
Ubiquity of Euglena mutabilis Population in Three Ecologically Distinct Acidic Habitats in Southwestern Japan


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Determination of Environmental Variables
2.3. Collection and Isolation of the Euglena Population
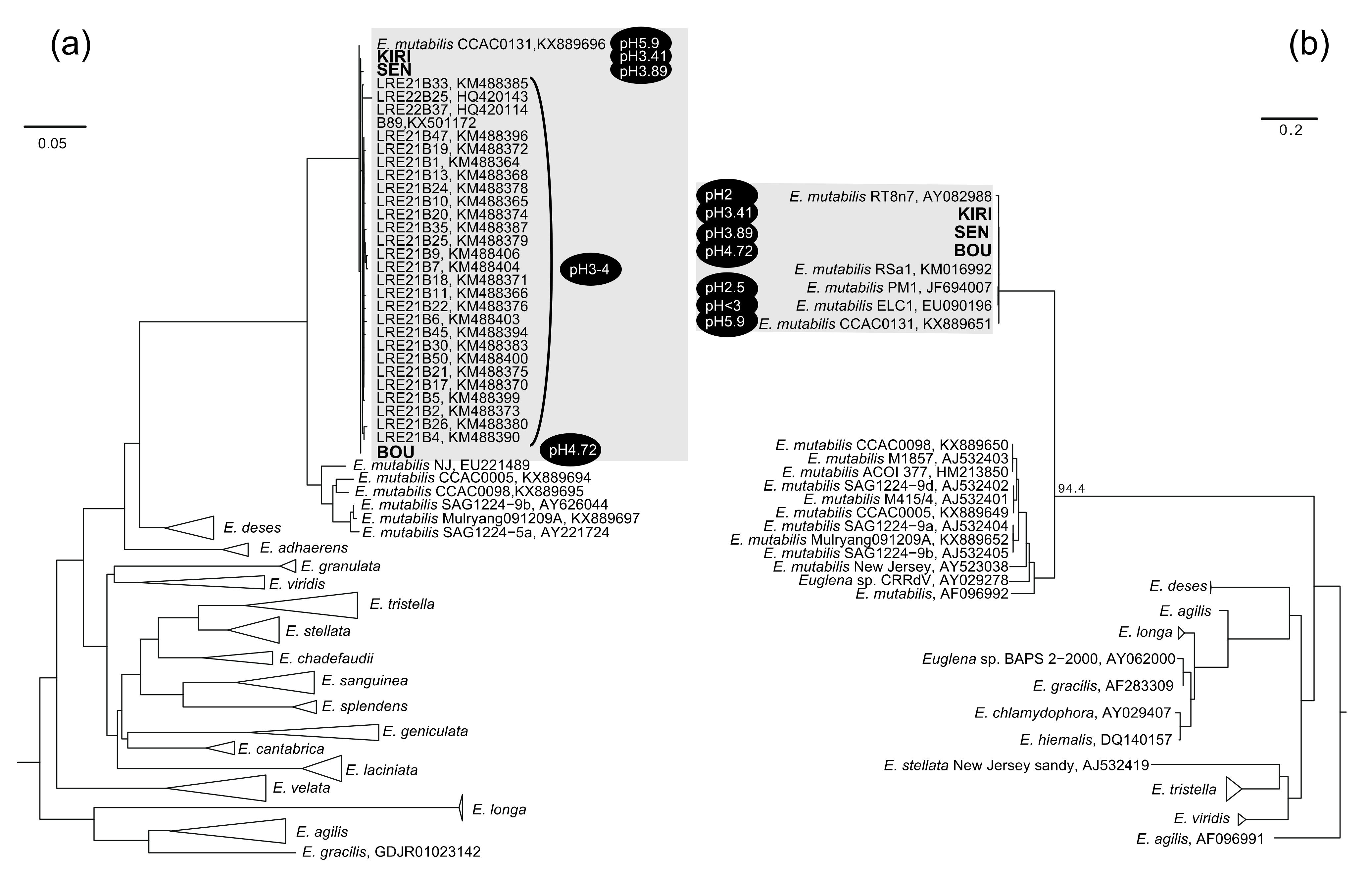
2.4. Phylogenetic Analysis of SSU rRNA Genes
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rijstenbil, J.W.; Merks, A.G.A.; Peene, J.; Poortvliet, T.C.W.; Wijnholds, J.A. Phytoplankton composition and spatial distribution of copper and zinc in the Fal Estuary (Cornwall, UK). Hydrobiol. Bull. 1991, 25, 37–43. [Google Scholar] [CrossRef]
- Brake, S.S.; Dannelly, H.K.; Connors, K.A.; Hasiotis, S.T. Influence of water chemistry on the distribution of an acidophilic protozoan in an acid mine drainage system at the abandoned Green Valley coal mine, Indiana, USA. Appl. Geochem. 2001, 16, 1641–1652. [Google Scholar] [CrossRef]
- Gross, W.; Oesterhelt, C.; Tischendorf, G.; Lederer, F. Characterization of a non-thermophilic strain of the red algal genus Galdieria isolated from Soos (Czech Republic). Eur. J. Phycol. 2002, 37, 477–482. [Google Scholar] [CrossRef] [Green Version]
- Pentecost, A. The distribution of Euglena mutabilis in Sphagna, with reference to the Malham Tarn Noethern Fen. Field Stud. 1982, 5, 591–606. [Google Scholar]
- Searles, P.S.; Kropp, B.R.; Flint, S.D.; Caldwell, M.M. Influence of solar UV-B radiation on peatland microbial communities of southern Argentinia. New Phytol. 2001, 152, 213–221. [Google Scholar] [CrossRef]
- Casiot, C.; Bruneel, O.; Personné, J.C.; Leblanc, M.; Elbaz-Poulichet, F. Arsenic oxidation and bioaccumulation by the acidophilic protozoan, Euglena mutabilis, in acid mine drainage (Carnoulès, France). Sci. Total Environ. 2004, 3202, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, J.W.; Lloyd, E.J.H.; Whitton, B.A. Chemistry and vegetation of highly acidic streams. Freshw. Biol. 1975, 5, 563–576. [Google Scholar] [CrossRef]
- Sabater, S.; Buchaca, T.; Cambra, J.; Catalan, J.; Guasch, H.; Ivorra, N.; Muñoz, I.; Navarro, E.; Real, M.; Romaní, A. Structure and function of benthic algal communities in an extremely acid river. J. Phycol. 2003, 39, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Bray, J.P.; Broady, P.A.; Niyogi, D.K.; Harding, J.S. Periphyton communities in New Zealand streams impacted by acid mine drainage. Mar. Freshw. Res. 2008, 59, 1084–1091. [Google Scholar] [CrossRef]
- Zheng, J.; Haraguchi, A. Inorganic nitrogen source for autotrophic growth of Euglena mutabilis Schmitz. Phycol. Res. 2018, 66, 155–158. [Google Scholar] [CrossRef]
- Negoro, K. On a Euglena sp. from acidic mineral water in Japan. J. Plant Res. (Bot. Mag. Tokyo) 1943, 57, 132–136, (in Japanese with German summary). [Google Scholar]
- Cramer, M.; Myers, J.C. Growth and photosynthetic characteristics of Euglena gracilis. Arch. Microbiol. 1952, 17, 384–402. [Google Scholar] [CrossRef]
- Hädder, D.P.; Melkonian, M. Phototaxis in the gliding flagellate, Euglena mutabilis. Arch. Microbiol. 1983, 135, 25–29. [Google Scholar] [CrossRef]
- Surek, B.; Melkonian, M. A cryptic cytostome is present in Euglena. Protoplasma 1986, 133, 39–49. [Google Scholar] [CrossRef]
- Wołowski, K.; Turnau, K.; Henriques, F.S. The algal flora of an extremely acidic, metal-rich drainage pond of São Domingos pyrite mine (Portugal). Cryptogam. Algol. 2008, 29, 313–324. [Google Scholar]
- DeLong, E.F. Archaea in coastal marine environments. Proc. Natl. Acad. Sci. USA 1992, 89, 5685–5689. [Google Scholar] [CrossRef] [Green Version]
- Sittenfeld, A.; Mora, M.; Ortega, J.M.; Albertazzi, F.; Cordero, A.; Roncel, M.; Sánchez, E.; Vargas, M.; Fernández, M.; Weckesser, J.; et al. Characterization of a photosynthetic Euglena strain isolated from an acidic hot mud pool of a volcanic area of Costa Rica. FEMS Microbiol. Ecol. 2002, 42, 151–161. [Google Scholar] [CrossRef]
- Yanagawa, K.; Shiraishi, F.; Tanigawa, Y.; Maeda, T.; Mustapha, N.A.; Owari, S.; Tomaru, H.; Matsumoto, R.; Kano, A. Endolithic microbial habitats hosted in carbonate nodules currently forming within sediment at a high methane flux site in the Sea of Japan. Geosciences 2019, 9, 463. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Kim, J.I.; Linton, E.W.; Shin, W. Morphological and genetic diversity of Euglena deses group (Euglenophyceae) with emphasis on cryptic species. Algae 2016, 31, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Beamud, S.G.; Diaz, M.M.; Pedrozo, F.L. Nutrient limitation of phytoplankton in a naturally acidic lake (Lake Caviahue, Argentina). Limnology 2010, 11, 103–113. [Google Scholar] [CrossRef]
- Zettler, L.A.A.; Gómez, F.; Zettler, E.; Keenan, B.G.; Amils, R.; Sogin, M.L. Eukaryotic diversity in Spain’s River of Fire. Nature 2002, 417, 137. [Google Scholar] [CrossRef] [PubMed]
- Kloos, K.; Hüsgen, U.M.; Bothe, H. DNA-probing for genus coding for denitrification, N2-fixation and nitrification in bacteria isolated from different soils. Z. Nat. 1998, 53c, 69–81. [Google Scholar]
- Ňancucheo, I.; Johnson, D.B. Acidophilic algae isolated from mine-impacted environments and their roles in sustaining heterotrophic acidophiles. Front. Microbiol. 2012, 3, 325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grettenberger, C.L.; Pearce, A.R.; Bibby, K.J.; Jones, D.S.; Burgos, W.D.; Macalady, J.L. Efficient low-pH iron removal by a microbial iron oxide mound ecosystem at Scalp Level Run. Appl. Environ. Microbiol. 2017, 83, e00015-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, D.S.; Kohl, C.; Grettenberger, C.; Larson, L.N.; Burgos, W.D.; Macalady, J.L. Geochemical niches of iron-oxidizing acidophiles in acidic coal mine drainage. Appl. Environ. Microbiol. 2015, 81, 1242–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Location | Sensui | Bougatsuru | Ebinokogen | |
| coal mine drainage | cold spring | mire pool | ||
| 33.78264° N | 33.09602° N | 31.94864 | ||
| 130.65363° E | 131.25940° E | 130.85560 | ||
| 16 m a.s.l. | 1238 m a.s.l. | 1243 m a.s.l. | ||
| (26 April 2019) | (28 November 2019) | (17 May 2019) | ||
| Water temperature | °C | 22.5 | 10.2 | 16.2 |
| pH | 4.10 | 4.78 | 3.41 | |
| EC | mS/m | 185 | 24.5 | 81.7 |
| Cl− | mg/L | 38.1 | 14.3 | 57.0 |
| NO3− | mg/L | 1.2 | 1.2 | not determined |
| PO43− | mg/L | 0.0 | 0.0 | 0.0 |
| SO42− | mg/L | 1375.0 | 170.1 | 210.0 |
| Na+ | mg/L | 31.2 | 11.8 | 24.1 |
| NH4+ | mg/L | 2.9 | 1.8 | 0.0 |
| K+ | mg/L | 3.0 | 1.5 | 5.4 |
| Mg2+ | mg/L | 11.3 | 6.8 | 12.3 |
| Ca2+ | mg/L | 131.3 | 19.5 | 32.8 |
| Al | mg/L | 20.9 | 0.3 | 9.86 |
| As | mg/L | 0.2 | 0.1 | 1.5 * |
| Fe | mg/L | 56.2 | 2.8 | 25.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yanagawa, K.; Haraguchi, A.; Yoshitake, K.; Asamatsu, K.; Harano, M.; Yamashita, K.; Ishibashi, J.-i. Ubiquity of Euglena mutabilis Population in Three Ecologically Distinct Acidic Habitats in Southwestern Japan. Water 2021, 13, 1570. https://doi.org/10.3390/w13111570
Yanagawa K, Haraguchi A, Yoshitake K, Asamatsu K, Harano M, Yamashita K, Ishibashi J-i. Ubiquity of Euglena mutabilis Population in Three Ecologically Distinct Acidic Habitats in Southwestern Japan. Water. 2021; 13(11):1570. https://doi.org/10.3390/w13111570
Chicago/Turabian StyleYanagawa, Katsunori, Akira Haraguchi, Kai Yoshitake, Katsuhiro Asamatsu, Masanari Harano, Kei Yamashita, and Jun-ichiro Ishibashi. 2021. "Ubiquity of Euglena mutabilis Population in Three Ecologically Distinct Acidic Habitats in Southwestern Japan" Water 13, no. 11: 1570. https://doi.org/10.3390/w13111570
APA StyleYanagawa, K., Haraguchi, A., Yoshitake, K., Asamatsu, K., Harano, M., Yamashita, K., & Ishibashi, J.-i. (2021). Ubiquity of Euglena mutabilis Population in Three Ecologically Distinct Acidic Habitats in Southwestern Japan. Water, 13(11), 1570. https://doi.org/10.3390/w13111570