Role of the Hyporheic Zone in Increasing the Resilience of Mountain Streams Facing Intermittency

 ,
, 

Abstract
:1. Introduction
- (1)
- different faunistic groups respond differently to intermittence, due to their different degree of specialization to life in the hyporheic; in particular, stygobites would be less affected by drought than stygophile and stygoxenes;
- (2)
- the connection with surface water and the direction of the vertical aquifer flow determine the faunistic composition at different depths.
- (3)
- the hyporheic acts as a refuge increasing the resilience of the benthic communities to intermittence, but resilience decreases with increasing degree of specialization to life in the hyporheic.
2. Materials and Methods
2.1. Study Area
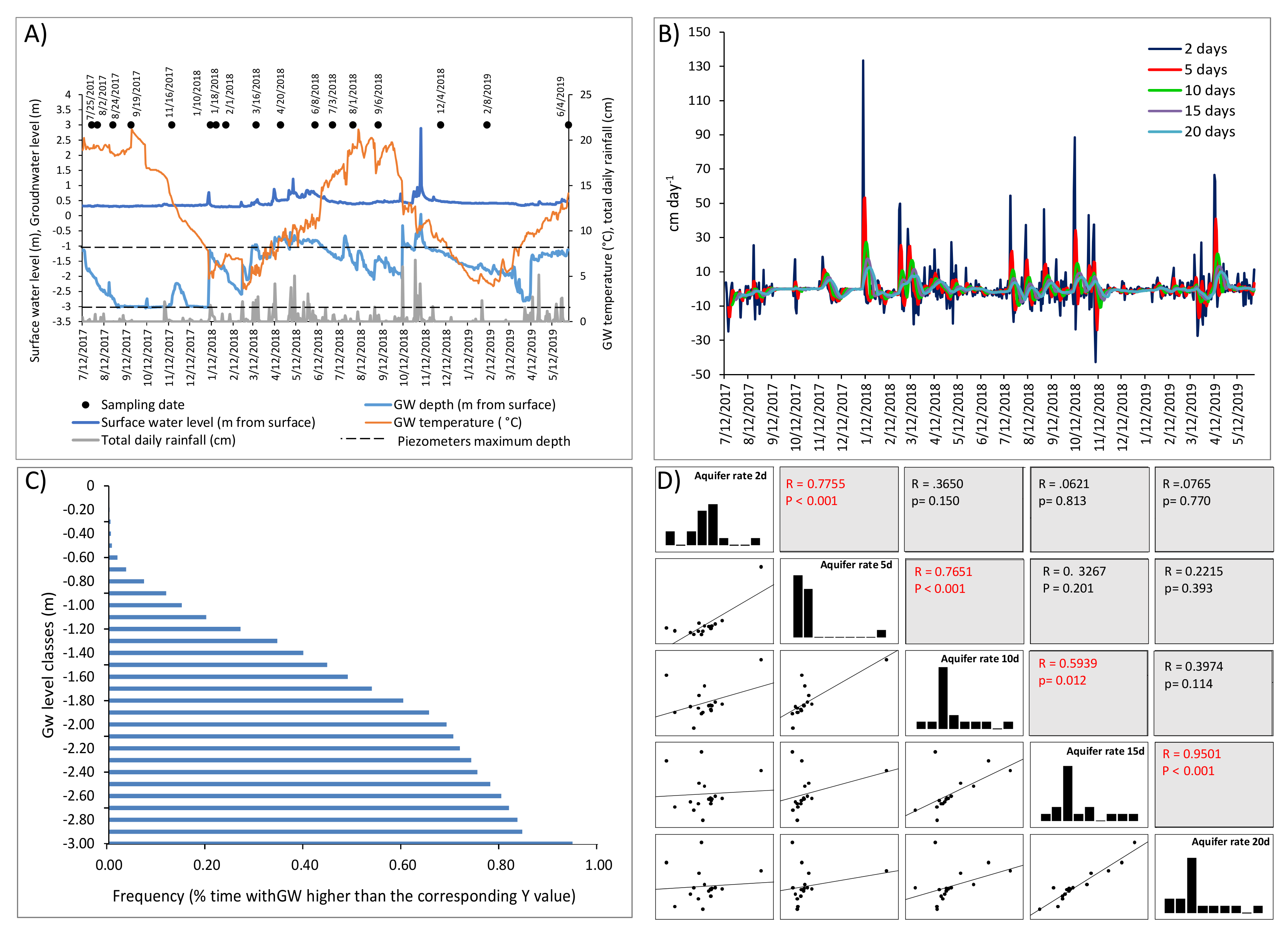
2.2. Hydrological and Physical-Chemical Data
2.3. Invertebrate Sampling
2.4. Data Analysis
- (1)
- Factor “station”: perm = permanent station; int-1 = intermittent station reaching −1 m depth; int-3 = intermittent station reaching −3 m depth;
- (2)
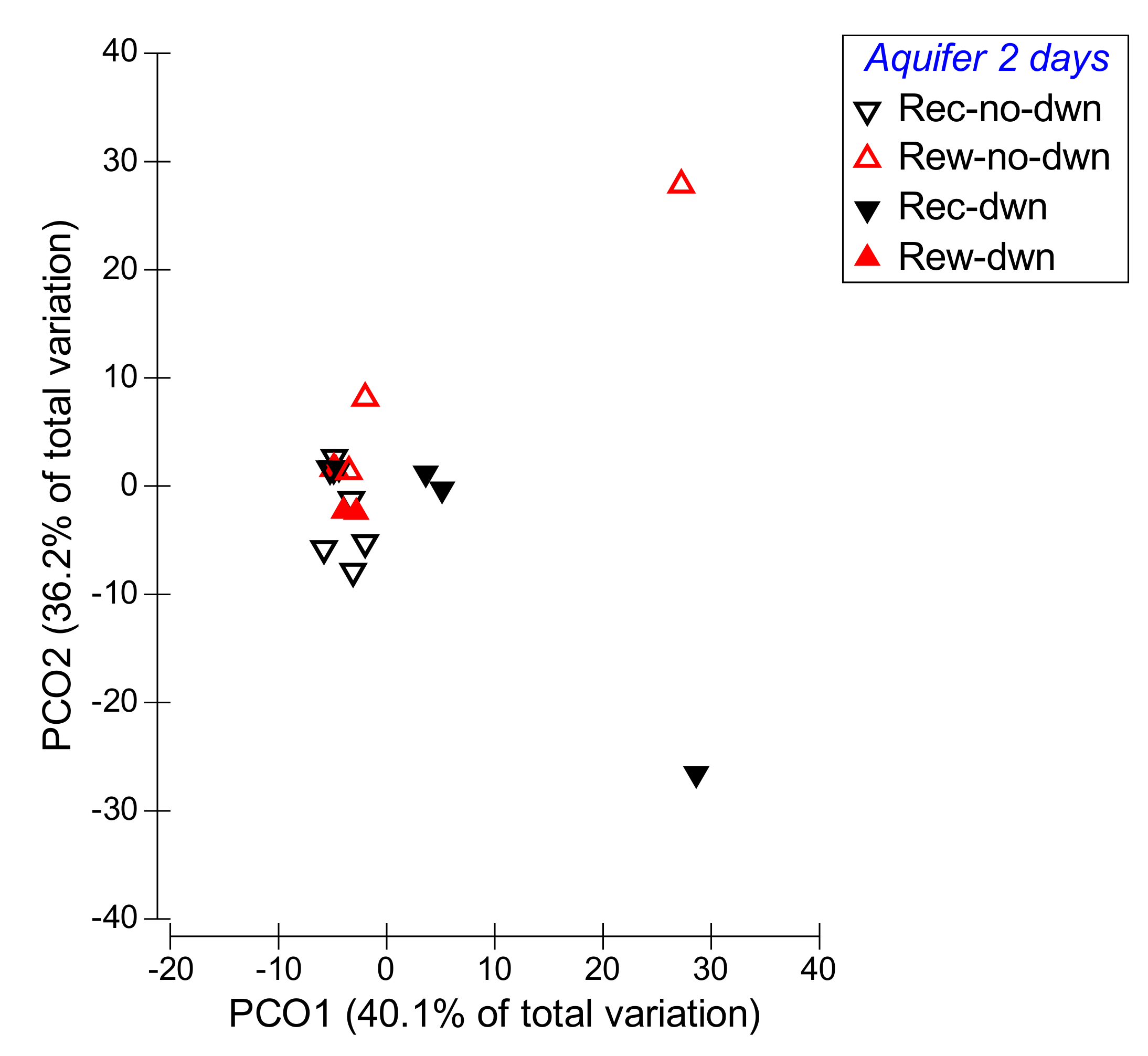
- Factor “aquifer phase” (for the two intermittent stations only): recession without downwelling, recession with downwelling, rewetting with downwelling, rewetting without downwelling. These phases were based on the aquifer recession rate calculated over relevant time intervals preceding the sampling date (see results: hydrology, for explanation of the time-interval selection), and the presence/absence of surface water. In detail: downwelling flow occurred when surface water was present and recharged the aquifer, and no downwelling when surface water was not present; recession when the water table level was lowering (negative recession rate); rewetting when the water table level was rising (positive recession rate).
- (3)
- Factor “hydrological phase” based on the condition of surface water at the site: drought (i.e., no surface water), low flow (i.e., surface water level below the threshold of 15 cm at perm and int-1, and 10 cm at int-3), high flow (i.e., surface water level above the 15/10 cm thresholds).
2.4.1. Community Metrics
2.4.2. Hydrology/Community Metrics Relationships
2.4.3. Species Traits
3. Results
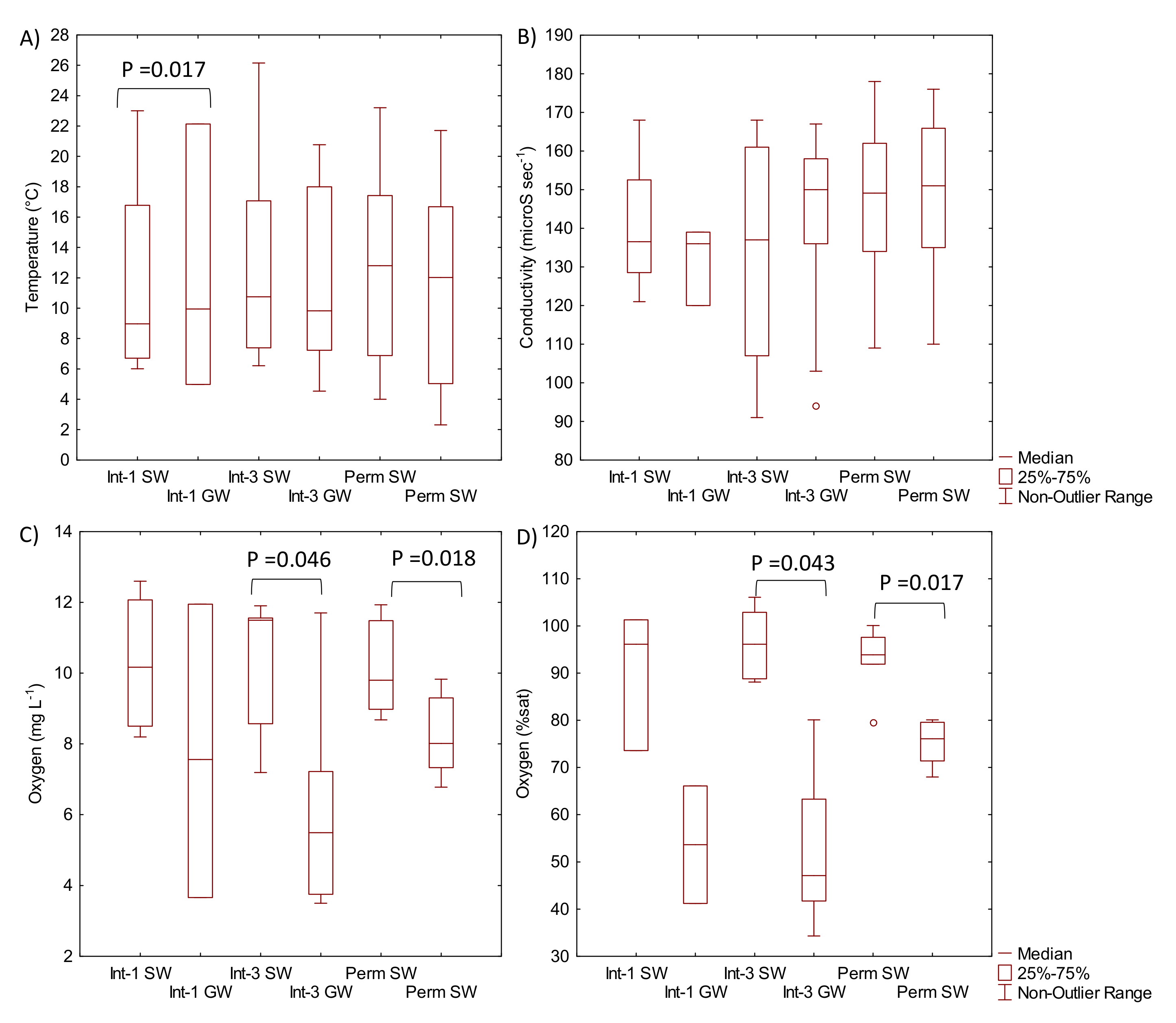
3.1. Hydrology and Physical-Chemical Variables
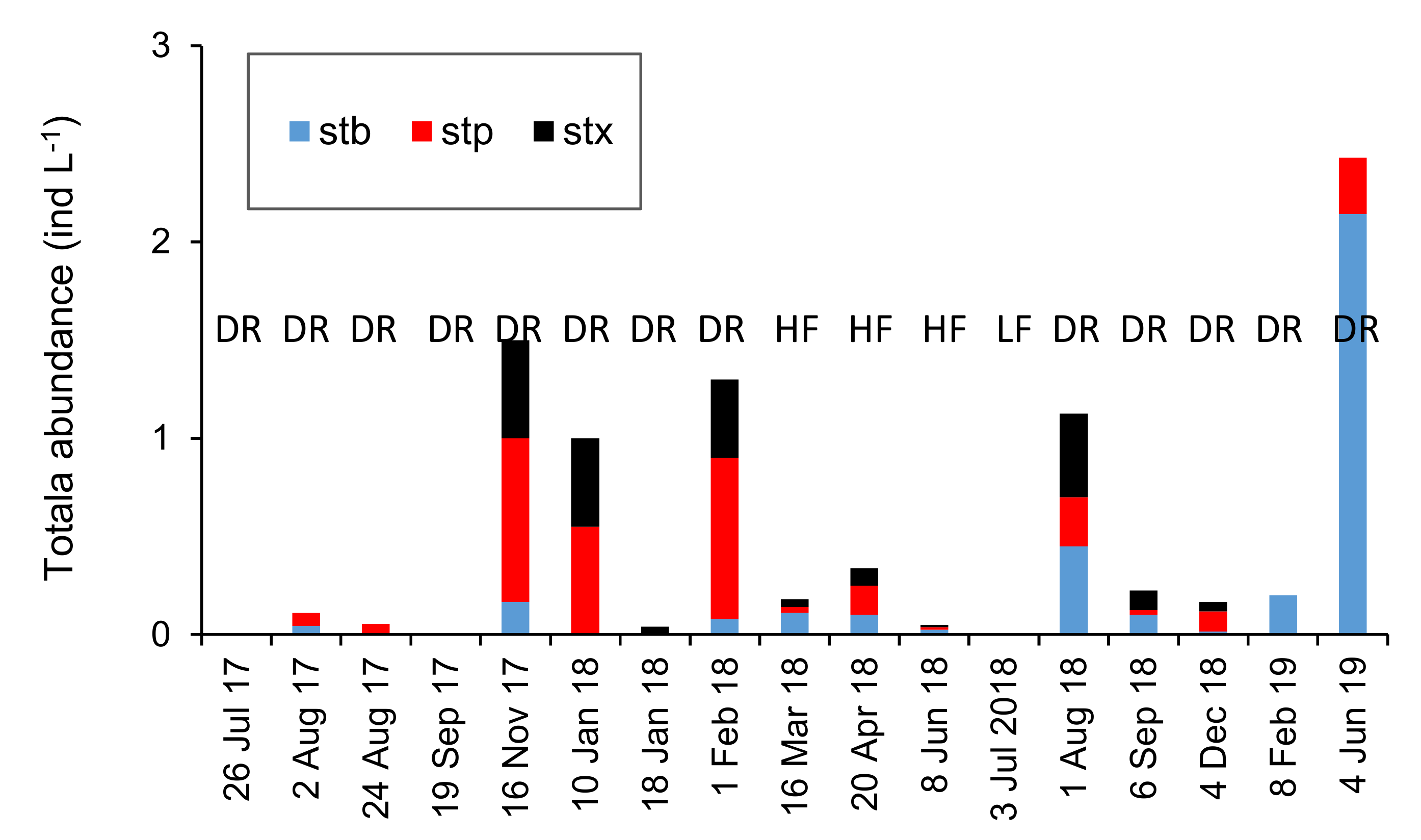
3.2. Hp1: Responses to Intermittence of the Three Faunistic Groups Differ: Community Metrics.
3.2.1. Community Metrics: Effect of Intermittence in The Shallow Hyporheic (−1 m)
3.2.2. Community Metrics: Effect of Intermittence Over Depth
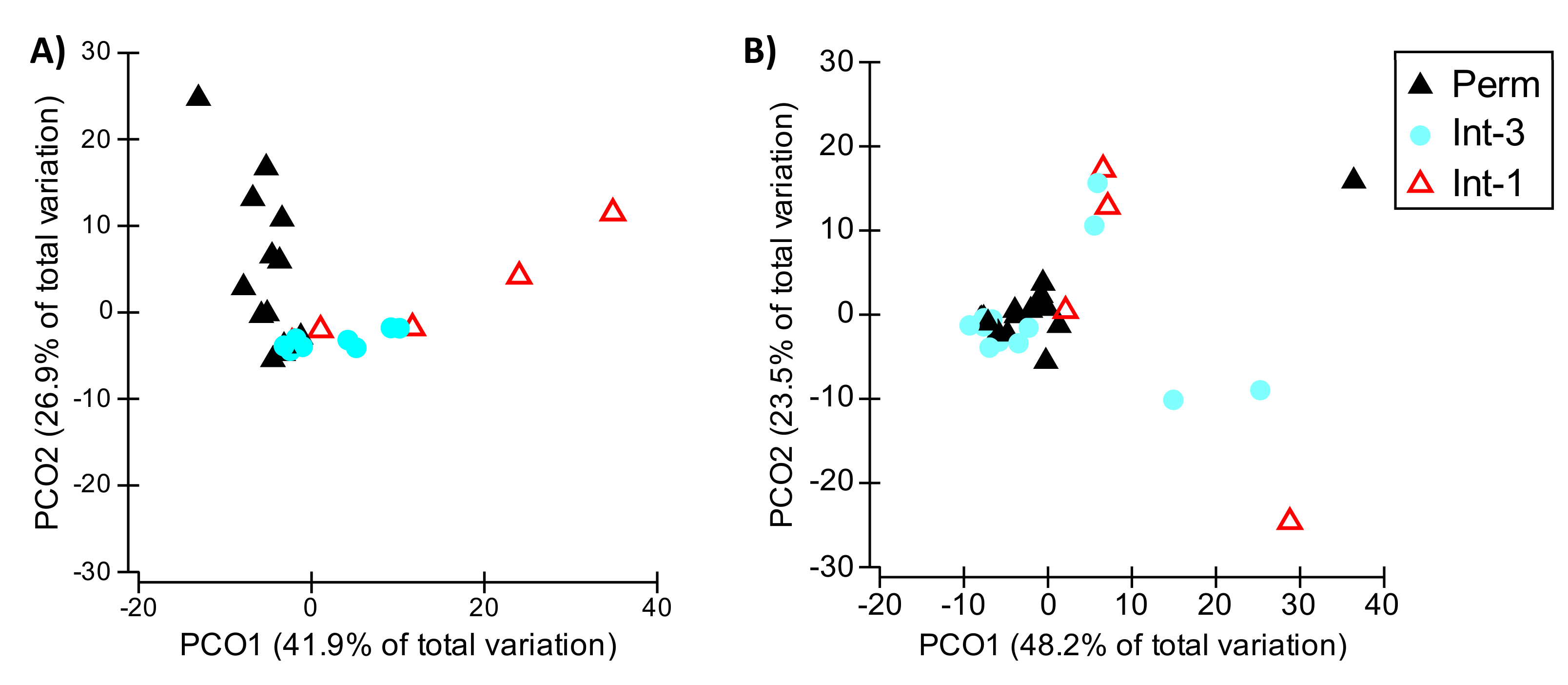
3.3. Hp 2: Hydrological Connectivity and Aquifer Flow Dynamics Drive Faunal Patterns: Hydrology/Community Metrics Relationships
3.4. Hp 3: the Hyporheic Use Increases the Resilience of the System, with Faunistic-Specific Resilience Responses
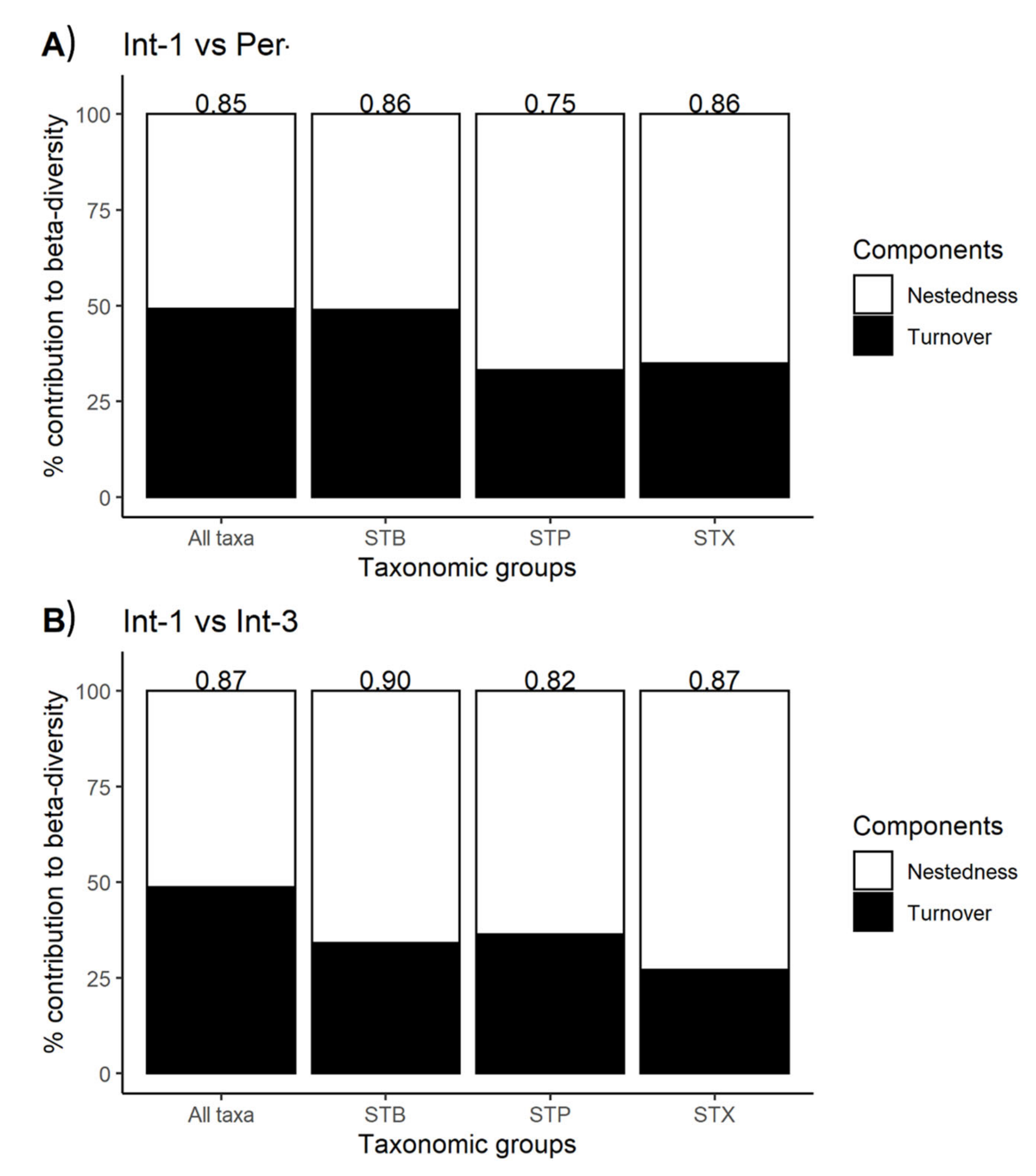
3.4.1. Analysis of Nestedness and Turnover
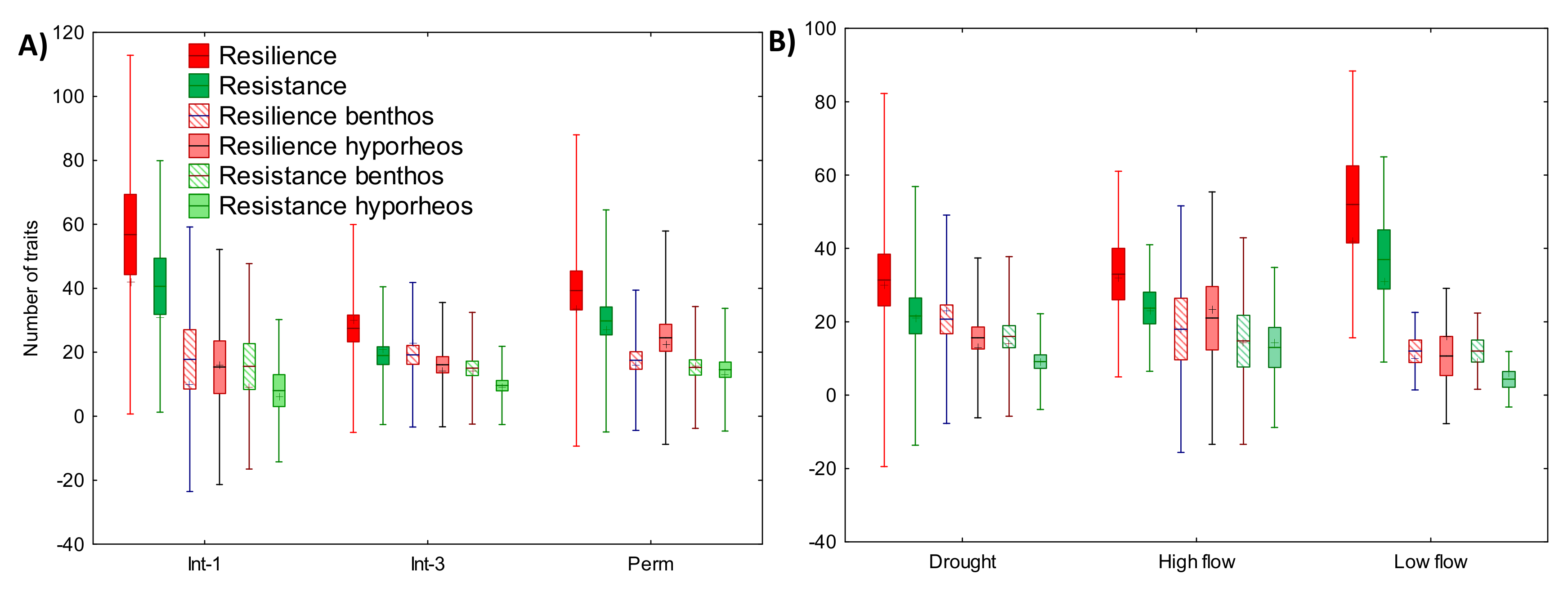
3.4.2. Species Traits
4. Discussion
4.1. Hp1: Responses to Intermittence of the Three Faunistic Groups Differ
4.2. Hp 2: Hydrological Connectivity and Aquifer Flow Dynamics Drive Faunal Patterns
4.3. Hp 3: the Hyporheic Use Increases the Resilience of the System, with Faunistic-Specific Resilience Responses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jiménez Cisneros, B.E.; Oki, T.; Arnell, N.W.; Benito, G.; Cogley, J.G.; Döll, P.; Jiang, T.; Mwakalila, S.S. Freshwater resources. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 229–269. [Google Scholar]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segan, D.B.; Murray, K.A.; Watson, J.E.M. A global assessment of current and future biodiversity vulnerability to habitat loss—climate change interactions. Glob. Ecol. Conserv. 2016, 5, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Beniston, M. Impacts of climatic change on water and associated economic activities in the Swiss Alps. J. Hydrol. 2012, 412, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Gobiet, A.; Kotlarski, S.; Beniston, M.; Heinrich, G.; Rajczak, J.; Stoffel, M. 21st century climate change in the European Alps—A review. Sci. Total Environ. 2014, 493, 1138–1151. [Google Scholar] [CrossRef]
- Fenoglio, S.; Bo, T.; Cucco, M.; Mercalli, L.; Malacarne, G. Effects of global climate change on freshwater biota: A review with special emphasis on the Italian situation. Ital. J. Zool. 2010, 77, 374–383. [Google Scholar] [CrossRef]
- Wilhite, D.A. Drought as a natural hazard: Concepts and definitions. In Drought: A Global Assessment; Wilhite, D.A., Ed.; Routledge: London, UK, 2000; Volume I, pp. 3–18. [Google Scholar]
- Allan, J.D.; Palmer, M.; Poff, N.L. Climate change and freshwater ecosystems. In Climate Change and Biodiversity; Lovejoy, T.E., Hannah, L., Eds.; Yale University Press: New Haven, CT, USA; London, UK, 2005; pp. 274–290. [Google Scholar]
- Durance, I.; Ormerod, S.J. Climate change effects on upland stream macroinvertebrates over a 25-year period. Glob. Chang. Biol. 2007, 13, 942–957. [Google Scholar] [CrossRef]
- Ledger, M.E.; Brown, L.E.; Edwards, F.K.; Milner, A.M.; Woodward, G. Drought alters the structure and functioning of complex food webs. Nat. Clim. Chang. 2013, 3, 223–227. [Google Scholar] [CrossRef]
- Pinna, M.; Marini, G.; Cristiano, G.; Mazzotta, L.; Vignini, P.; Cicolani, B.; Di Sabatino, A. Influence of aperiodic summer droughts on leaf litter breakdown and macroinvertebrate assemblages: Testing the drying memory in a Central Apennines River (Aterno River, Italy). Hydrobiologia 2016, 782, 111–126. [Google Scholar] [CrossRef]
- Storey, R. Macroinvertebrate community responses to duration, intensity and timing of annual dry events in intermittent forested and pasture streams. Aquat. Sci. 2016, 78, 395–414. [Google Scholar] [CrossRef]
- Leigh, C.; Bonada, N.; Boulton, A.J.; Hugueny, B.; Larned, S.T.; Vander Vorste, R.; Datry, T. Invertebrate assemblage responses and the dual roles of resistance and resilience to drying in intermittent rivers. Aquat. Sci. 2016, 78, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Aspin, T.W.H.; Khamis, K.; Matthews, T.J.; Milner, A.M.; O’Callaghan, M.J.; Trimmer, M.G.; Woodward, G.; Ledger, M.E. Extreme drought pushes stream invertebrate communities over functional thresholds. Glob. Chang. Biol. 2019, 25, 230–244. [Google Scholar] [CrossRef] [PubMed]
- Lepori, F.; Hjerdt, N. Disturbance and aquatic biodiversity: Reconciling contrasting views. BioScience 2006, 56, 809–818. [Google Scholar] [CrossRef]
- Bonada, N.; Dolédec, S.; Statzner, B. Taxonomic and biological trait differences of stream macroinvertebrate communities between mediterranean and temperate regions: Implications for future climatic scenarios. Glob. Chang. Biol. 2007, 13, 1658–1671. [Google Scholar] [CrossRef]
- Verberk, W.C.E.P.; van Noordwijk, C.G.E.; Hildrew, A.G. Delivering on a promise: Integrating species traits to transform descriptive community ecology into a predictive science. Freshw. Sci. 2013, 32, 531–547. [Google Scholar] [CrossRef] [Green Version]
- Lake, P.S. Resistance, Resilience and Restoration. Ecol. Manag. Restor. 2013, 14, 20–24. [Google Scholar] [CrossRef]
- Lake, P.S. Ecological effects of perturbation by drought in flowing waters. Freshw. Biol. 2003, 48, 1161–1172. [Google Scholar] [CrossRef]
- Doretto, A.; Piano, E.; Falasco, E.; Fenoglio, S.; Bruno, M.C.; Bona, F. Investigating the role of refuges and drift on the resilience of macroinvertebrate communities to drying conditions: An experiment in artificial streams. River Res. Appl. 2018, 34, 777–785. [Google Scholar] [CrossRef]
- Fenoglio, S.; Bo, T.; Cucco, M.; Malacarne, G. Response of benthic invertebrate assemblages to varying drought conditions in the Po river (NW Italy). Ital. J. Zool. 2007, 74, 191–201. [Google Scholar] [CrossRef]
- Tierno de Figueroa, J.M.; López-Rodríguez, M.J.; Fenoglio, S.; Sánchez-Castillo, P.; Fochetti, R. Freshwater biodiversity in the rivers of the Mediterranean Basin. Hydrobiologia 2013, 719, 137–186. [Google Scholar] [CrossRef]
- Robson, B.J.; Chester, E.T.; Austin, C.M. Why life history information matters: Drought refuges and macroinvertebrate persistence in non-perennial streams subject to a drier climate. Mar. Freshw. Res. 2011, 62, 801–810. [Google Scholar] [CrossRef]
- Datry, T.; Larned, S.T.; Fritz, K.M.; Bogan, M.T.; Wood, P.J.; Meyer, E.I.; Santos, A.N. Broad-scale patterns of invertebrate richness and community composition in temporary rivers: Effects of flow intermittence. Ecography 2014, 37, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, J.; Belyea, L.R. Nested hierarchies and scale dependence of mechanisms of flow refugium use. J. N. Am. Benthol. Soc. 1997, 16, 221–238. [Google Scholar] [CrossRef]
- Boulton, A.J.; Lake, P.S. Effects of drought on stream insects and its ecological consequences. In Aquatic Insects: Challenges to Populations; Lancaster, J., Briers, R.A., Eds.; CAB International: Wallingford, UK, 2008; pp. 81–102. [Google Scholar]
- Stubbington, R.; Wood, P.J.; Boulton, A.J. Low flow controls on benthic and hyporheic macroinvertebrate assemblages during supra-seasonal drought. Hydrol. Proc. 2009, 23, 2252–2263. [Google Scholar] [CrossRef] [Green Version]
- Stubbington, R. The hyporheic zone as an invertebrate refuge: A review of variability in space, time, taxa and behavior. Mar. Freshw. Res. 2012, 63, 293–311. [Google Scholar] [CrossRef] [Green Version]
- Datry, T.; Pella, H.; Leigh, C.; Bonada, N.; Hugueny, B. A landscape approach to advance intermittent river ecology. Freshw. Biol. 2016, 61, 1200–1213. [Google Scholar] [CrossRef] [Green Version]
- Dole-Olivier, M.-J. The hyporheic refuge hypothesis reconsidered: A review of hydrological aspects. Mar. Freshw. Res. 2011, 62, 1281–1302. [Google Scholar] [CrossRef]
- Boulton, A.J.; Findlay, S.; Marmonier, P.; Stanley, E.H.; Valett, H.M. The functional significance of the hyporheic zone in streams and rivers. Annu. Review Ecol. Syst. 1998, 29, 59–81. [Google Scholar] [CrossRef] [Green Version]
- Delucchi, C.M. Movement patterns of invertebrates in temporary and permanent streams. Oecologia 1989, 78, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Clinton, S.M.; Grimm, N.B.; Fisher, S.G. Response of a hyporheic invertebrate assemblage to drying disturbance in a desert stream. J. N. Am. Benthol. Soc. 1996, 15, 700–712. [Google Scholar] [CrossRef]
- Fenoglio, S.; Bo, T.; Bosi, G. Deep interstitial habitat as a refuge for Agabus paludosus (Fabricus) (Coleoptera: Dytiscidae) during summer droughts. Coleopt. Bull. 2006, 60, 37–41. [Google Scholar] [CrossRef]
- Jacobi, G.Z.; Cary, S.J. Winter stoneflies (Plecoptera) in seasonal habitats in New Mexico, USA. J. N. Am. Benthol. Soc. 1996, 15, 690–699. [Google Scholar] [CrossRef]
- Boulton, A.J. Over-summering refuges of aquatic macroinvertebrates in two intermittent streams in central Victoria. Trans. R. Soc. S. Aust. 1989, 113, 23–34. [Google Scholar]
- Bo, T.; Cucco, M.; Fenoglio, S.; Malacarne, G. Colonisation patterns and vertical movements of stream invertebrates in the interstitial zone: A case study in the Apennines, NW Italy. Hydrobiologia 2006, 568, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Bruno, M.C.; Maiolini, B.; Carolli, M.; Silveri, L. Impact of hydropeaking on hyporheic invertebrates in an Alpine stream (Trentino, Italy). Ann. Limnol. Int. J. Lim. 2009, 45, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, J.; Hildrew, A.G. Flow refugia and the microdistribution of lotic invertebrates. J. N. Am. Benthol. Soc. 1993, 12, 385–393. [Google Scholar] [CrossRef]
- Dole-Olivier, M.-J.; Marmonier, P.; Beffy, J.-L. Response of invertebrates to lotic disturbance: Is the hyporheic zone a patchy refugium? Freshw. Biol. 1997, 37, 257–276. [Google Scholar] [CrossRef]
- Gibert, J.; Stanford, J.A.; Dole-Olivier, M.-J.; Ward, J.V. Basic attributes of groundwater ecosystems and prospects for research. In Groundwater Ecology; Gibert, J., Danielopol, D.L., Stanford, J.A., Eds.; Academic Press: San Diego, CA, USA, 1994; pp. 7–40. [Google Scholar]
- Marmonier, P.; Dole, M.J. Les Amphipodes des sédiments d’un bras court-circuité du Rhône: Logique de répartition et réaction aux crues. Rev. Fr. Sci. Eau 1986, 5, 461–486. [Google Scholar]
- Boulton, A.J.; Stibbe, S.E.; Grimm, N.B.; Fisher, S.G. Invertebrate recolonization of small patches of defaunated hyporheic sediments in a Sonoran Desert stream. Freshw. Biol. 1991, 26, 267–277. [Google Scholar] [CrossRef]
- Bruno, M.C.; Bottazzi, E.; Rossetti, G. Downward, upstream or downstream? Assessment of meio- and macrofaunal colonization patterns in a gravel-bed stream using artificial substrates. Ann. Limnol. Int. J. Lim. 2012, 48, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.D.; Hynes, H.B.N. The occurrence of benthos deep in the substratum of a stream. Freshw. Biol. 1974, 4, 233–256. [Google Scholar] [CrossRef]
- Giere, O. Meiobenthology: The Microscopic Motile Fauna of Aquatic Sediments, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Ptatscheck, C.; Gansfort, B.; Majdi, N.; Traunspurger, W. The influence of environmental and spatial factors on benthic invertebrate metacommunities differing in size and dispersal mode. Aquat. Ecol. 2020, 54, 447–461. [Google Scholar] [CrossRef] [Green Version]
- Wasson, J.; Chandesris, A.; Garcia-Bautista, A.; Pella, H.; Villeneuve, B. Rebecca-relationships between ecological and chemical status of surface waters—European Hydro-Ecoregions, EU 6th Framework Programme Contract No. SSPI-CT-2003-502158; Cemagref: Lyon, France, 2007; pp. 1–44. [Google Scholar]
- Falasco, E.; Piano, E.; Doretto, A.; Fenoglio, S.; Bona, F. Lentification in Alpine rivers: Patterns of diatom assemblages and functional traits. Aquat. Sci. 2018, 80, 36. [Google Scholar] [CrossRef]
- Piana, F.; Fioraso, G.; Irace, A.; Mosca, P.; d’Atri, A.; Barale, L.; Falletti, P.; Monegato, G.; Morelli, M.; Tallone, S.; et al. Geology of Piemonte region (NW Italy, Alps–Apennines interference zone). J. Maps 2017, 13, 395–405. [Google Scholar] [CrossRef]
- Doretto, A.; Bona, F.; Falasco, E.; Morandini, D.; Piano, E.; Fenoglio, S. Stay with the flow: How macroinvertebrate communities recover during the rewetting phase in Alpine streams affected by an exceptional drought. River Res. Appl. 2020, 36, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Piano, E.; Doretto, A.; Falasco, E.; Fenoglio, S.; Gruppuso, L.; Nizzoli, D.; Viaroli, P.; Bona, F. If Alpine streams run dry: The drought memory of benthic communities. Aquat. Sci. 2019, 81, 1–14. [Google Scholar] [CrossRef]
- Piano, E.; Doretto, A.; Falasco, E.; Gruppuso, L.; Fenoglio, S.; Bona, F. The role of recurrent dewatering events in shaping ecological niches of scrapers in intermittent Alpine streams. Hydrobiologia 2019, 841, 1–13. [Google Scholar] [CrossRef]
- Dussart, B.H. Les Copépodes des eaux Continentales d’Europe Occidentale. Tome I: Calanoïdes et Harpacticoïdes; N. Boubée et Cie: Paris, France, 1967; pp. 1–500. [Google Scholar]
- Dussart, B.H. Les Copépodes des eaux Continentales d’Europe Occidentale. Tome II: Cyclopoïdes et Biologie; N. Boubée et Cie: Paris, France, 1969; pp. 1–292. [Google Scholar]
- Pesce, G.L.; Galassi, D.M.P. New or rare species of Diacyclops Kiefer, 1927 (Copepoda, Cyclopoida) from different groundwater habitats in Italy. Hydrobiologia 1987, 148, 103–144. [Google Scholar] [CrossRef]
- Pesce, G.L. The genus Diacyclops Kiefer in Italy: A taxonomic, ecological and biogeographical up-to-date review (Crustacea Copepoda Cyclopidae). Arthropoda Sel. 1995, 3, 13–19. [Google Scholar]
- Karaman, G.S. Fauna d’Italia. 31, Crustacea: Amphipoda di Acqua Dolce; Calderini: Bologna, Italy, 1993; pp. 1–337. [Google Scholar]
- Karaytug, S. Copepoda: Cyclopoida. Genera Paracyclops, Ochridacyclops and Key to the Eucyclopinae. Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; SPB Academic Publishing: Hague, The Netherlands, 1999; pp. 1–217. [Google Scholar]
- Van Damme, K.; Brancelj, A.; Dumont, H.J. Adaptations to the hyporheic in Aloninae (Crustacea: Cladocera): Allocation of Alona protzi Hartwig, 1900 and related species to Phreatalona gen. nov. Hydrobiologia 2009, 618, 1–34. [Google Scholar] [CrossRef]
- Stoch, F.; Bruno, M.C. Acanthocyclops magistridussarti sp. nov., from ground waters of peninsular Italy, with comments on the intraspecific variability of the antennary basis ornamentation (Copepoda, Cyclopoida, Cyclopidae). Crustaceana Monogr. 2011, 16, 489–506. [Google Scholar] [CrossRef] [Green Version]
- Campaioli, S.; Ghetti, P.F.S.; Minelli, A.; Ruffo, S. Manuale Per Il Riconoscimento dei Macroinvertebrati delle Acque Dolci Italiane, Volume I; Provincia Autonoma di Trento: Trento, Italy, 1994; pp. 1–357. [Google Scholar]
- Campaioli, S.; Ghetti, P.F.S.; Minelli, A.; Ruffo, S. Manuale per Il Riconoscimento dei Macroinvertebrati delle Acque Dolci Italiane, Volume II; Provincia Autonoma di Trento: Trento, Italy, 1999; pp. 358–484. [Google Scholar]
- Fochetti, R.; Ravizza, C.A.; Tierno De Figueroa, J.M. Fauna d’Italia. 43, Plecoptera; Calderini: Bologna, Italy, 2009; pp. 1–350. [Google Scholar]
- Waringer, J.A.; Graf, W. Atlas of Central European Trichoptera Larvae/Atlas der Mitteleuropäischen Köcherfliegenlarven; Erik Mauch Verlag: Dinkelscherben, Germany, 2011; pp. 1–468. [Google Scholar]
- Ruffo, S.; Stoch, F. Checklist e distribuzione della fauna italiana—con CD-ROM. In Memorie del Museo Civico di Storia Naturale di Verona, 2. Serie, Sezione Scienze della Vita; Memorie del Museo Civico di Storia Naturale di Verona: Verona, Italy, 2005; pp. 1–307. [Google Scholar]
- Clarke, K.R.; Somerfield, P.J.; Chapman, M.G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted bray-curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 2006, 330, 55–80. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008; pp. 1–214. [Google Scholar]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Global Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Poff, N.L.; Olden, J.D.; Vieira, N.K.M.; Finn, D.S.; Simmons, M.P.; Kondratieff, B.C. Functional trait niches of North American lotic insects: Traits-based ecological applications in light of phylogenetic relationships. J. N. Am. Benthol. Soc. 2006, 25, 730–755. [Google Scholar] [CrossRef] [Green Version]
- PRIMER-E Ltd. PRIMER Version 6.1.18 & PERMANOVA+ Version 1.0.8; Plymouth Routines in Multivariate Ecological Research: Plymouth, UK, 2015. [Google Scholar]
- TIBCO Software Inc. Statistica (Data Analysis Software System), Version 13; TIBCO Software Inc.: Palo Alto, CA, USA, 2017. [Google Scholar]
- Cardoso, P.; Mammola, S.; Rigal, F.; Carvalho, J.C. BAT: Biodiversity Assessment Tools. R package version 2.0.1. Available online: https://CRAN.R-project.org/package=BAT (accessed on 16 April 2020).
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Orghidan, T. Ein neuer Lebensraum des unterirdischen Wassers, der hyporheische Biotop. Arch. Hydrobiol. 1959, 55, 392–414. [Google Scholar]
- Wood, P.J.; Boulton, A.J.; Little, S.; Stubbington, R. Is the hyporheic zone a refugium for aquatic macroinvertebrates during severe low flow conditions? Fund. Appl. Limnol. 2010, 176, 377–390. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, S.I.; Hahn, H.J. What is groundwater and what does this mean to fauna?—An opinion. Limnologica 2012, 42, 1–6. [Google Scholar] [CrossRef]
- Stubbington, R.; Greenwood, A.M.; Wood, P.J.; Armitage, P.D.; Gunn, J.; Robertson, A.L. The response of perennial and temporary headwater stream invertebrate communities to hydrological extremes. Hydrobiologia 2009, 630, 299–312. [Google Scholar] [CrossRef]
- Maazouzi, C.; Galassi, D.M.P.; Claret, C.; Cellot, B.; Fiers, F.; Martin, D.; Marmonier, P.; Dole-Olivier, M.-J. Do benthic invertebrates use hyporheic refuges during streambed drying? A manipulative field experiment in nested hyporheic flowpaths. Ecohydrology 2017, 10, e1865. [Google Scholar] [CrossRef] [Green Version]
- Datry, T.; Larned, S.T.; Scarsbrook, M.R. Responses of hyporheic invertebrate assemblages to large-scale variation in flow permanence and surface-subsurface exchange. Freshw. Biol. 2007, 52, 1452–1462. [Google Scholar] [CrossRef]
- Boulton, A.J. The subsurface macrofauna. In Streams and Ground Waters; Jones, J.B., Mulholland, P.J., Eds.; Academic Press: San Diego, CA, USA, 2000; pp. 337–362. [Google Scholar]
- Hancock, P.J.; Boulton, A.J.; Humphreys, W.F. Aquifers and hyporheic zones: Towards an ecological understanding of groundwater. Hydrogeol. J. 2005, 13, 98–111. [Google Scholar] [CrossRef]
- Palmer, M.A.; Bely, A.E.; Berg, K.E. Response of invertebrates to lotic disturbance: A test of the hyporheic refuge hypothesis. Oecologia 1992, 89, 182–194. [Google Scholar] [CrossRef]
- Swan, C.M.; Palmer, M.A. What drives small-scale patterns in lotic meiofauna communities? Freshw. Biol. 2000, 44, 109–121. [Google Scholar] [CrossRef]
- Schmid, P.E. Stochasticity in resource utilisation by a larval Chironomidae (Diptera) community in the bed sediments of a gravel stream. In Groundwater/Surface Water Ecotones: Biological and Hydrological Interactions and Management Options; Gibert, J., Mathieu, J., Fournier, F., Eds.; Academic Press: San Diego, CA, USA, 1997; pp. 21–28. [Google Scholar]
- Robertson, A.L. Lotic meiofaunal community dynamics: Colonisation, resilience and persistence in a spatially and temporally heterogeneous environment. Freshw. Biol. 2000, 44, 135–147. [Google Scholar] [CrossRef]
- Schmid-Araya, J.M. Invertebrate recolonization patterns in the hyporheic zone of a gravel stream. Limnol. Oceanogr. 2000, 45, 1000–1005. [Google Scholar] [CrossRef]
- Dole-Olivier, M.-J.; Marmonier, P. Patch distribution of interstitial communities: Prevailing factors. Freshw. Biol. 1992, 27, 177–191. [Google Scholar] [CrossRef]
- Strayer, D.; May, S.E.; Nielsen, P.; Wolheim, W.; Hausam, S. Oxygen, organic matter, and sediment granulometry as controls on hyporheic animal communities. Arch. Hydrobiol. 1997, 140, 131–144. [Google Scholar] [CrossRef]
- Silver, P.; Palmer, M.A.; Swan, C.M.; Wooster, D. The small scale ecology of freshwater meiofauna. In Freshwater Meiofauna: Biology and Ecology; Rundle, S.D., Robertson, A.L., Schmid-Araya, J.M., Eds.; Backhuys: Leiden, The Netherlands, 2002; pp. 217–239. [Google Scholar]
- Aspin, T.W.H.; Matthews, T.J.; Khamis, K.; Milner, A.M.; Wang, Z.; O’ Callaghan, M.J.; Ledger, M.E. Drought intensification drives turnover of structure and function in stream invertebrate communities. Ecography 2018, 41, 1992–2004. [Google Scholar] [CrossRef] [Green Version]
- Van Looy, K.; Tonkin, J.D.; Floury, M.; Leigh, C.; Soininen, J.; Larsen, S.; Heino, J.; Poff, N.L.; Delong, M.; Jähnig, S.C.; et al. The three Rs of river ecosystem resilience: Resources, recruitment, and refugia. River Res. Applic. 2019, 35, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Boulton, A.J.; Stanley, E.H. Hyporheic processes during flooding and drying in a Sonoran Desert stream. II. Faunal dynamics. Arch. Hydrobiol. 1995, 134, 27–52. [Google Scholar]
- Vander Vorste, R.; Malard, F.; Datry, T. Is drift the primary process promoting the resilience of river invertebrate communities? A manipulative field experiment in an intermittent alluvial river. Freshw. Biol. 2016, 61, 1276–1292. [Google Scholar] [CrossRef]
- Robertson, A.L.; Lancaster, J.; Hildrew, A.G. Stream hydraulics and the distribution of microcrustacea: A role for refugia? Freshw. Biol. 1995, 33, 469–484. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stb—Mean Abundance | Stp—Mean Abundance | Stx—Mean Abundance | Stb—Mean Number of Taxa | Stp—Mean Number of Taxa | Syx—Mean Number of Taxa | |
|---|---|---|---|---|---|---|
| Perm | 0.7 | 0.32 | 0.29 | 13 | 18 | 20 |
| Int-1 | 0.84 | 1.22 | 0.65 | 16 | 13 | 18 |
| Int-3 | 0.23 | 0.21 | 0.14 | 10 | 13 | 15 |
| Rec-no-dwn 2d | 0.17 | 0.45 | 0.21 | 11 | 11 | 18 |
| Rew-no-dwn 2d | 0.60 | 0.42 | 0.26 | 5 | 8 | 9 |
| Rec-dwn 2d | 0.84 | 0.78 | 0.53 | 11 | 8 | 13 |
| Rew-dwn 2d | 0.10 | 0.14 | 0.08 | 6 | 5 | 6 |
| Rec-no-dwn 5d | 0.16 | 0.45 | 0.23 | 11 | 11 | 19 |
| Rew-no-dwn 5d | 0.63 | 0.42 | 0.24 | 5 | 7 | 6 |
| Rec-dwn 5d | 0.18 | 0.43 | 0.14 | 10 | 7 | 7 |
| Rew-dwn 5 d | 1.39 | 0.68 | 0.83 | 7 | 6 | 12 |
| Overall | Stygobites | ||
|---|---|---|---|
| PERMANOVA | PERMANOVA | K-W, M-W | |
| Factor: Station | ns | ns | ns |
| ns | ns | int-3 vs. int-1, p = 0.019 | |
| ns | ns | ns | |
| Factor: Aquifer | p = 0.021 (2 days) | p = 0.016 (2 days) | ns |
| rec-no-dwn vs. rew-no-dwn, p = 0.016 (2 days) | rec-no-dwn vs. rew-no-dwn, p = 0.014 (2 days) | ns | |
| rec-no-dwn vs. rec-dwn, p = 0.013 (2 days) | rec-no-dwn vs. rec-dwn, p = 0.014 (2 days) | ns | |
| Stygophiles | Stygoxenes | ||
| PERMANOVA | K-W, M-W | PERMANOVA | |
| Factor: Station | p =0.01 | ns | p = 0.001 |
| perm vs. int-1, p= 0.012 | perm vs. int-1, p = 0.006, | perm vs. int-1, p = 0.002 | |
| int-3 vs. int-1, p = 0.008 | int-3 vs. int-1, p = 0.006 | int-3 vs. int-1, p = 0.009 | |
| Factor: Aquifer | ns | ns | ns |
| ns | ns | ns | |
| ns | ns | ns | |
| Among Sites | Among Hydrological Phases | |||||
|---|---|---|---|---|---|---|
| Valid N | Kruskal–Wallis H | p-Value | Valid N | Kruskal–Wallis H | p-Value | |
| Resilience | 36 | 5.167 | 0.076 | 20 | 3.237 | 0.198 |
| Resistance | 36 | 6.949 | 0.031 | 20 | 3.787 | 0.151 |
| Resilience BT | 36 | 0.578 | 0.749 | 20 | 1.074 | 0.585 |
| Resilience HR | 36 | 2.743 | 0.254 | 20 | 0.648 | 0.723 |
| Resistance BT | 36 | 0.329 | 0.848 | 20 | 0.290 | 0.865 |
| Resistance HR | 36 | 4.299 | 0.117 | 20 | 2.856 | 0.240 |
| Over all samples | Over all hydrological phases | |||||
| Valid N | Wilcoxon T | p-value | Valid N | Wilcoxon T | p-value | |
| Resist vs. resil | 35 | 36.5 | <0.001 | 19 | 1.5 | <0.001 |
| Resil BT vs. resil HR | 35 | 291.5 | 0.700 | 19 | 61.5 | 0.178 |
| Resist BT vs. resist HR | 34 | 161.5 | 0.020 | 18 | 25.5 | 0.009 |
| Resist BT vs. resil BT | 29 | 12.0 | <0.001 | 17 | 7 | <0.001 |
| Resist HR vs. resil HR | 32 | 0.0 | <0.001 | 17 | 0 | <0.001 |
| Int-1 | Int-3 | |||||
| Valid N | Wilcoxon T | p-value | Valid N | Wilcoxon T | p-value | |
| Resist vs. resil | 5 | 0.0 | 0.043 | 14 | 1.5 | 0.001 |
| Resil BT vs. resil HR | 4 | 2.0 | 0.273 | 15 | 42.5 | 0.320 |
| Resist BT vs. resist HR | 4 | 0.0 | 0.068 | 14 | 20.0 | 0.041 |
| Perm | High flows | |||||
| Valid N | Wilcoxon T | p-value | Valid N | Wilcoxon T | p-value | |
| Resist vs. resil | 16 | 16.0 | 0.007 | 4 | 0.0 | 0.068 |
| Resil BT vs. resil HR | 16 | 32.5 | 0.066 | 3 | 2.0 | 0.593 |
| Resist BT vs. resist HR | 16 | 61.0 | 0.717 | 3 | 2.5 | 0.789 |
| Low flows | Drought | |||||
| Valid N | Wilcoxon T | p-value | Valid N | Wilcoxon T | p-value | |
| Resist vs. resil | 3 | 0.0 | 0.109 | 12 | 1.5 | 0.003 |
| Resil BT vs. resil HR | 3 | 2.0 | 0.593 | 13 | 22.5 | 0.108 |
| Resist BT vs. resist HR | 3 | 0.0 | 0.109 | 12 | 8.5 | 0.017 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruno, M.C.; Doretto, A.; Boano, F.; Ridolfi, L.; Fenoglio, S. Role of the Hyporheic Zone in Increasing the Resilience of Mountain Streams Facing Intermittency. Water 2020, 12, 2034. https://doi.org/10.3390/w12072034
Bruno MC, Doretto A, Boano F, Ridolfi L, Fenoglio S. Role of the Hyporheic Zone in Increasing the Resilience of Mountain Streams Facing Intermittency. Water. 2020; 12(7):2034. https://doi.org/10.3390/w12072034
Chicago/Turabian StyleBruno, Maria Cristina, Alberto Doretto, Fulvio Boano, Luca Ridolfi, and Stefano Fenoglio. 2020. "Role of the Hyporheic Zone in Increasing the Resilience of Mountain Streams Facing Intermittency" Water 12, no. 7: 2034. https://doi.org/10.3390/w12072034
APA StyleBruno, M. C., Doretto, A., Boano, F., Ridolfi, L., & Fenoglio, S. (2020). Role of the Hyporheic Zone in Increasing the Resilience of Mountain Streams Facing Intermittency. Water, 12(7), 2034. https://doi.org/10.3390/w12072034