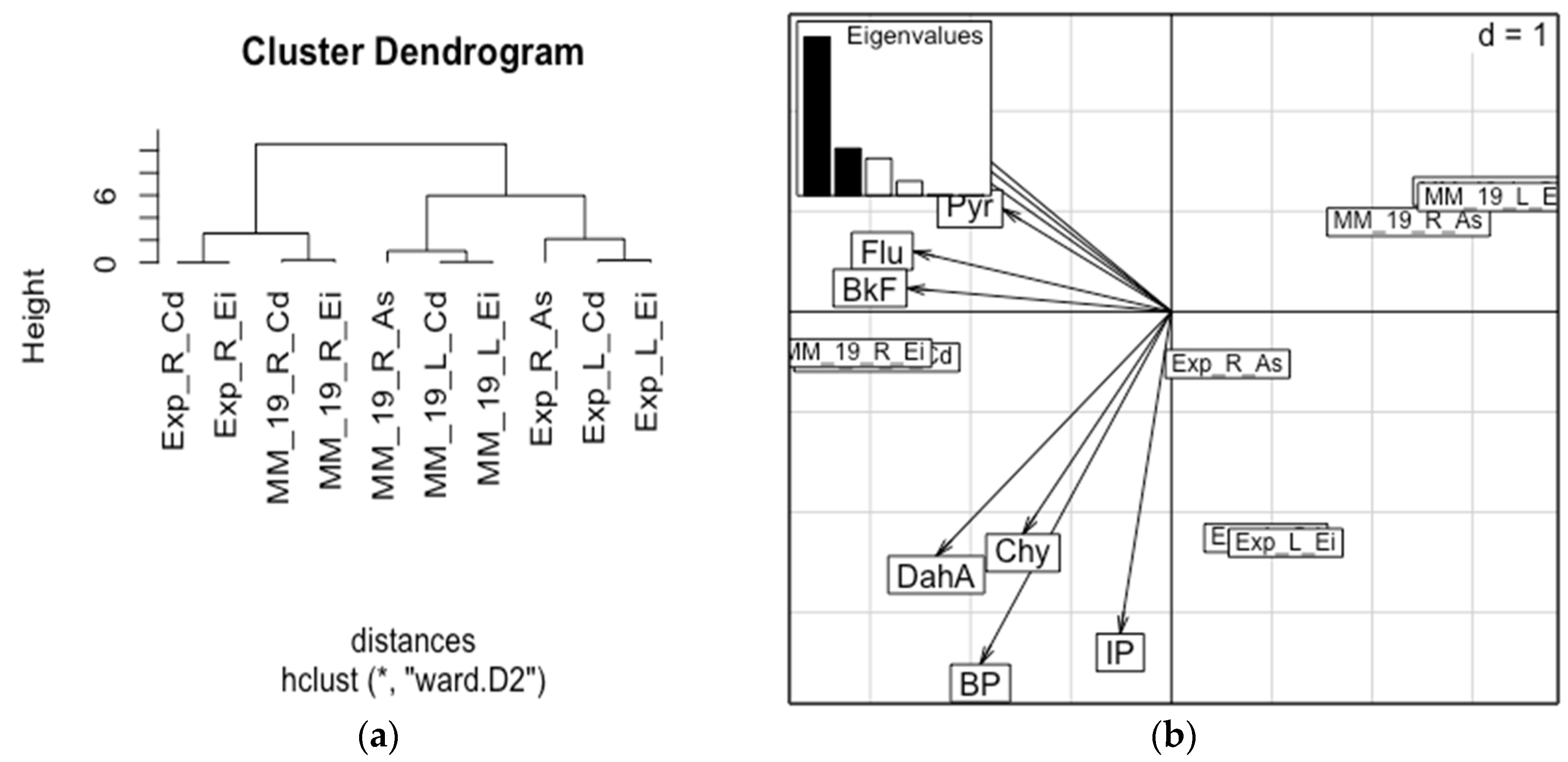{kind=link}
| Models | Phloem Flow Multiplying Factor (α) | E. indica | C. dactylon | A. sessilis |
|---|---|---|---|---|
| MM_19 | Initial value | 5.00 × 10−2 | 6.64 × 10−5 | 6.64 × 10−5 |
| Calibrated value | 6.64 × 10−5 | 6.95 × 10−5 | 9.22 × 10−5 | |
| Mackay_97 | Initial value | 5.00 × 10−2 | 2.72 × 10−4 | 2.72 × 10−4 |
| Calibrated value | 2.72 × 10−4 | 6.07 × 10−4 | 4.69 × 10−2 |
| NSE | Models | E. indica | C. dactylon | A. sessilis |
|---|---|---|---|---|
| NSE_L | MM_19 | 5.50 × 10−1 | 8.07 × 10−1 | 8.45 × 10−1 |
| Mackay_97 | 5.87 × 10−1 | 5.20 × 10−1 | −6.56 × 103 | |
| NSE_R | MM_19 | 8.60 × 10−1 | 7.24 × 10−1 | −1.05 × 102 |
| Mackay_97 | −1.47 × 10−1 | −1.95 × 10−1 | 7.87 × 10−1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsodoum Nguemté, P.; Kengne Noumsi, I.M.; Djumyom Wafo, G.V.; Djocgoue, P.F.; Wanko, A. Modelling PAHs Transfer from Polluted Soil to Herbaceous Species in Phytoremediation Attempts. Water 2020, 12, 1759. https://doi.org/10.3390/w12061759
Matsodoum Nguemté P, Kengne Noumsi IM, Djumyom Wafo GV, Djocgoue PF, Wanko A. Modelling PAHs Transfer from Polluted Soil to Herbaceous Species in Phytoremediation Attempts. Water. 2020; 12(6):1759. https://doi.org/10.3390/w12061759
Chicago/Turabian StyleMatsodoum Nguemté, Pulchérie, Ives Magloire Kengne Noumsi, Guy Valérie Djumyom Wafo, Pierre François Djocgoue, and Adrien Wanko. 2020. "Modelling PAHs Transfer from Polluted Soil to Herbaceous Species in Phytoremediation Attempts" Water 12, no. 6: 1759. https://doi.org/10.3390/w12061759
APA StyleMatsodoum Nguemté, P., Kengne Noumsi, I. M., Djumyom Wafo, G. V., Djocgoue, P. F., & Wanko, A. (2020). Modelling PAHs Transfer from Polluted Soil to Herbaceous Species in Phytoremediation Attempts. Water, 12(6), 1759. https://doi.org/10.3390/w12061759