
Drivers of Macrophyte and Diatom Diversity in a Shallow Hypertrophic Lake

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
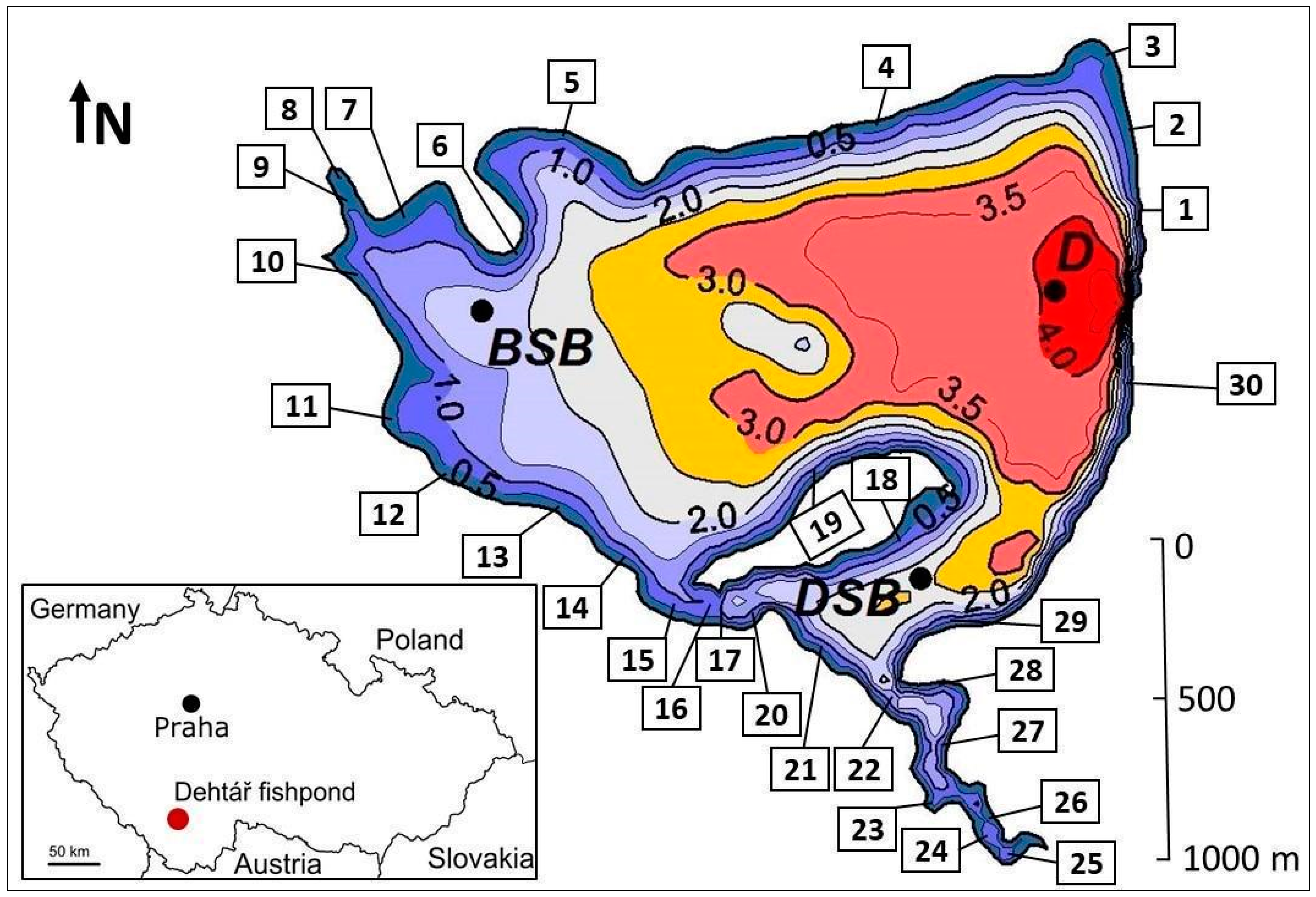
2.1. Study Site
2.2. Methods
2.2.1. Physico-Chemical Parameters of Water
2.2.2. Other Environmental Factors, Fishpond Management Data
2.2.3. Vascular Plant Data Recording
2.2.4. Diatom Sampling and Processing
2.2.5. Data Processing and Analyses
3. Results
3.1. Environmental Parameters
3.2. Vascular Plant and Charophyte Species and Functional Diversity
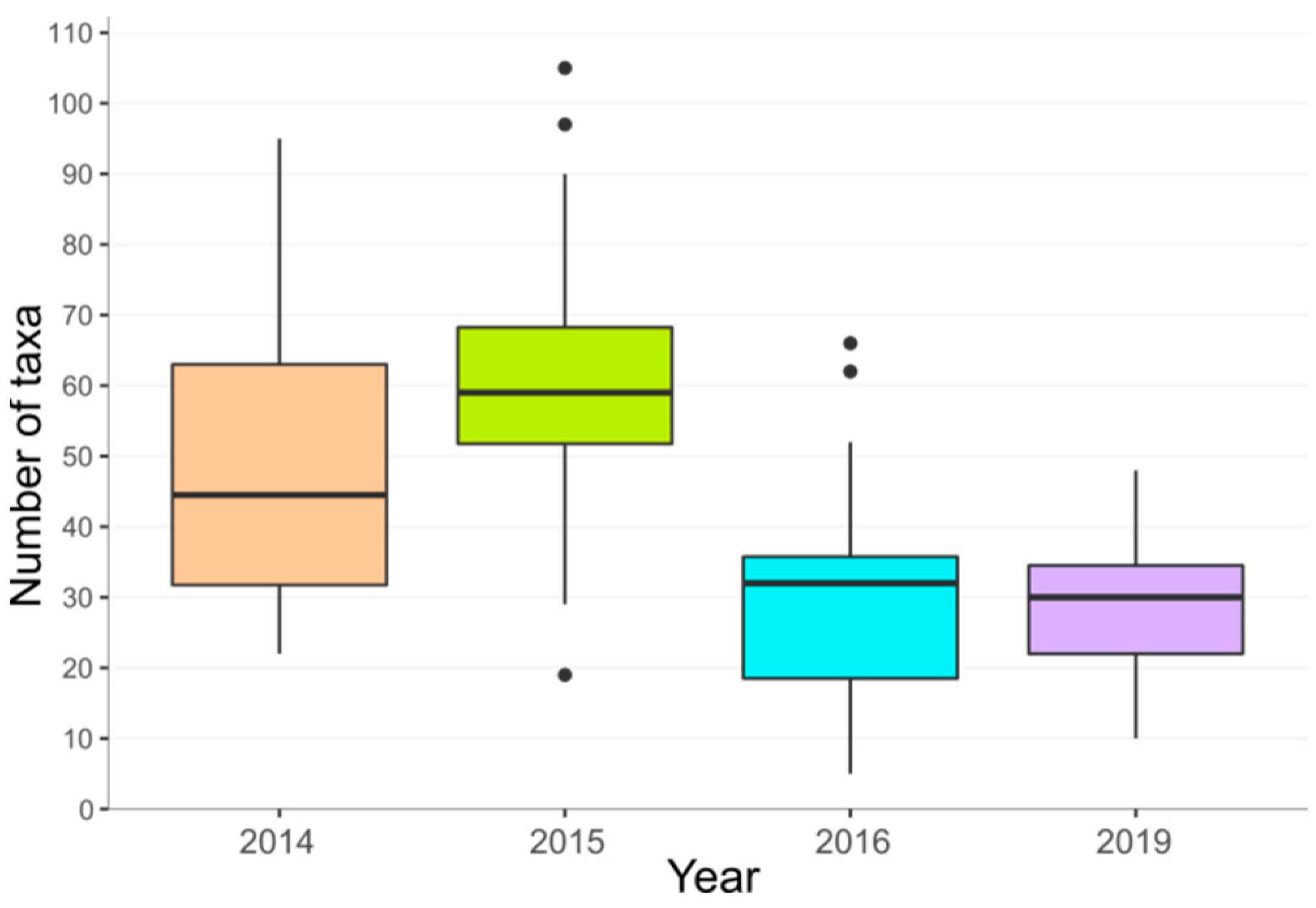
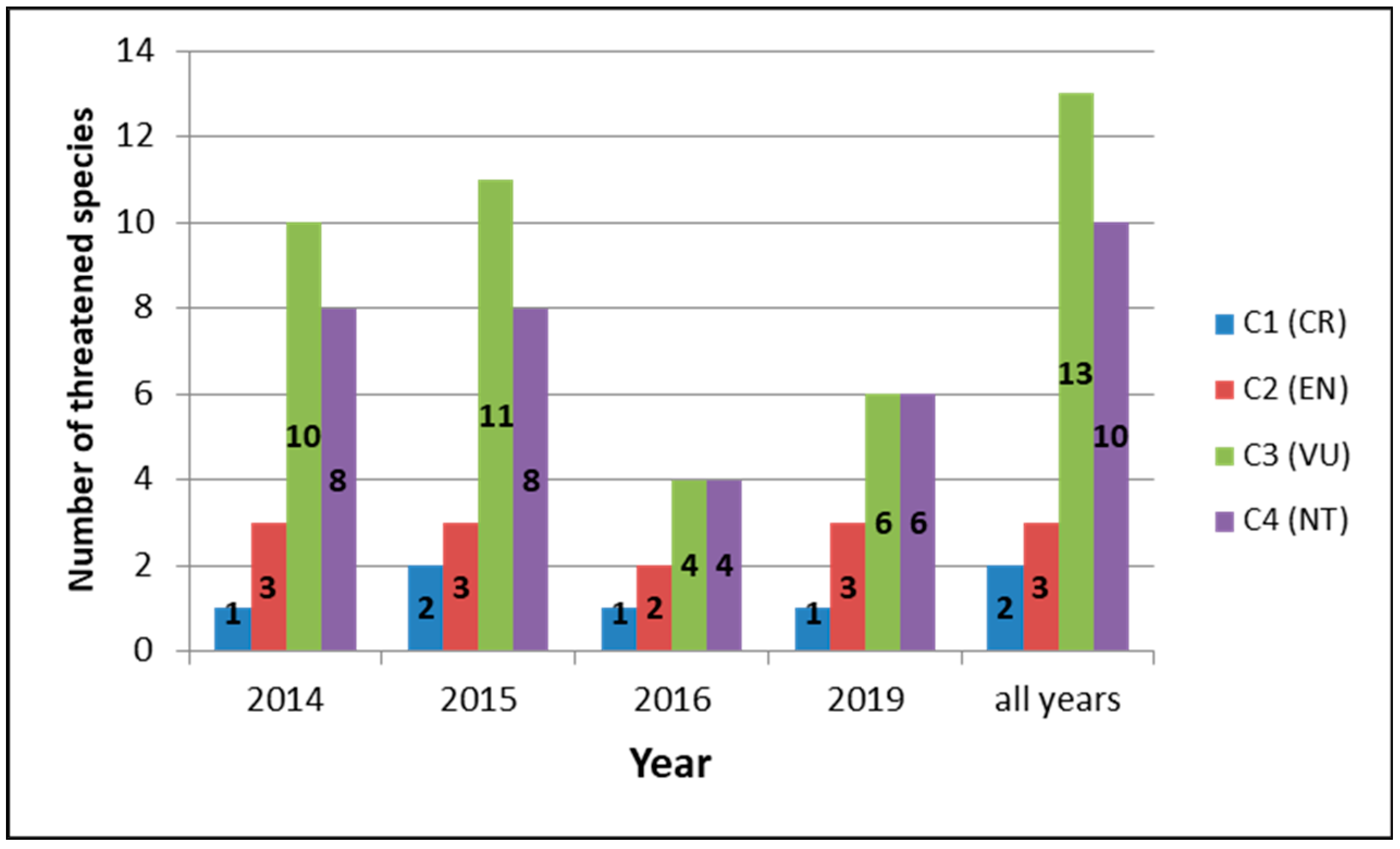
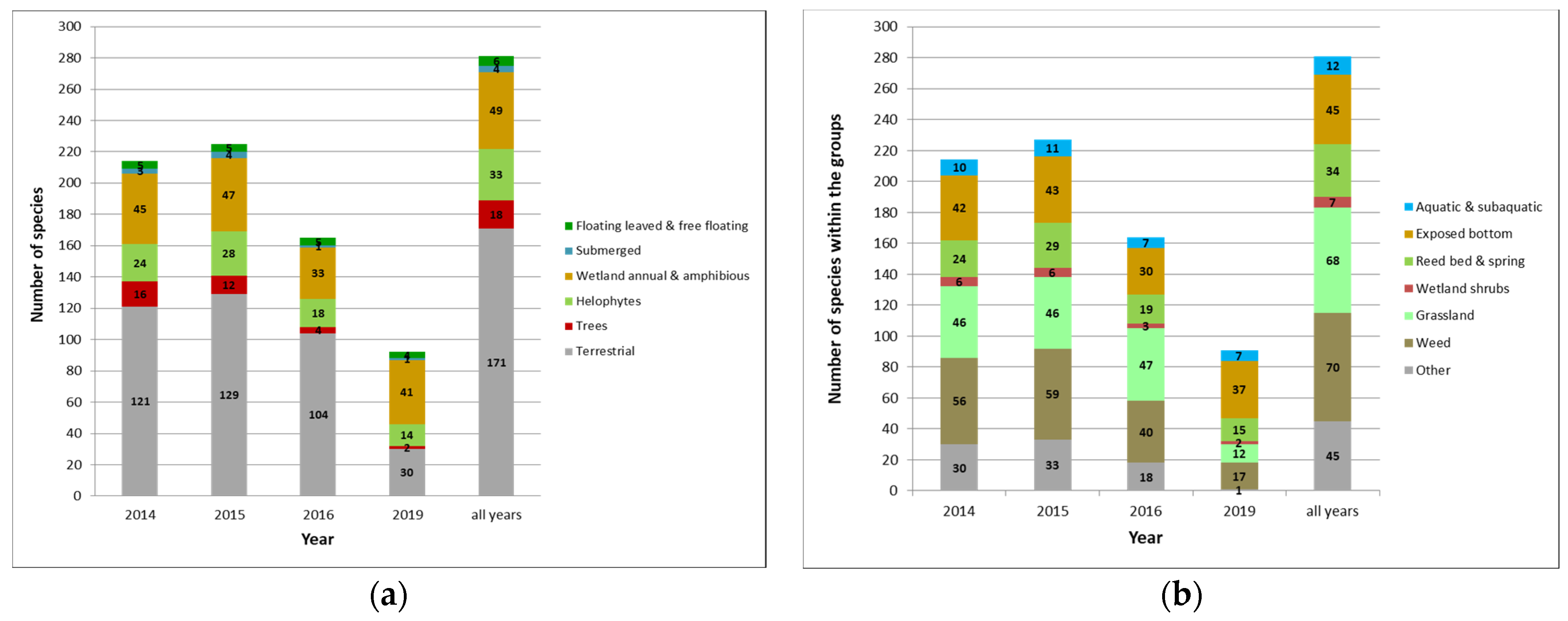
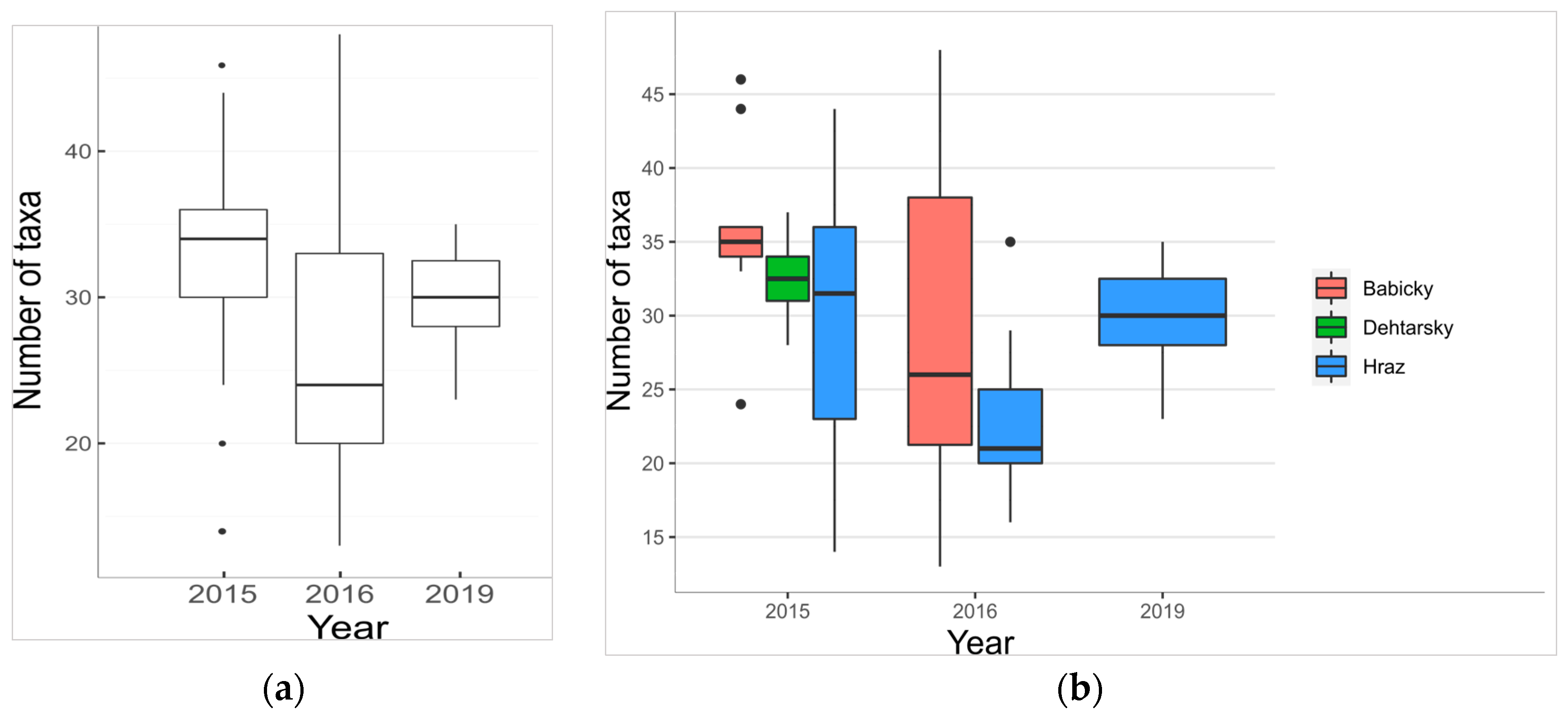
3.2.1. Species Richness, Representation of Threatened Species
3.2.2. Vegetation Structure and Species Composition of Vascular Plant and Charophyte Assemblages
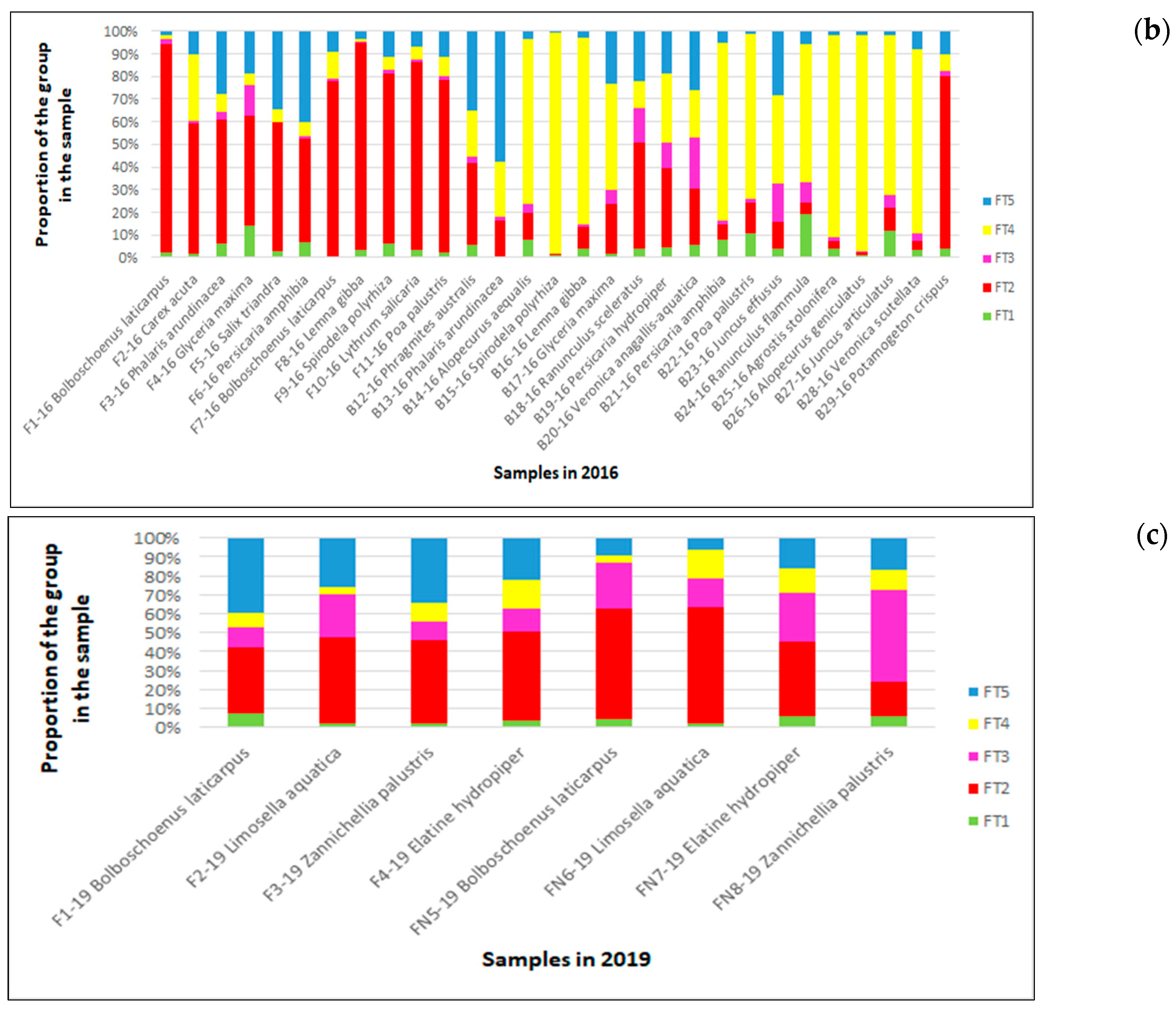
3.2.3. Ecological and Functional Groups of Vascular Plants and Charophytes
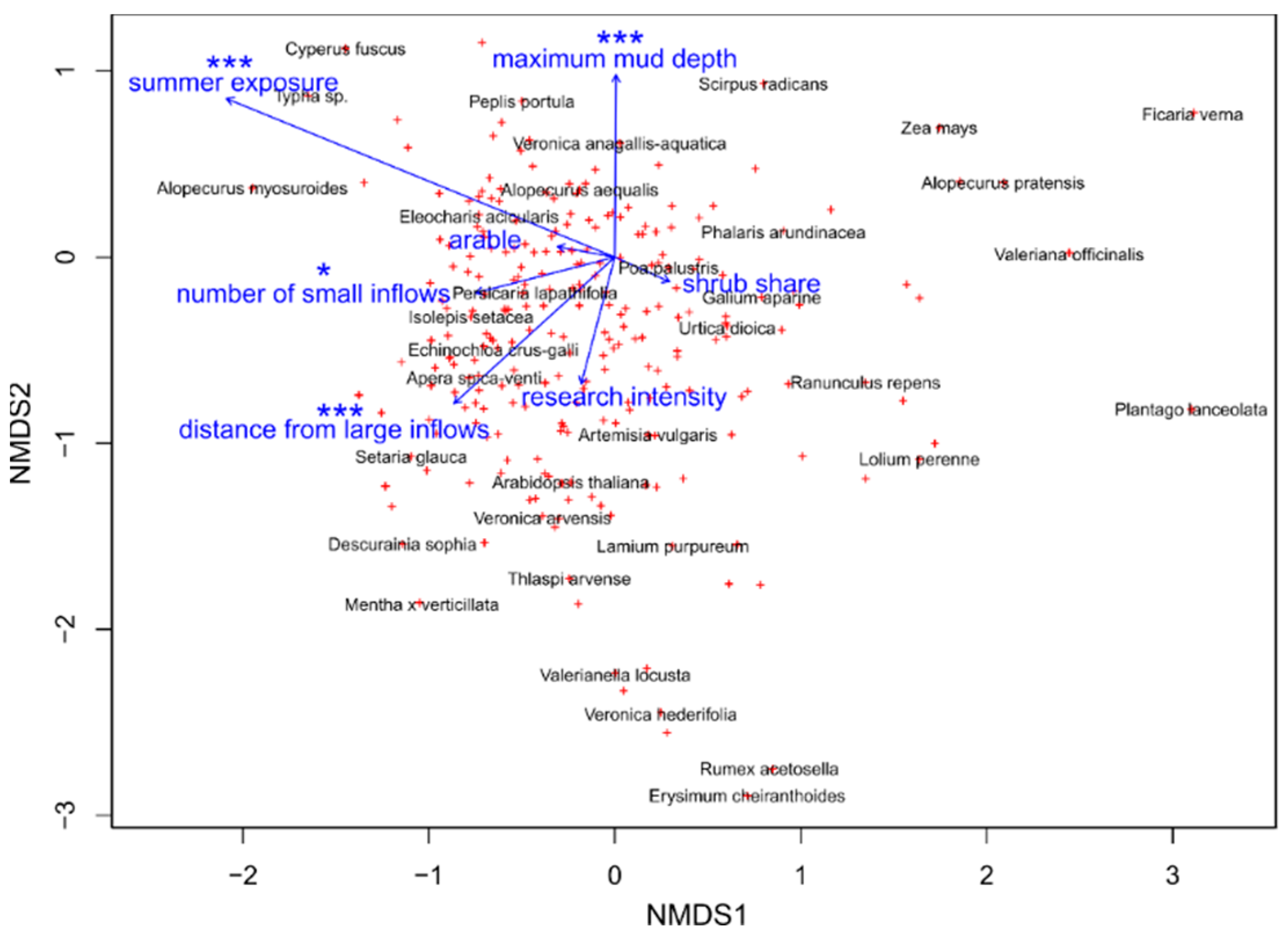
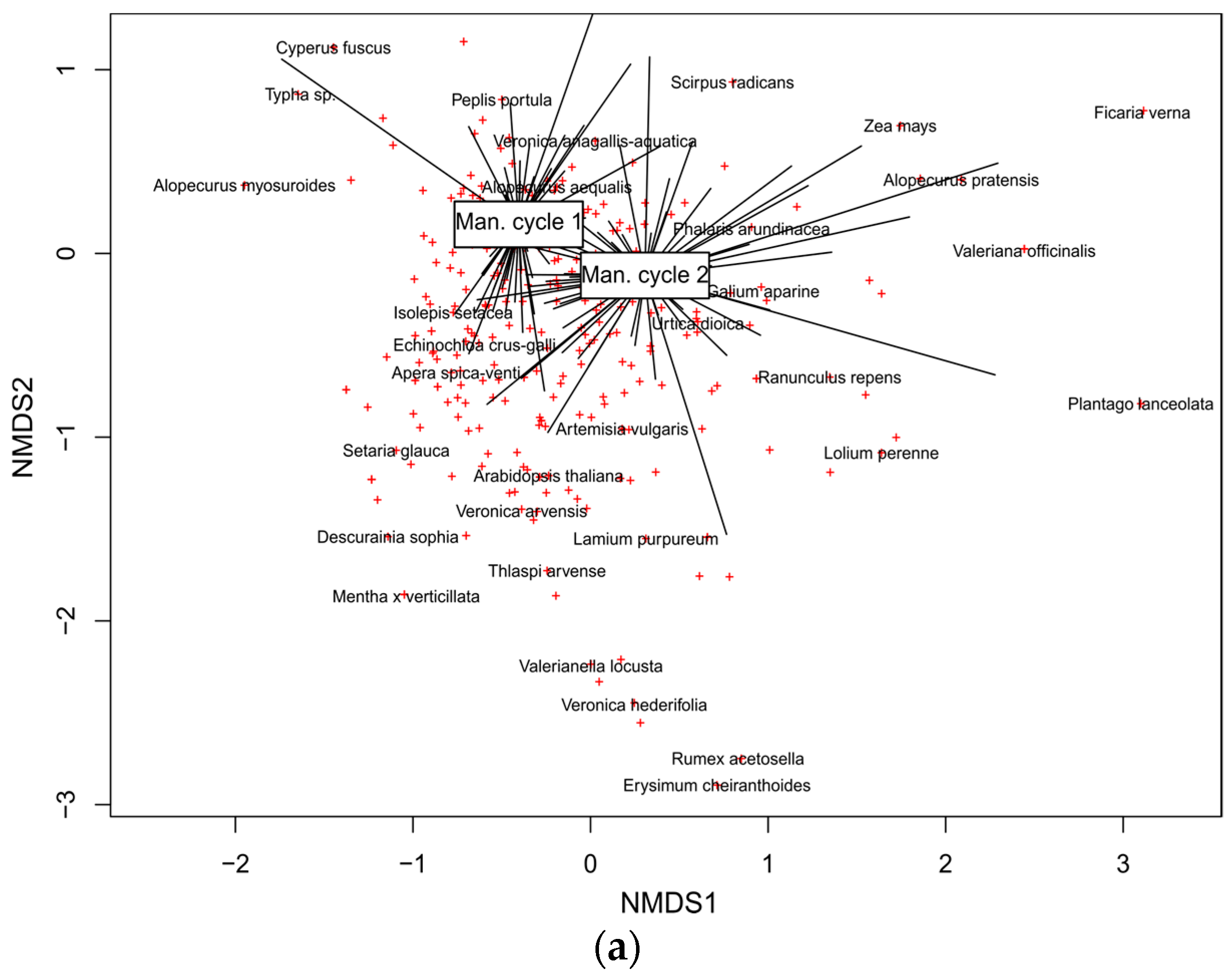
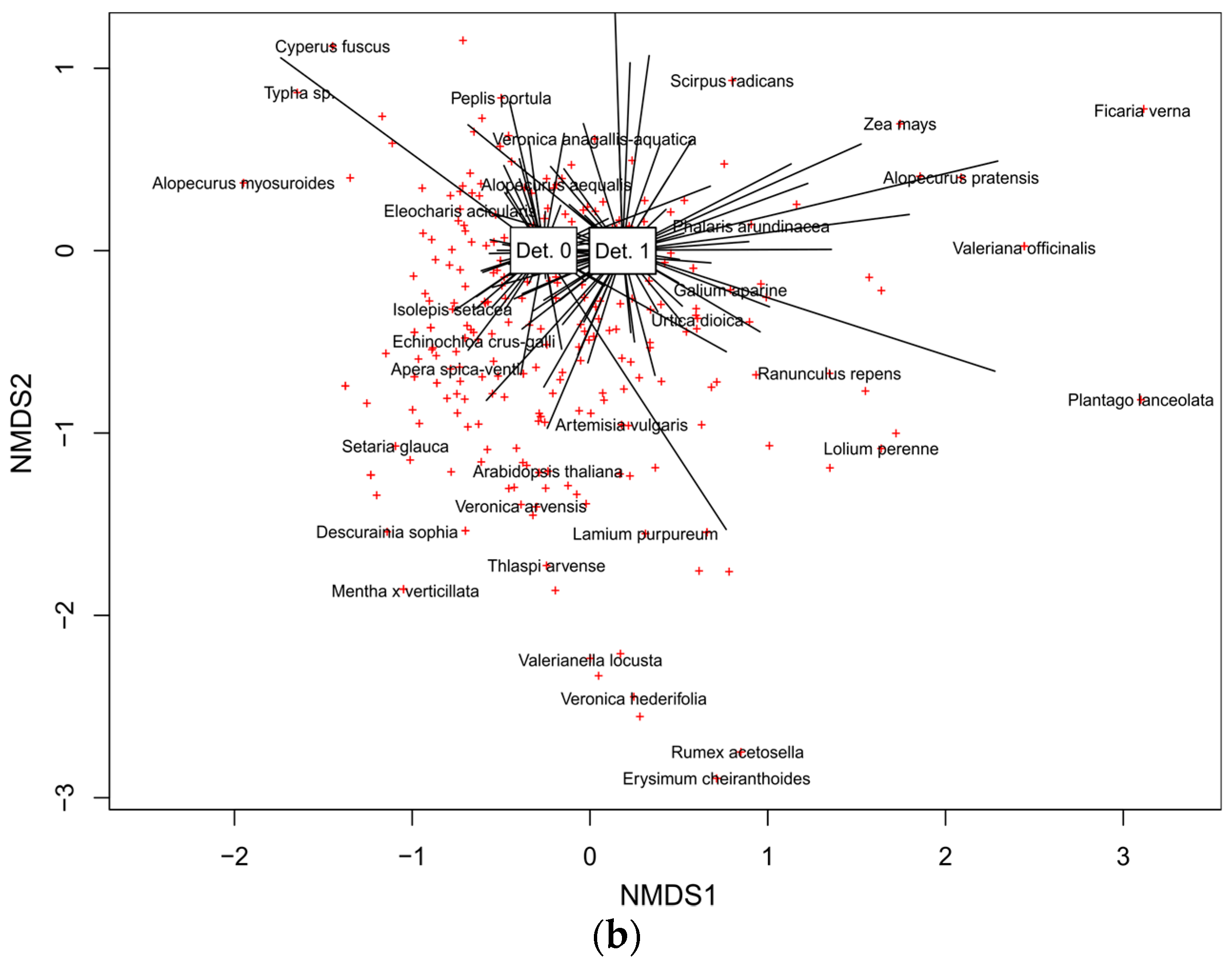
3.2.4. Main Drivers of Species Composition in Vascular Plant and Charophyte Assemblages
3.3. Diatoms
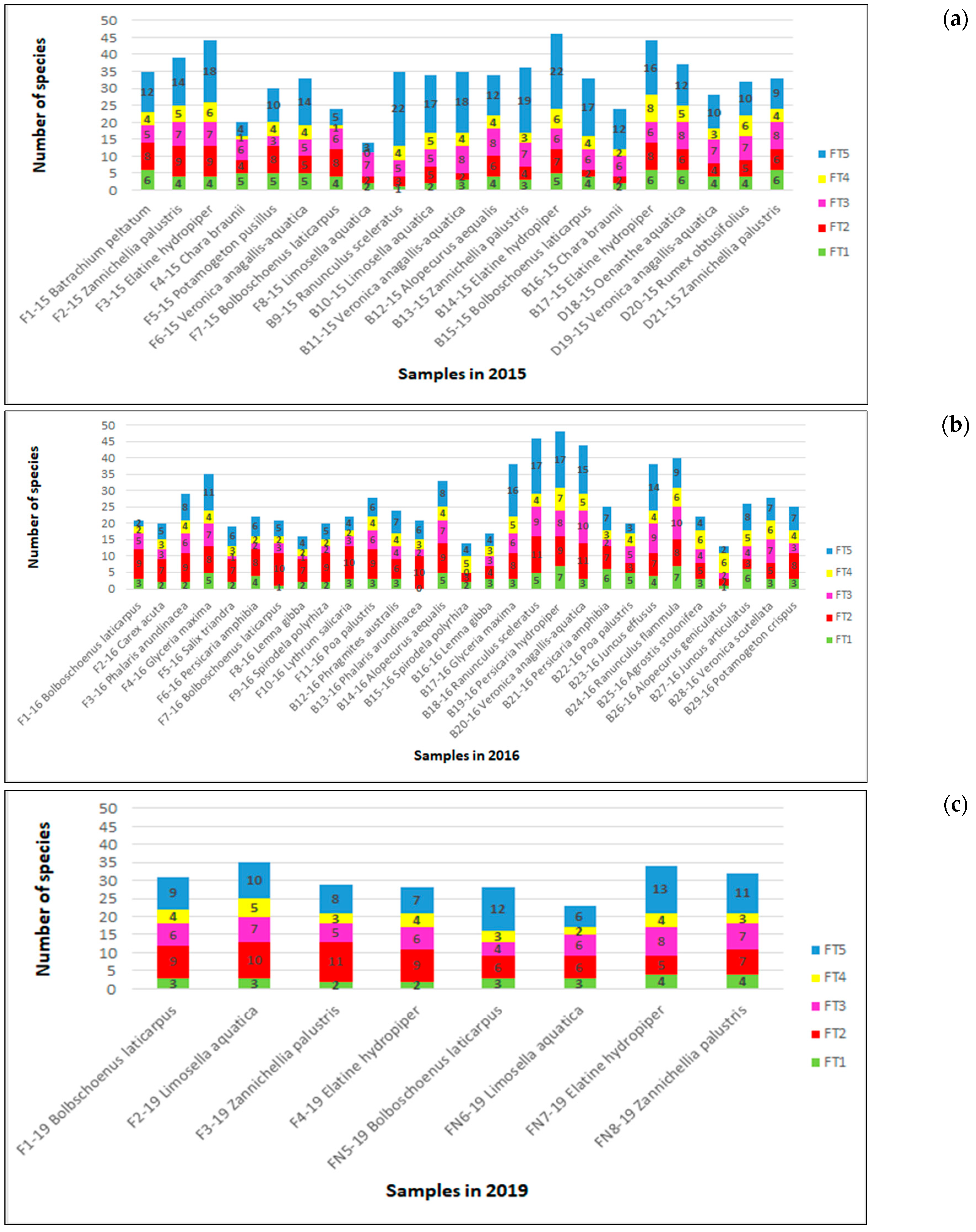
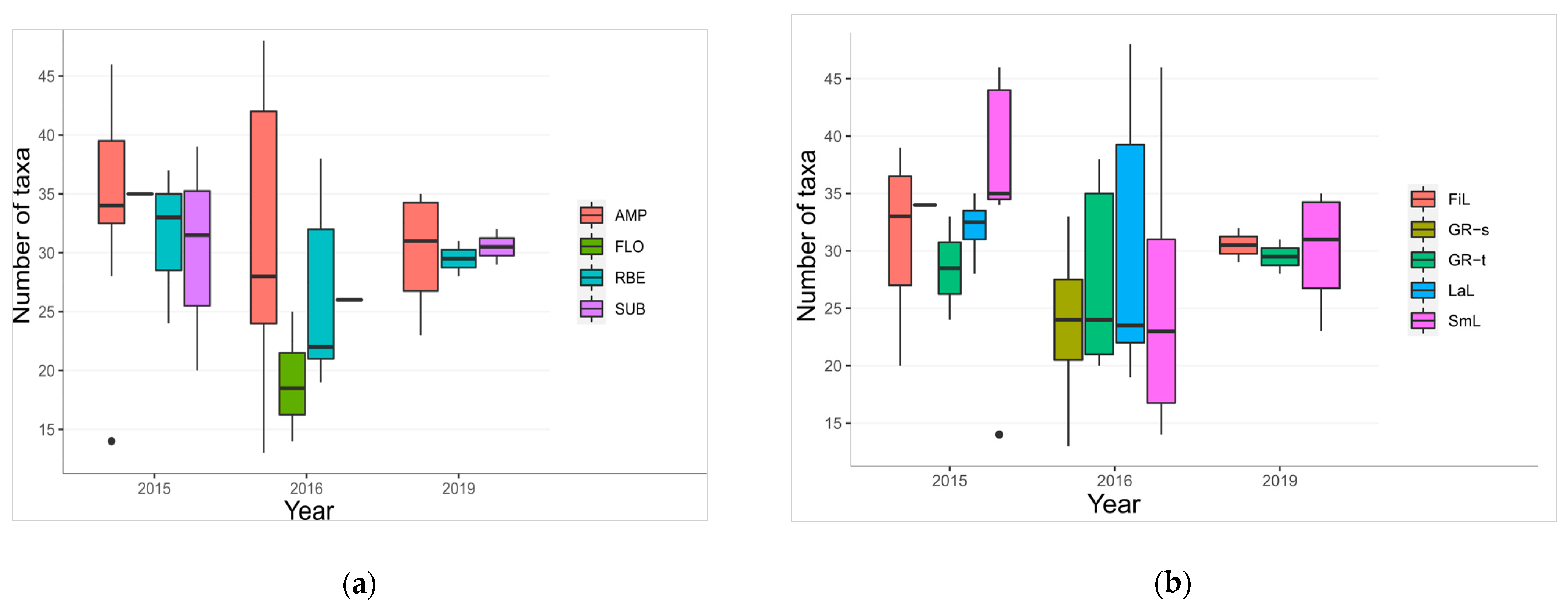
3.3.1. Diatom Species and Functional Group Richness
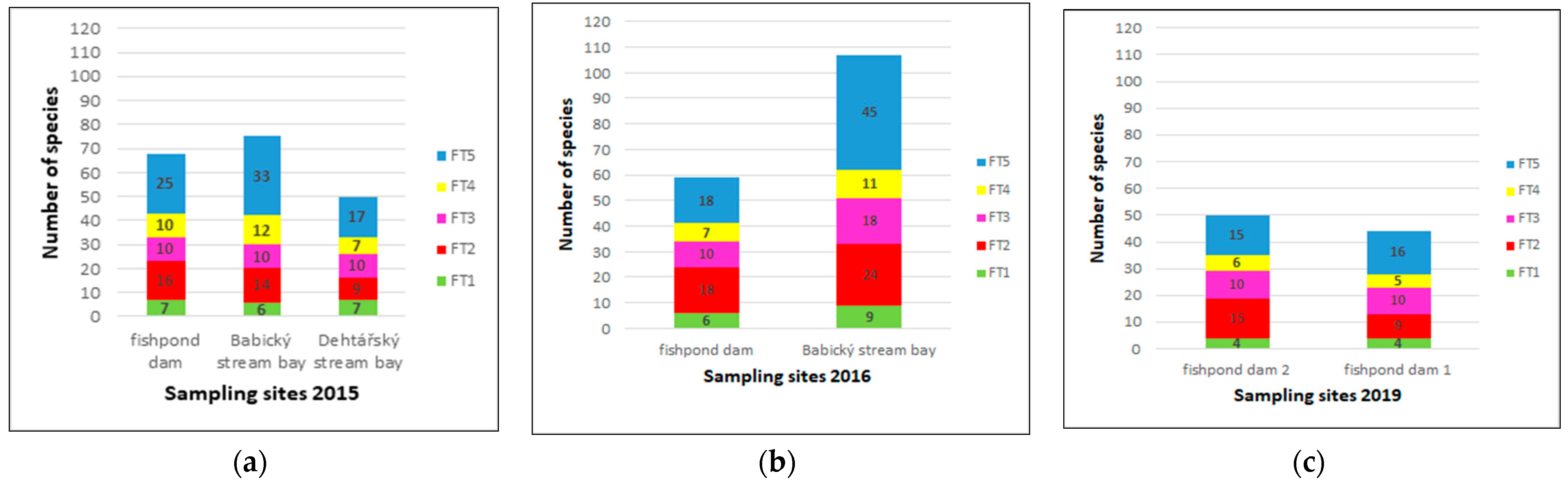
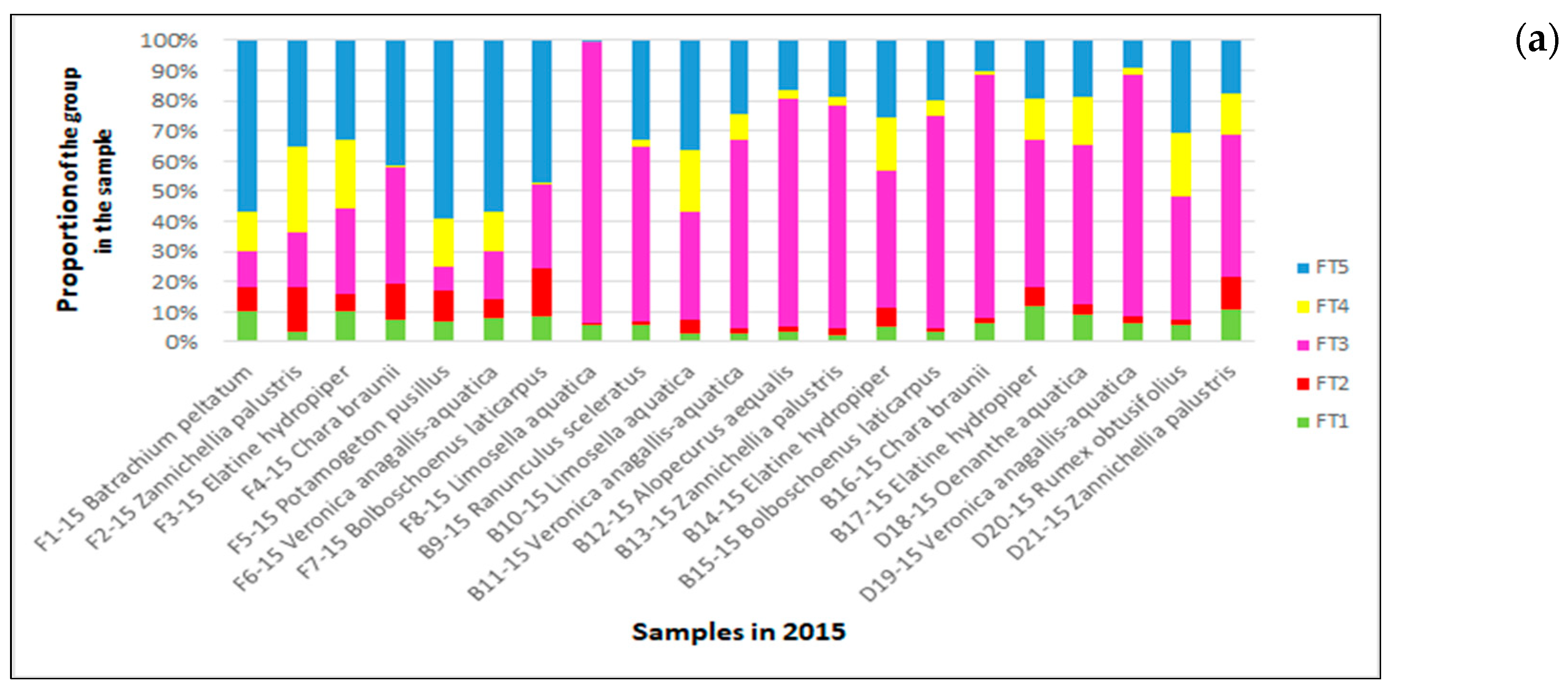
3.3.2. Quantitative Representation of Diatom Species and Their Functional Groups
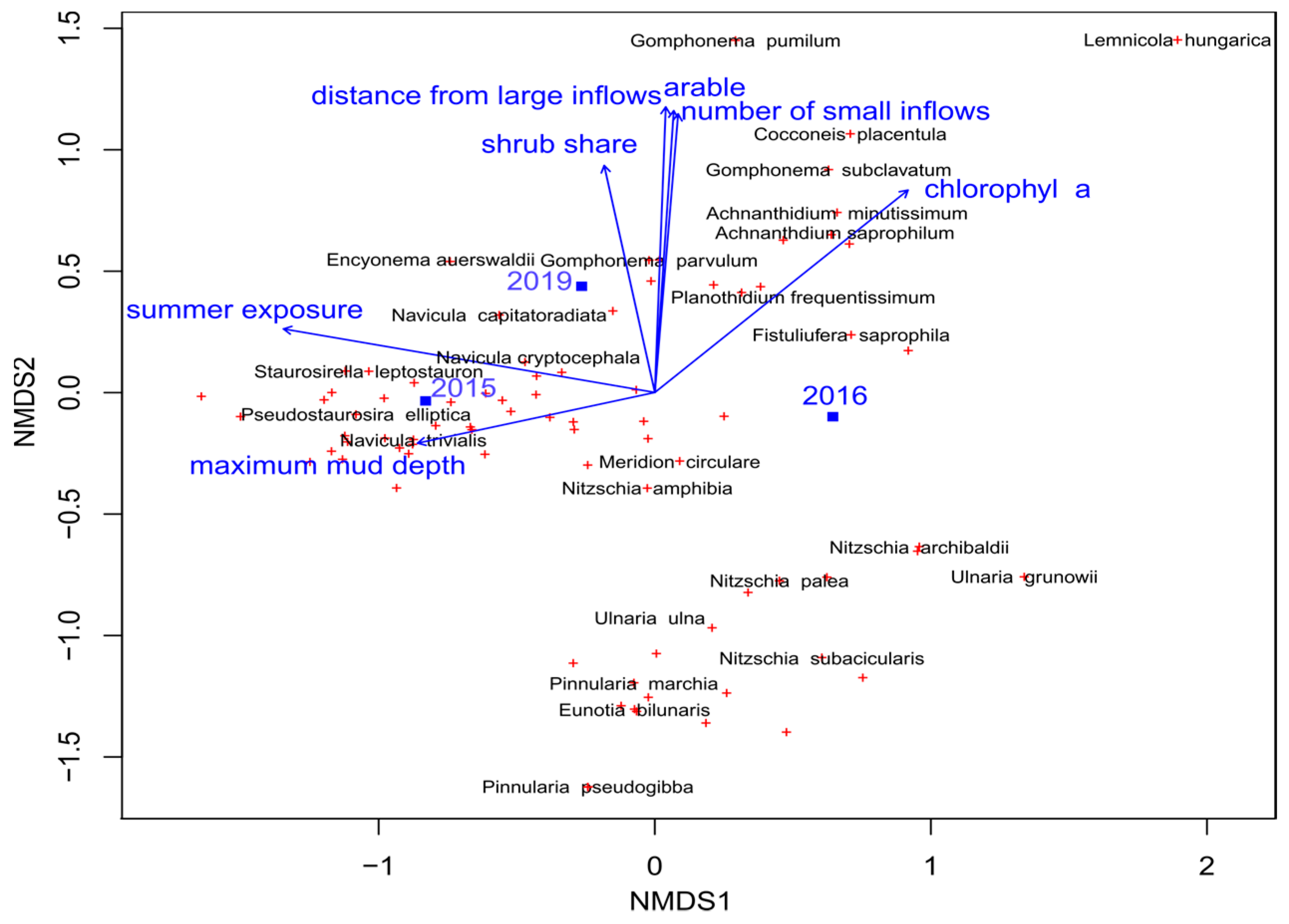
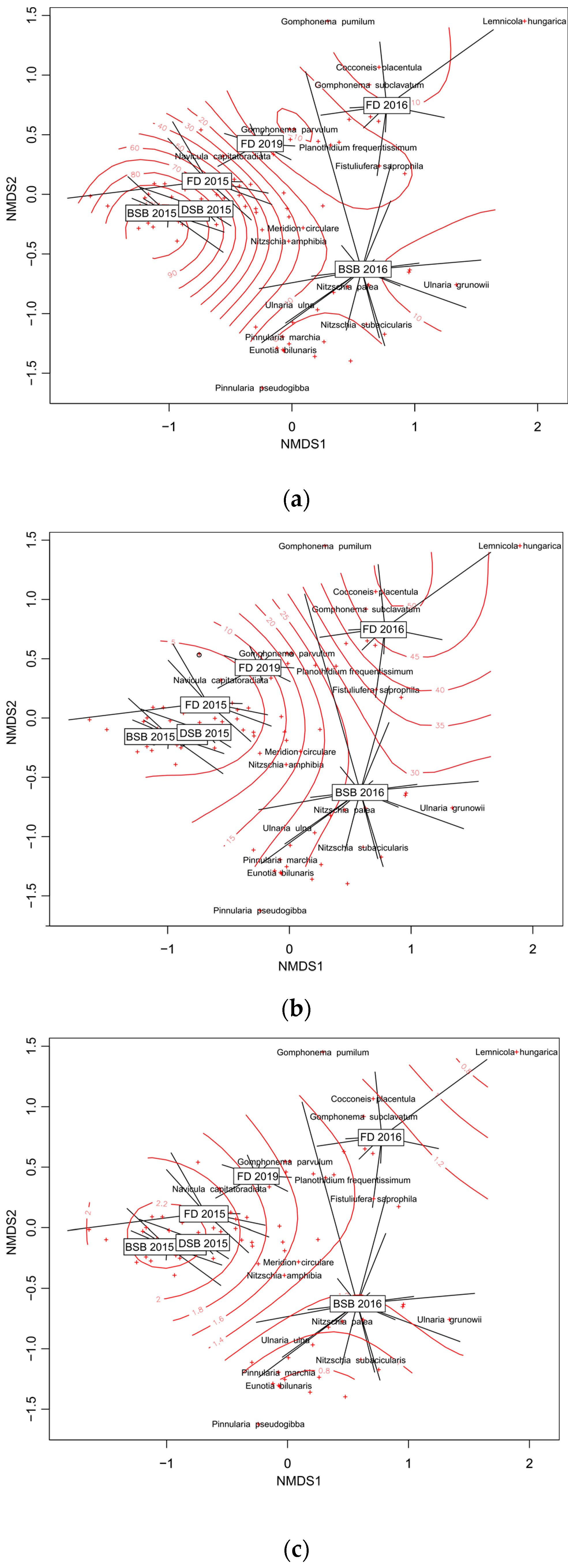
3.3.3. Impact of Environmental Factors on Structure and Species Composition of Diatom Assemblages
3.3.4. Relationships between Macrophytes and Epiphytic Diatom Assemblages
4. Discussion
4.1. Joint Characteristics of Plant and Epiphytic Diatom Assemblages
4.2. Do Macrophytes and Diatoms Follow the Same Environmental Gradients?
4.2.1. Nutritional Gradient
4.2.2. Light Limitation
4.2.3. Moisture and Temperature Fluctuations
4.2.4. Omitted Role of Wave Action in Fishponds
4.3. A Real Biennial Fluctuation or a Trend of Hypertrophy Progression?
4.4. Diatoms and Macrophytes—Do Direct Relationships Exist?
4.5. Importance of Research Intensity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Precipitation—Dehtář (mm) | Precipitation—S Bohemia (mm; Average) | Precipitation Normal—S Bohemia (mm; Average) | Temperatures—Dehtář (°C) | Temperature—S Bohemia (°C; Average) | Temperature Normal—S Bohemia |
|---|---|---|---|---|---|---|
| April 2014 | 21.9 * | 35 | 41 | 10.6 * | 9.0 | 7.2 |
| May 2014 | 116.2 * | 126 | 71 | 12.8 * | 11.3 | 12.5 |
| June 2014 | 41.4 | 33 | 85 | 17.6 * | 15.7 | 15.3 |
| July 2014 | 128.8 | 120 | 92 | 19.7 * | 18.3 | 17.3 |
| August 2014 | 139.2 | 104 | 85 | 16.3 | 15.0 | 16.7 |
| April 2015 | 17.4 | 28 | 41 | 8.1 | 7.2 | 7.2 |
| May 2015 | 59.1 * | 64 | 71 | 13.0 | 12.0 | 12.5 |
| June 2015 | 54 | 68 | 85 | 16.8 | 15.7 | 15.3 |
| July 2015 | 41.4 | 30 | 92 | 21.1 | 20.0 | 17.3 |
| August 2015 | 31.6 | 42 | 85 | 21.0 | 20.5 | 16.7 |
| April 2016 | 33.6 * | 35 | 41 | 8.8 * | 7.1 | 7.2 |
| May 2016 | 108.8 * | 95 | 71 | 13.9 * | 12.5 | 12.5 |
| June 2016 | 88.8 | 94 | 85 | 17.6 | 16.3 | 15.3 |
| July 2016 | 177.7 | 143 | 92 | 19.5 | 18.1 | 17.3 |
| August 2016 | 90.7 | 35 | 85 | 17.9 | 16.4 | 16.7 |
| April 2019 | 10.4 * | 16 | 41 | 10.3 * | 8.6 | 7.2 |
| May 2019 | 74.5 * | 85 | 71 | 11.5 * | 9.9 | 12.5 |
| June 2019 | 85.5 * | 69 | 85 | 21.9 * | 20.0 | 15.3 |
| July 2019 | 104.6 * | 69 | 92 | 20.5 * | 18.6 | 17.3 |
| August 2019 | 76.1 * | 70 | 85 | 20.1 * | 18.3 | 16.7 |
References
- Sand-Jensen, K.; Riis, T.; Vestergaard, O.; Larsen, S.E. Macrophyte decline in Danish lakes and streams over the past 100 years. J. Ecol. 2000, 88, 1030–1040. [Google Scholar] [CrossRef]
- Declerck, S.; Vandekerkhove, J.; Johansson, L.; Muylaert, K.; Conde-Porcuna, J.M.; Van der Gucht, K.; Pérez-Martínez, C.; Lauridsen, T.; Schwenk, K.; Zwart, G.; et al. Multi-group biodiversity in shallow lakes along gradients of phosphorus and water plant cover. Ecology 2005, 86, 1905–1915. [Google Scholar] [CrossRef] [Green Version]
- Alahuhta, J.; Rosbakh, S.; Chepinoga, V.; Heino, J. Environmental determinants of lake macrophyte communities in Baikal Siberia. Aquat. Sci. 2020, 82, 39. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, F.; Hornstrom, E.; Mossberg, P.; Nyberg, P. Ecological effects of lime treatment of acidified lakes and rivers in Sweden. Hydrobiologia 1983, 101, 145–164. [Google Scholar] [CrossRef]
- Murphy, K.J. Plant communities and plant diversity in softwater lakes of northern Europe. Aquat. Bot. 2002, 73, 287–324. [Google Scholar] [CrossRef]
- Bakker, E.S.; Van Donk, E.; Immers, A.K. Lake restoration by in-lake iron addition: A synopsis of iron impact on aquatic organisms and shallow lake ecosystems. Aquat. Ecol. 2016, 50, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Hilt, S.; Alirangues Nuñez, M.M.; Bakker, E.S.; Blindow, I.; Davidson, T.A.; Gillefalk, M.; Hansson, L.-A.; Janse, J.H.; Janssen, A.B.G.; Jeppesen, E.; et al. Response of submerged macrophyte communities to external and internal restoration measures in north temperate shallow lakes. Front. Plant Sci. 2018, 9, 194. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Kronvang, B.; Meerhoff, M.; Søndergaard, M.; Hansen, K.M.; Andersen, H.E.; Lauridsen, T.L.; Liboriussen, L.; Beklioglu, M.; Özen, A.; et al. Climate change effects on runoff, catchment phosphorus loading and lake ecological state, and potential adaptations. J. Environ. Qual. 2009, 38, 1930–1941. [Google Scholar] [CrossRef]
- Liebscher, P.; Rendek, J. Rybníky České Republiky; Academia: Prague, Czech Republic, 2014; pp. 1–584. [Google Scholar]
- Hoffmann, R.C. Economic development and aquatic ecosystems in medieval Europe. Am. Hist. Rev. 1996, 101, 631–669. [Google Scholar] [CrossRef]
- Turoňová, D. (Ed.) Národní přírodní rezervace Novozámecký rybník: Přírodovědecké průzkumy a péče o chráněné území. Příroda 2002, 20, 1–176. [Google Scholar]
- Zákravský, P.; Hroudová, Z. Vliv řízeného rybničního managementu na obnovu rákosin v NPR Velký a Malý Tisý. Zprávy Čes. Bot. Společ. 2007, 42, 167–196. [Google Scholar]
- Miklín, J.; Macháček, P. Birds of Lednické rybníky Fishponds (Czech Republic). J. Maps 2016, 12 (Suppl. 1), 239–248. [Google Scholar] [CrossRef]
- Meerhoff, M.; Jeppesen, E. Shallow lakes and ponds. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Elsevier: Oxford, UK, 2009; Volume 2, pp. 645–655. [Google Scholar]
- Søndergaard, M.; Johansson, L.S.; Levi, E.E.; Lauridsen, T.L.; Jeppesen, E. Lake types and their definition: A case study from Denmark. Inland Waters 2020, 10, 227–240. [Google Scholar] [CrossRef]
- Potužák, J.; Duras, J.; Drozd, B. Mass balance of fishponds: Are they sources or sinks of phosphorus? Aquac. Int. 2016, 24, 1725–1745. [Google Scholar] [CrossRef]
- Rutegwa, M.; Potužák, J.; Hejzlar, J.; Drozd, B. Carbon metabolism and nutrient balance in a hypereutrophic semiintensive fishpond. Knowl. Manag. Aquat. Ecosyst. 2019, 420, 49. [Google Scholar] [CrossRef] [Green Version]
- Šumberová, K.; Lososová, Z.; Fabšicová, M.; Horáková, V. Variability of vegetation of exposed pond bottoms in relation to management and environmental factors. Preslia 2006, 78, 235–252. [Google Scholar]
- Francová, K.; Šumberová, K.; Kučerová, A.; Čtvrtlíková, M.; Šorf, M.; Borovec, J.; Drozd, B.; Janauer, G.A.; Vrba, J. Macrophyte assemblages in fishponds under different fish farming management. Aquat. Bot. 2019, 159, 103131. [Google Scholar] [CrossRef]
- Francová, K.; Šumberová, K.; Kučerová, A.; Šorf, M.; Grill, S.; Exler, N.; Vrba, J. Drivers of plant species composition of ecotonal vegetation in two fishpond management types. Wetl. Ecol. Manag. 2021, 29, 93–100. [Google Scholar] [CrossRef]
- Schäfer-Guignier, O. Weiher in der Franche-Comté: Eine floristisch-ökologische und vegetationskundliche Untersuchung. Diss. Bot. 1994, 213, 1–239. [Google Scholar]
- Wezel, A.; Oertli, B.; Rosset, V.; Arthaud, F.; Leroy, B.; Smith, R.; Angélibert, S.; Bornette, G.; Vallod, D.; Robin, J. Biodiversity patterns of nutrient-rich fish ponds and implications for conservation. Limnology 2014, 15, 213–223. [Google Scholar] [CrossRef]
- Bubíková, K.; Hrivnák, R. Artificial ponds in Central Europe do not fall behind the natural ponds in terms of macrophyte diversity. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 8. [Google Scholar] [CrossRef] [Green Version]
- Waldon, B. The conservation of small water reservoirs in the Krajeńskie Lakeland (North-West Poland). Limnologica 2012, 42, 320–327. [Google Scholar] [CrossRef]
- Scheffer, M.; van Nes, E.H. Shallow lakes theory revisited: Various alternative regimes driven by climate, nutrients, depth and lake size. Hydrobiologia 2007, 584, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Waldon-Rudzionek, B. Is the flora of oxbow lakes different from that of fishponds? A comparison of two types of water reservoirs in the Noteć River valley and Bydgoszcz Canal valley (NW Poland). Ecol. Quest. 2017, 25, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Broyer, J.; Curtet, L. Biodiversity and fish farming intensification in French fishpond systems. Hydrobiologia 2012, 694, 205–218. [Google Scholar] [CrossRef]
- Müller-Stoll, W.R.; Pietsch, W. Ökologische Untersuchungen über die Gesellschaft des Eleocharito-Caricetum bohemicae auf wasserfrei gewordenen Teichböden in Zentraleuropa. Verh. Zool. Bot. Ges. Osterr. 1985, 123, 51–70. [Google Scholar]
- Richert, E.; Achtziger, R.; Dajdok, Z.; Günther, A.; Heilmeier, H.; Hübner, A.; John, H.; Šumberová, K. Rare wetland grass Coleanthus subtilis in Central and Western Europe—Current distribution, habitat types, and threats. Acta Soc. Bot. Pol. 2016, 85, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Poschlod, P.; Rosbakh, S. Mudflat species: Threatened or hidden? An extensive seed bank survey of 108 fish ponds in Southern Germany. Biol. Conserv. 2018, 225, 154–163. [Google Scholar] [CrossRef]
- Zając, M.; Zając, A. Zbiorowiska z klasy Isoëto-Nanojuncetea na dnach wysychających stawów w południowej części Kotliny Oświęcimskiej. Zesz. Nauk. Uniw. Jagiell. Pr. Bot. 1988, 17, 155–160. (In Polish) [Google Scholar]
- Bernhardt, K.-G.; Koch, M.; Kropf, M.; Ulbel, E.; Webhofer, J. Comparison of two methods characterising the seed bank of amphibious plants in submerged sediments. Aquat. Bot. 2008, 88, 171–177. [Google Scholar] [CrossRef]
- Kúr, P.; Pachschwöll, C.; Štech, M. Notes on the distribution of Spergularia echinosperma and the newly recognized species S. kurkae in Austria. Neilreichia 2018, 9, 269–282. [Google Scholar]
- Lemmens, P.; Mergeay, J.; De Bie, T.; Van Wichelen, J.; De Meester, L.; Declerck, S.A. How to maximally support local and regional biodiversity in applied conservation? Insights from pond management. PLoS ONE 2013, 8, e72538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hroudová, Z.; Zákravský, P. Vegetation dynamics in a fishpond littoral related to human impact. Hydrobiologia 1999, 415, 139–145. [Google Scholar] [CrossRef]
- Grzywna, A.; Sender, J.; Bronowicka-Mielniczuk, U. Physical and chemical variables, species composition and coverage of macrophytes in ponds (case study in Eastern Poland). Appl. Ecol. Environ. Res. 2018, 16, 2129–2139. [Google Scholar] [CrossRef]
- Potužák, J.; Hůda, J.; Pechar, L. Changes in fish production effectivity in eutrophic fishponds—Impact of zooplankton structure. Aquac. Int. 2007, 15, 201–210. [Google Scholar] [CrossRef]
- Duras, J.; Potužák, J.; Marcel, M.; Pechar, L. Rybníky a jakost vody. Vodní Hospodářství 2015, 7, 16–24. [Google Scholar]
- Potužák, J.; Šumberová, K.; Fránková, M.; Fabšičová, M.; Ducháček, M.; Císař, K.; Duras, J. Tříleté dobrodružství výzkumu rybníka Dehtář aneb s rybníkem mezi “jezerní” badatele projektu NETLAKE/Three years long investigation of fishpond Dehtář—First fishpond in the NETLAKE COST ACTION project. In Rybníky 2017, Sborník Příspěvků Odborné Konference, 15–16 June 2017, Česká Zemědělská Univerzita, Praha, Czech Republic; David, V., Davidová, T., Eds.; Česká Společnost Krajinných Inženýrů (ČSKI): Prague, Czech Republic, 2017; pp. 73–81. [Google Scholar]
- Kubelka, V.; Havlíček, J.; Mikeš, V. (Eds.) Zajímavá a vzácná ornitologická pozorování v jižních Čechách IV. Sbor. Jihočes. Muz. Čes. Budějovicích, Přír. Vědy 2017, 57, 99–125. [Google Scholar]
- Hroudová, Z.; Kloubec, B.; Zákravský, P. Dynamika litorální vegetace a avifauny Opatovického rybníka u Třeboně. Příroda 1999, 14, 73–98. [Google Scholar]
- Jupp, B.P.; Spence, D.H.N. Limitations on macrophytes in a eutrophic lake, Loch Leven: II. Wave action, sediments and waterfowl grazing. J. Ecol. 1977, 65, 431–446. [Google Scholar] [CrossRef]
- Azza, N.; van de Koppel, J.; Denny, P.; Kansiime, F. Shoreline vegetation distribution in relation to wave exposure and bay characteristics in a tropical great lake, Lake Victoria. J. Trop. Ecol. 2007, 23, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Keddy, P.A. Quantifying within-lake gradients of wave energy: Interrelation- ships of wave energy, substrate particle size and shoreline plants in Axe Lake, Ontario. Aquat. Bot. 1982, 14, 41–55. [Google Scholar] [CrossRef]
- Kolář, J.; Kučerová, A.; Jakubec, P.; Vymazal, J. Seed bank of Littorella uniflora (L.) Asch. in the Czech Republic, Central Europe: Does burial depth and sediment type influence seed germination? Hydrobiologia 2017, 794, 347–358. [Google Scholar] [CrossRef]
- Lepší, M.; Boublík, K.; Štech, M.; Hans, V. (Eds.) Červená kniha květeny jižní části Čech; Jihočeské muzeum v Českých Budějovicích: České Budějovice, Czech Republic, 2013; pp. 235–236, 333. (In Czech) [Google Scholar]
- Jupp, B.P.; Spence, D.H.N. Limitations on macrophytes in a eutrophic lake, Loch Leven: I. Effects of phytoplankton. J. Ecol. 1977, 65, 175–186. [Google Scholar] [CrossRef]
- Morris, P.A.; Hill, N.M.; Reekie, E.G.; Hewlin, H.L. Lakeshore diversity and rarity relationships along interacting disturbance gradients: Catchment area, wave action and depth. Biol. Conserv. 2002, 106, 79–90. [Google Scholar] [CrossRef]
- Poulíčková, A.; Hájková, P.; Křenková, P.; Hájek, M. Distribution of diatoms and bryophytes on linear transects through spring fens. Nova Hedwigia 2004, 78, 411–424. [Google Scholar] [CrossRef]
- Fránková, M.; Bojková, J.; Poulíčková, A.; Hájek, M. The structure and species richness of the diatom assemblages of the Western Carpathian spring fens along the gradient of mineral richness. Fottea 2009, 9, 355–368. [Google Scholar] [CrossRef] [Green Version]
- Marvan, P.; Komárek, J.; Ettl, H.; Komárková, J. Structure and functioning of algal communities in fishponds. 4.1. Structural Elements. Principal Populations of Algae. Spatial Distribution. In Pond Littoral Ecosystems. Structure and Functioning. Methods and Results of Quantitative Ecosystem Research in the Czechoslovakian IBP Wetland Project; Dykyjová, D., Květ, J., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1978; pp. 295–313. [Google Scholar]
- Kollár, J.; Fránková, M.; Hašler, P.; Letáková, M.; Poulíčková, A. Epiphytic diatoms in lotic and lentic waters—Diversity and representation of species complexes. Fottea 2015, 15, 259–271. [Google Scholar] [CrossRef] [Green Version]
- Mutinová, P.T.; Neustupa, J.; Bevilacqua, S.; Terlizzi, A. Host specificity of epiphytic diatom (Bacillariophyceae) and desmid (Desmidiales) communities. Aquat. Ecol. 2016, 50, 697–709. [Google Scholar] [CrossRef]
- Letáková, M.; Poulíčková, A. Epifytické rozsivky vybraných rybníků v České republice. Zpr. Vlastiv. Muz. Olomouci 2017, 313, 44–58. [Google Scholar]
- Fránková, M.; Šumberová, K.; Potužák, J.; Vild, O. The role of plant substrate type in shaping the composition and diversity of epiphytic diatom assemblages in a eutrophic reservoir. Fundam. Appl. Limnol. 2017, 189, 117–135. [Google Scholar] [CrossRef]
- Caput, K.; Plenković-Moraj, A. Epiphytic diatoms on sawgrass (Cladium mariscus) in the karstic Plitvice lakes (Croatia). Biologia 2000, 55, 343–350. [Google Scholar]
- Cejudo-Figueiras, C.; Álvarez-Blanco, I.; Bécares, E.; Blanco, S. Epiphytic diatoms and water quality in shallow lakes: The neutral substrate hypothesis revisited. Mar. Freshwater Res. 2010, 61, 1457–1467. [Google Scholar] [CrossRef]
- Kapetanović, T.; Jahn, R.; Redžić, S.; Carić, M. Diatoms in a poor fen of Bijambare protected landscape, Bosnia & Herzegovina. Nova Hedwigia 2011, 93, 125–151. [Google Scholar]
- Blanco, S.; Cejudo-Figueiras, C.; Álvarez-Blanco, I.; van Donk, E.; Gross, E.M.; Hansson, L.-A.; Irvine, K.; Jeppesen, E.; Kairessalo, T.; Moss, B.; et al. Epiphytic diatoms along environmental gradients in western European shallow lakes. CLEAN Soil Air Water 2014, 42, 229–235. [Google Scholar] [CrossRef]
- Letáková, M.; Cantonati, M.; Hašler, P.; Nicola, A.; Poulíčková, A. Substrate specificity and fine-scale distribution of epiphytic diatoms in a shallow tarn in the Brenta Dolomites (south-eastern Alps). Plant Ecol. Evol. 2016, 149, 144–156. [Google Scholar] [CrossRef]
- Letáková, M.; Fránková, M.; Poulíčková, A. Ecology and applications of freshwater epiphytic diatoms—Review. Cryptogam. Algol. 2018, 39, 3–22. [Google Scholar] [CrossRef]
- Evtimova, V.V.; Donohue, I. Quantifying ecological responses to amplified water level fluctuations in standing waters: An experimental approach. J. Appl. Ecol. 2014, 51, 1282–1291. [Google Scholar] [CrossRef]
- Česká Geologická Služba. Geovědní Mapy 1:25 000. Available online: https://mapy.geology.cz/geocr25 (accessed on 25 March 2021).
- Geoportal Inspire. Půdní Mapa ČR 1:250 000—Klasifikace dle TKSP a WRB. Available online: https://geoportal.gov.cz/web/guest/map (accessed on 25 March 2021).
- Organization for Economic Cooperation and Development (OECD). Eutrophication of Waters. Monitoring, Assessment and Control; Final Report; OECD Cooperative Programme on Monitoring of Inland Waters (Eutrophication Control); OECD: Paris, France, 1982; pp. 1–154. [Google Scholar]
- Hartman, P.; Přikryl, I.; Štědronský, E. Hydrobiologie; Informatorium: Prague, Czech Republic, 2005; pp. 1–359. [Google Scholar]
- Klepal, V.; Hluboká nad Vltavou Fish Farm, Hluboká nad Vltavou, Czech Republic. Personal communication, 2016.
- Francová, K.; Šumberová, K.; Janauer, G.A.; Adámek, Z. Effects of fish farming on macrophytes in temperate carp ponds. Aquac. Int. 2019, 27, 413–436. [Google Scholar] [CrossRef]
- Jílek, B. K fytocenologii rybničních společenstev. Preslia 1956, 28, 66–77. [Google Scholar]
- Gazda, J. Příspěvek k fytocenologii jihočeských rybníků. Sborn. Kraj. Vlastiv. Muz. České Budějovice Přír. Vědy 1958, 1, 64–76. [Google Scholar]
- Knižátková, E.; Lacina, D. Souhrny doporučených opatření pro ptačí oblasti. Ochr. Přír. 2010, 10, 12–14. [Google Scholar]
- Thiers, B.M. Index Herbariorum; The New York Botanical Garden: Bronx, NY, USA; Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 20 March 2021).
- Taylor, J.C.; Harding, W.R.; Archibald, C.G.M. A Methods Manual for Collection, Preparation and Analysis of Diatom Samples; Version 1.0; WRC Report TT 281/07; Water Research Commission: Gezina, South Africa, 2007; pp. 1–49. Available online: http://docs.niwa.co.nz/library/public/1770054839.pdf (accessed on 14 March 2016).
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae: Naviculaceae. In Süβwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Jena, Germany, 1986; 1. Teil; pp. 1–876. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae: Epithemiaceae, Surirellaceae. In Süβwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Jena, Germany, 1988; 2. Teil; pp. 1–876. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae: Centrales, Fragilariaceae, Eunotiaceae. In Süβwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Jena, Germany, 1991; 3. Teil; pp. 1–576. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae), Gomphonema. In Süβwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Jena, Germany, 1991; 4. Teil; pp. 1–437. [Google Scholar]
- Krammer, K. The Genus Pinnularia. In Diatoms of Europe; Lange-Bertalot, H., Ed.; A. R. G. Gantner Verlag K. G.: Rugell, Switzerland, 2000; Volume 1, pp. 1–703. [Google Scholar]
- Krammer, K. Cymbella. In Diatoms of Europe; Lange-Bertalot, H., Ed.; A. R. G. Gantner Verlag K. G.: Rugell, Switzerland, 2002; Volume 3, pp. 1–584. [Google Scholar]
- Krammer, K. Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella Supplements to cymbelloid taxa. In Diatoms of Europe; Lange-Bertalot, H., Ed.; A. R. G. Gantner Verlag K. G.: Rugell, Switzerland, 2003; Volume 4, pp. 1–530. [Google Scholar]
- Lange-Bertalot, H. Navicula sensu stricto, 10 genera separated from Navicula sensu stricto, Frustulia. In Diatoms of Europe; Lange-Bertalot, H., Ed.; A. R. G. Gantner Verlag K. G.: Rugell, Switzerland, 2001; Volume 2, pp. 1–526. [Google Scholar]
- Hofmann, G.; Werum, M.; Lange-Bertalot, H. Diatomeen im Süßwasserbenthos von Mitteleuropa. Bestimmungsflora Kieselalgen für die ökologische Praxis. Über 700 der häufigsten Arten und ihre Ökologie; Koeltz Scientific Books: Königstein, Germany, 2013; pp. 1–908. [Google Scholar]
- Lange-Bertalot, H.; Hofmann, G.; Werum, M.; Cantonati, M. Freshwater Benthic Diatoms of Central Europe: Over 800 Common Species Used in Ecological Assessment; English Edition with Updated Taxonomy and Added Species; Koeltz Botanical Books: Schmitten-Oberreifenberg, Germany, 2017; pp. 1–942. [Google Scholar]
- Fránková, M.; Poulíčková, A.; Neděla, V.; Tihlaříková, E.; Šumberová, K.; Letáková, M. The low temperature method for environmental scanning electron microscopy—A new method for observation of diatom assemblages in vivo. Diatom Res. 2019, 33, 397–403. [Google Scholar] [CrossRef]
- Kairesalo, T. Dynamics of epiphytic communities on Equisetum fluviatile L. In Periphyton of Freshwater Ecosystems; Wetzel, R.G., Ed.; Developments in Hydrobiology; Springer: Dordrecht, Germany, 1983; Volume 17, pp. 153–160. [Google Scholar]
- CHMI (Czech Hydrometeorological Institute). Historical Data—Meteorology and Climatology. Available online: https://www.chmi.cz/historicka-data/pocasi/uzemni-teploty (accessed on 10 March 2021).
- Pyšek, P.; Danihelka, J.; Sádlo, J.; Chrtek, J., Jr.; Chytrý, M.; Jarošík, V.; Kaplan, Z.; Krahulec, F.; Moravcová, L.; Pergl, J.; et al. Catalogue of alien plants of the Czech Republic (2nd edition): Checklist update, taxonomic diversity and invasion patterns. Preslia 2012, 84, 155–255. [Google Scholar]
- Grulich, V. Red List of vascular plants of the Czech Republic: 3rd edition. Preslia 2012, 84, 631–645. [Google Scholar]
- Grulich, V.; Chobot, K. (Eds.) Červený seznam cévnatých rostlin ČR [The Red List of vascular plants of the Czech Republic]. Příroda 2017, 35, 75–132. [Google Scholar]
- Wild, J.; Kaplan, Z.; Danihelka, J.; Petřík, P.; Chytrý, M.; Novotný, P.; Rohn, M.; Šulc, V.; Brůna, J.; Chobot, K.; et al. Plant distribution data for the Czech Republic integrated in the Pladias database. Preslia 2019, 91, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Danihelka, J.; Chrtek, J., Jr.; Kaplan, Z. Checklist of vascular plants of the Czech Republic. Preslia 2012, 84, 647–811. [Google Scholar]
- Caisová, L.; Gąbka, M. Charophytes (Characeae, Charophyta) in the Czech Republic: Taxonomy, autecology and distribution. Fottea 2009, 9, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R Package Version 2.5.-6. 2019. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 30 March 2021).
- Anderson, M.J. A new method for non-parametric multi-variate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 30 March 2021).
- Riato, L.; Van Ginkel, C.; Taylor, J.C. Zooplankton and diatoms of temporary and permanent freshwater pans in the Mpumalanga Highveld region, South Africa. Afr. Zool. 2014, 49, 113–127. [Google Scholar] [CrossRef]
- Popiela, A.; Łysko, A.; Molnár, V.A.; Kącki, Z.; Lukács, B.A. Distribution, morphology and habitats of Elatine triandra (Elatinaceae) in Europe, with particular reference to the central part of the continent. Acta Bot. Gallica 2015, 162, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Riato, L.; Della Bella, V.; Leira, M.; Taylor, J.C.; Oberholster, P.J. A diatom functional-based approach to assess changing environmental conditions in temporary depressional wetlands. Ecol. Indic. 2017, 78, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Kúr, P.; Píšová, S.; Tremetsberger, K.; Dřevojan, P.; Kącki, Z.; Hroudová, Z.; Böckelmann, J.; Bernhardt, K.G.; Šumberová, K. Ecology and genetics of Cyperus fuscus in Central Europe—A model for ephemeral wetland plant research and conservation. Water 2021, 13, 1277. [Google Scholar] [CrossRef]
- Juříček, M. Flora and vegetation of fishponds in the Bohemian-Moravian Highlands. Acta Rer. Natur. 2012, 13, 33–51. [Google Scholar]
- Cantonati, M. Diatom communities of springs in the southern Alps. Diatom Res. 1998, 13, 201–220. [Google Scholar] [CrossRef]
- Cantonati, M.; Corradini, G.; Jüttner, I.; Cox, E.J. Diatom assemblages in high mountain streams of the Alps and the Himalaya. Nova Hedwigia 2001, 123, 37–61. [Google Scholar]
- Cantonati, M.; Gerecke, R.; Bertuzzi, E. Springs of the Alps, sensitive ecosystems to environmental change: From biodiversity assessment to long-term studies. Hydrobiologia 2006, 562, 59–96. [Google Scholar] [CrossRef]
- Pätzig, M.; Kalettka, T.; Glemnitz, M.; Berger, G. What governs macrophyte species richness in kettle hole types? A case study from Northeast Germany. Limnologica 2012, 42, 340–354. [Google Scholar] [CrossRef]
- Poulíčková, A.; Letáková, M.; Hašler, P.; Coxb, E.; Duchoslav, M. Species complexes within epiphytic diatoms and their relevance for the bioindication of trophic status. Sci. Total Environ. 2017, 599–600, 820–833. [Google Scholar] [CrossRef] [PubMed]
- Kopp, R. Hydrochemie Nejen Pro Rybáře; Mendelova Univerzita v Brně: Brno, Czech Republic, 2015. [Google Scholar]
- Björnerås, C.; Weyhenmeyer, G.A.; Evans, C.D.; Gessner, M.O.; Grossart, H.-P.; Kangur, K.; Kokorite, I.; Kortelainen, P.; Laudon, H.; Lehtoranta, J.; et al. Widespread increases in iron concentration in European and North American freshwaters. Global Biogeochem. Cycles 2017, 31, 1488–1500. [Google Scholar] [CrossRef] [Green Version]
- Lacoul, P.; Freedman, B. Environmental influences on aquatic plants in freshwater ecosystems. Environ. Rev. 2006, 14, 89–136. [Google Scholar] [CrossRef]
- Emson, D.; Sayer, C.D.; Bennion, H.; Patmore, I.R.; Rioual, P. Mission possible: Diatoms can be used to infer past duckweed (lemnoid Araceae) dominance in ponds. J. Paleolimnol. 2018, 60, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, Z.; Danihelka, J.; Štěpánková, J.; Ekrt, L.; Chrtek, J., Jr.; Zázvorka, J.; Grulich, V.; Řepka, R.; Prančl, J.; Ducháček, M.; et al. Distributions of vascular plants in the Czech Republic. Part 2. Preslia 2016, 88, 229–322. [Google Scholar]
- Lovas-Kiss, Á.; Vizi, B.; Vincze, O.; Molnár, V.A.; Green, A.J. Endozoochory of aquatic ferns and angiosperms by mallards in Central Europe. J. Ecol. 2018, 106, 1714–1723. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Scola, S.; Angeli, N.; Guella, G.; Frassanito, R. Environmental controls of epilithic diatom depth-distribution in an oligotrophic lake characterized by marked water-level fluctuations. Eur. J. Phycol. 2009, 44, 15–29. [Google Scholar] [CrossRef]
- Carmignani, J.R.; Roy, A.H. Ecological impacts of winter water level drawdowns on lake littoral zones: A review. Aquat. Sci. 2017, 79, 803–824. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.H. The survival of freshwater algae during dry periods: Part I. An investigation of the algae of five small ponds. J. Ecol. 1958, 46, 149–167. [Google Scholar] [CrossRef]
- Hejný, S. Ökologische Charakteristik der Wasser und Sumpfpflanzen in den Slowakischen Tiefebenen; Verlag der Slowakischen Akademie der Wissenschaften: Bratislava, Czechoslovakia, 1960. [Google Scholar]
- Von Lampe, M. Wuchsform, Wuchsrhythmus und Verbreitung der Arten der Zwergbinsengesellschaften. Diss. Bot. 1996, 266, 1–353. [Google Scholar]
- Šumberová, K.; Lososová, Z.; Ducháček, M.; Horáková, V.; Fabšičová, M. Distribution, habitat ecology, soil seed bank and seed dispersal of threatened Lindernia procumbens and alien Lindernia dubia (Antirrhinaceae) in the Czech Republic. Phyton 2012, 52, 39–72. [Google Scholar]
- Leira, M.; Meijide-Failde, R.; Torres, E. Diatom communities in thermo-mineral springs of Galicia (NW Spain). Diatom Res. 2017, 32, 29–42. [Google Scholar] [CrossRef]
- Ponader, K.C.; Potapova, M.G. Diatoms from the genus Achnanthidium in flowing waters of the Appalachian Mountains (North America): Ecology, distribution and taxonomic notes. Limnologica 2007, 37, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Souffreau, C.; Vanormelingen, P.; Verleyen, E.; Sabbe, K.; Vyverman, W. Tolerance of benthic diatoms from temperate aquatic and terrestrial habitats to experimental desiccation and temperature stress. Phycologia 2010, 49, 309–324. [Google Scholar] [CrossRef] [Green Version]
- Vasiljević, B.; Simić, S.B.; Paunović, M.; Zuliani, T.; Krizmanić, J.; Marković, V.; Tomović, J. Contribution to the improvement of diatom-based assessments of the ecological status of large rivers–The Sava River Case Study. Sci. Total Environ. 2017, 605, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Falasco, E.; Piano, E.; Bona, F. Diatom flora in Mediterranean streams: Flow intermittency threatens endangered species. Biodivers. Conserv. 2016, 25, 2965–2986. [Google Scholar] [CrossRef]
- Ács, É.; Bíró, T.; Berta, C.; Duleba, M.; Földi, A.; Grigorszky, I.; Hidas, A.; Knisz, J.; Korponai, J.L.; Trábert, Z.; et al. Long-term changes of species composition and functional traits of epiphytic diatoms in the Szigetköz region (Hungary) of the Danube River. Water 2020, 12, 776. [Google Scholar] [CrossRef] [Green Version]
- Patrick, R. The effects of increasing light and temperature on the structure of diatom communities. Limnol. Oceanogr. 1971, 16, 405–421. [Google Scholar] [CrossRef]
- Carrick, H.J.; Lowe, R.L. Benthic algal response to N and P enrichment along a pH gradient. Hydrobiologia 1989, 179, 119–127. [Google Scholar] [CrossRef]
- Kaczmarek, H.; Mazaeva, O.A.; Kozyreva, E.A.; Babicheva, V.A.; Tyszkowski, S.; Rybchenko, A.A.; Brykała, D.; Bartczak, A.; Słowiński, M. Impact of large water level fluctuations on geomorphological processes and their interactions in the shore zone of a dam reservoir. J. Great Lakes Res. 2016, 42, 926–941. [Google Scholar] [CrossRef]
- Van Zuidam, B.G.; Peeters, E.T.H.M. Wave forces limit the establishment of submerged macrophytes in large shallow lakes. Limnol. Oceanogr. 2015, 60, 1536–1549. [Google Scholar] [CrossRef]
- Bertrin, V.; Boutry, S.; Alard, D.; Haury, J.; Jan, G.; Moreira, S.; Ribaudo, C. Prediction of macrophyte distribution: The role of natural versus anthropogenic physical disturbances. Appl. Veg. Sci. 2018, 21, 395–410. [Google Scholar] [CrossRef]
- Kaplan, Z.; Danihelka, J.; Chrtek, J., Jr.; Zázvorka, J.; Koutecký, P.; Ekrt, L.; Řepka, R.; Štěpánková, J.; Jelínek, B.; Grulich, V.; et al. Distributions of vascular plants in the Czech Republic. Part 8. Preslia 2019, 91, 257–368. [Google Scholar] [CrossRef]
- Gaberščik, A.; Grašič, M.; Abram, D.; Zelnik, I. Water level fluctuations and air temperatures affect common reed habitus and productivity in an intermittent wetland ecosystem. Water 2020, 12, 2806. [Google Scholar] [CrossRef]
- Hillman, W.S. The Lemnaceae, or Duckweeds: A Review of the Descriptive and Experimental Literature. Bot. Rev. 1961, 27, 221–287. [Google Scholar] [CrossRef]
- Kolář, F.; Šumberová, K.; Ducháček, M. Cerastium dubium—Rožec pochybný. In Nálezy Zajímavých a Nových Druhů v Květeně Jižní Části Čech XXI; Lepší, M., Lepší, P., Eds.; Jihočeské Muzeum v Českých Budějovicích: České Budějovice, Czech Republic, 2015; Volume 55, pp. 16–17. [Google Scholar]
- Poulíčková, A.; Neustupa, J.; Špačková, J.; Škaloud, P. Distribution of epipelic diatoms in artificial fishponds along environmental and spatial gradients. Hydrobiologia 2009, 624, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, Z.; Danihelka, J.; Šumberová, K.; Chrtek, J., Jr.; Rotreklová, O.; Ekrt, L.; Štěpánková, J.; Taraška, V.; Trávníček, B.; Prančl, J.; et al. Distributions of vascular plants in the Czech Republic. Part 5. Preslia 2017, 89, 333–439. [Google Scholar] [CrossRef]
- Feuchtmayr, H.; Moran, R.; Hatton, K.; Connor, L.; Heyes, T.; Moss, B.; Harvey, I.; Atkinson, D. Global warming and eutrophication: Effects on water chemistry and autotrophic communities in experimental hypertrophic shallow lake mesocosms. J. Appl. Ecol. 2009, 46, 713–723. [Google Scholar] [CrossRef]
- Lu, J.; Bunn, S.E.; Burford, M.A. Nutrient release and uptake by littoral macrophytes during water level fluctuations. Sci. Total Environ. 2018, 622, 29–40. [Google Scholar] [CrossRef]
- Blindow, I. The composition and density of epiphyton on several species of submerged macrophytes—The neutral hypothesis tested. Aquat. Bot. 1987, 29, 157–168. [Google Scholar] [CrossRef]
- Buczkó, K. The occurrence of the epiphytic diatom Lemnicola hungarica on different European Lemnaceae species. Fottea 2007, 7, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Riato, L.; Leira, M. Heterogeneity of epiphytic diatoms in shallow lakes: Implications for lake monitoring. Ecol. Indic. 2020, 111, 105988. [Google Scholar] [CrossRef]
- Totti, C.; Poulin, M.; Romagnoli, T.; Perrone, C.; Pennesi, C.; De Stefano, M. Epiphytic diatom communities on intertidal seaweeds from Iceland. Polar Biol. 2009, 32, 1681–1691. [Google Scholar] [CrossRef]


















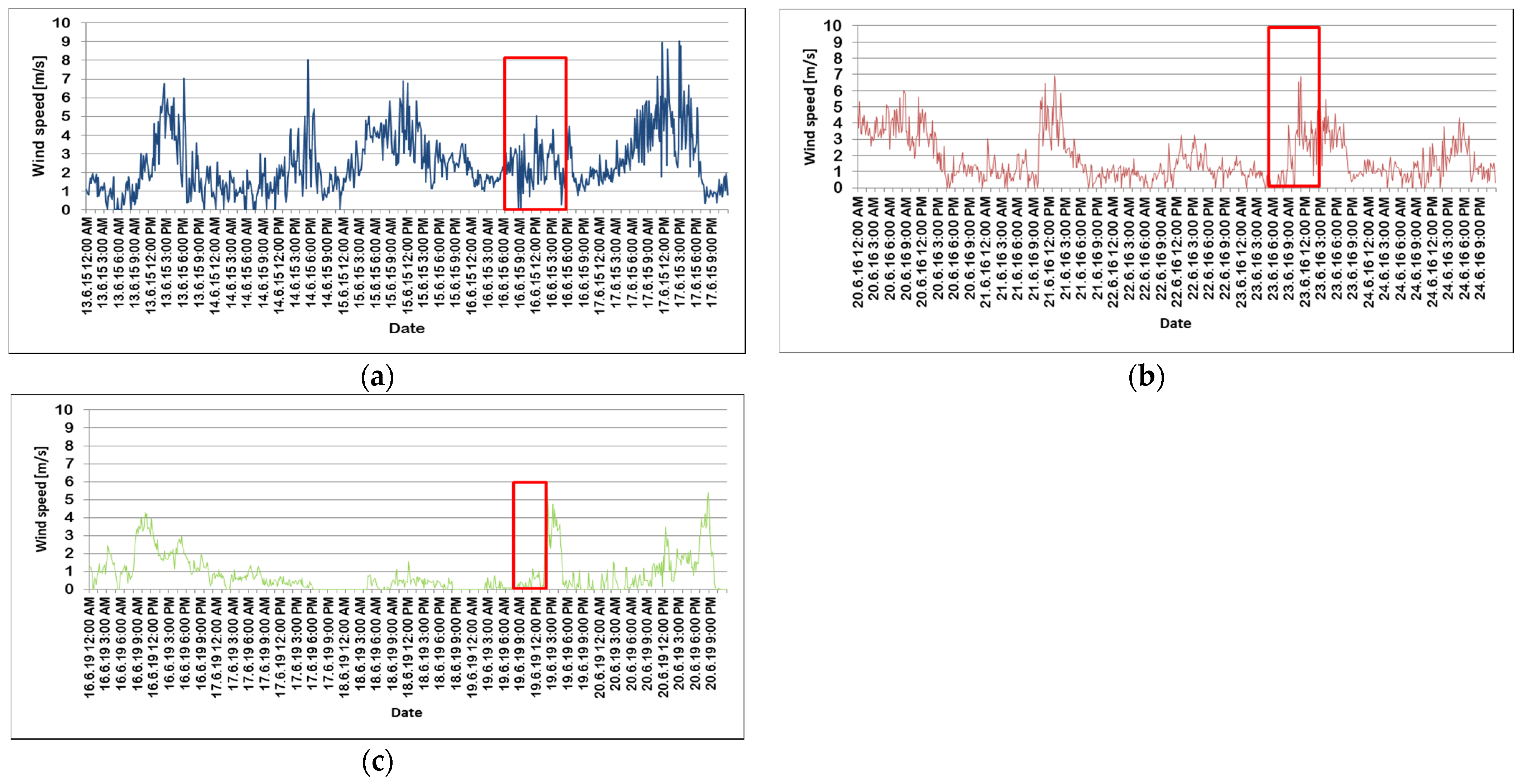
| Wind Events | 2014 (12.5–31.8) | 2015 (1.4.–31.8.) | 2016 (23.5.–31.8.) | 2019 (1.4–31.8.) |
|---|---|---|---|---|
| no wind (<0.2 m/s) | 548 (3.42%) | 1602 (7.3%) | 749 (5.17%) | 8415 (38.20%) |
| 5.5–7.9 m/s | 892 (5.56%) | 1516 (6.91%) | 548 (3.78%) | 80 (0.36%) |
| 8.0–13.9 m/s | 190 (1.18%) | 395 (1.80%) | 120 (0.83%) | 2 (0.01%) |
| >13.9 m/s | 3 (0.02%) | 27 (0.12%) | 3 (0.02%) | 0 |
| rest (>0.02 and <5.5 m/s, %) | 89.82 | 81.87 | 90.20 | 61.43 |
| mean (m/s) | 2.36 | 2.50 | 1.99 | 0.95 |
| max. (m/s) | 16.64 | 19.18 | 17.39 | 11.57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šumberová, K.; Vild, O.; Ducháček, M.; Fabšičová, M.; Potužák, J.; Fránková, M. Drivers of Macrophyte and Diatom Diversity in a Shallow Hypertrophic Lake. Water 2021, 13, 1569. https://doi.org/10.3390/w13111569
Šumberová K, Vild O, Ducháček M, Fabšičová M, Potužák J, Fránková M. Drivers of Macrophyte and Diatom Diversity in a Shallow Hypertrophic Lake. Water. 2021; 13(11):1569. https://doi.org/10.3390/w13111569
Chicago/Turabian StyleŠumberová, Kateřina, Ondřej Vild, Michal Ducháček, Martina Fabšičová, Jan Potužák, and Markéta Fránková. 2021. "Drivers of Macrophyte and Diatom Diversity in a Shallow Hypertrophic Lake" Water 13, no. 11: 1569. https://doi.org/10.3390/w13111569
APA StyleŠumberová, K., Vild, O., Ducháček, M., Fabšičová, M., Potužák, J., & Fránková, M. (2021). Drivers of Macrophyte and Diatom Diversity in a Shallow Hypertrophic Lake. Water, 13(11), 1569. https://doi.org/10.3390/w13111569