
Temperature Dependence of Freshwater Phytoplankton Growth Rates and Zooplankton Grazing Rates



Abstract
:1. Introduction
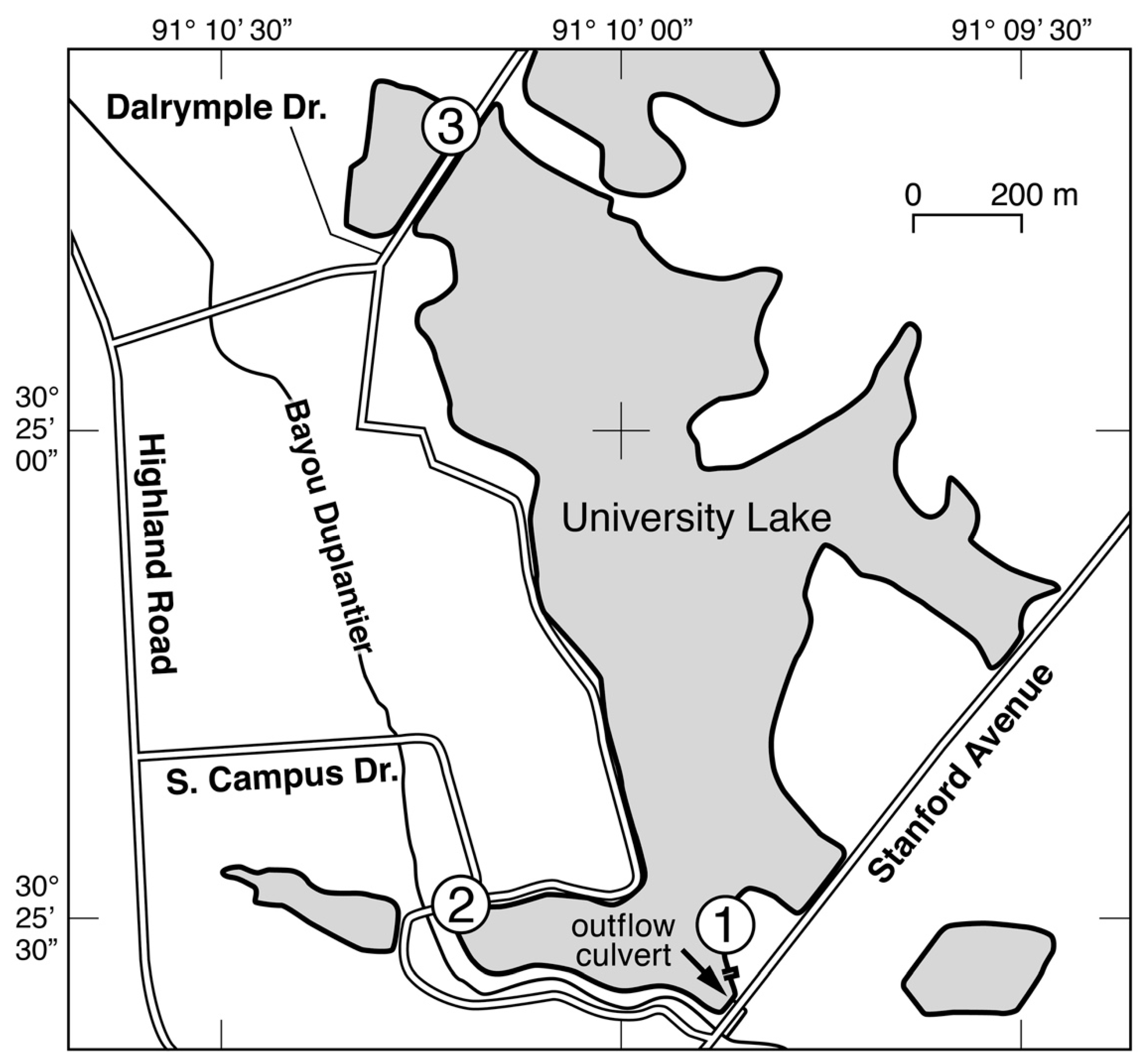
2. Materials and Methods
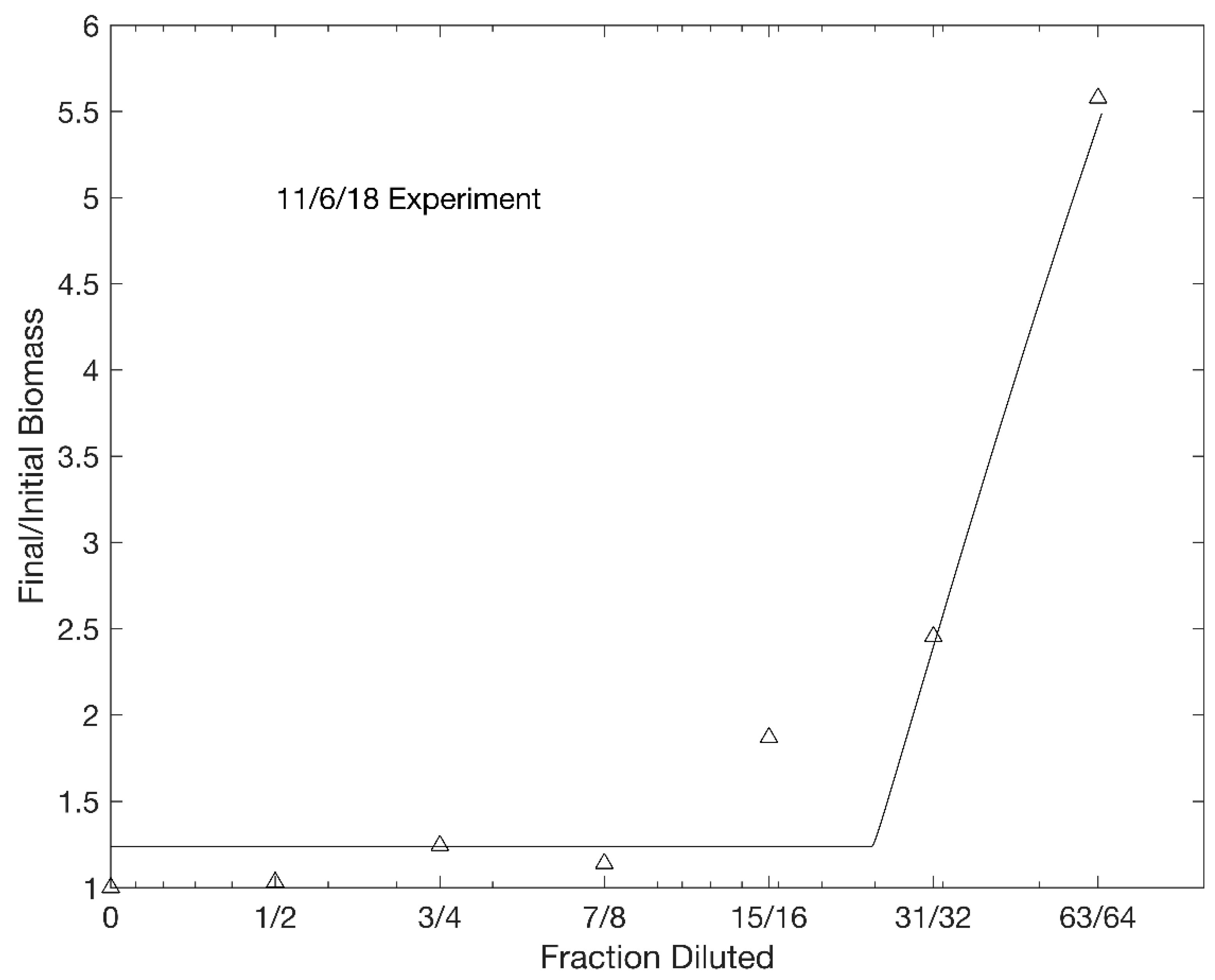
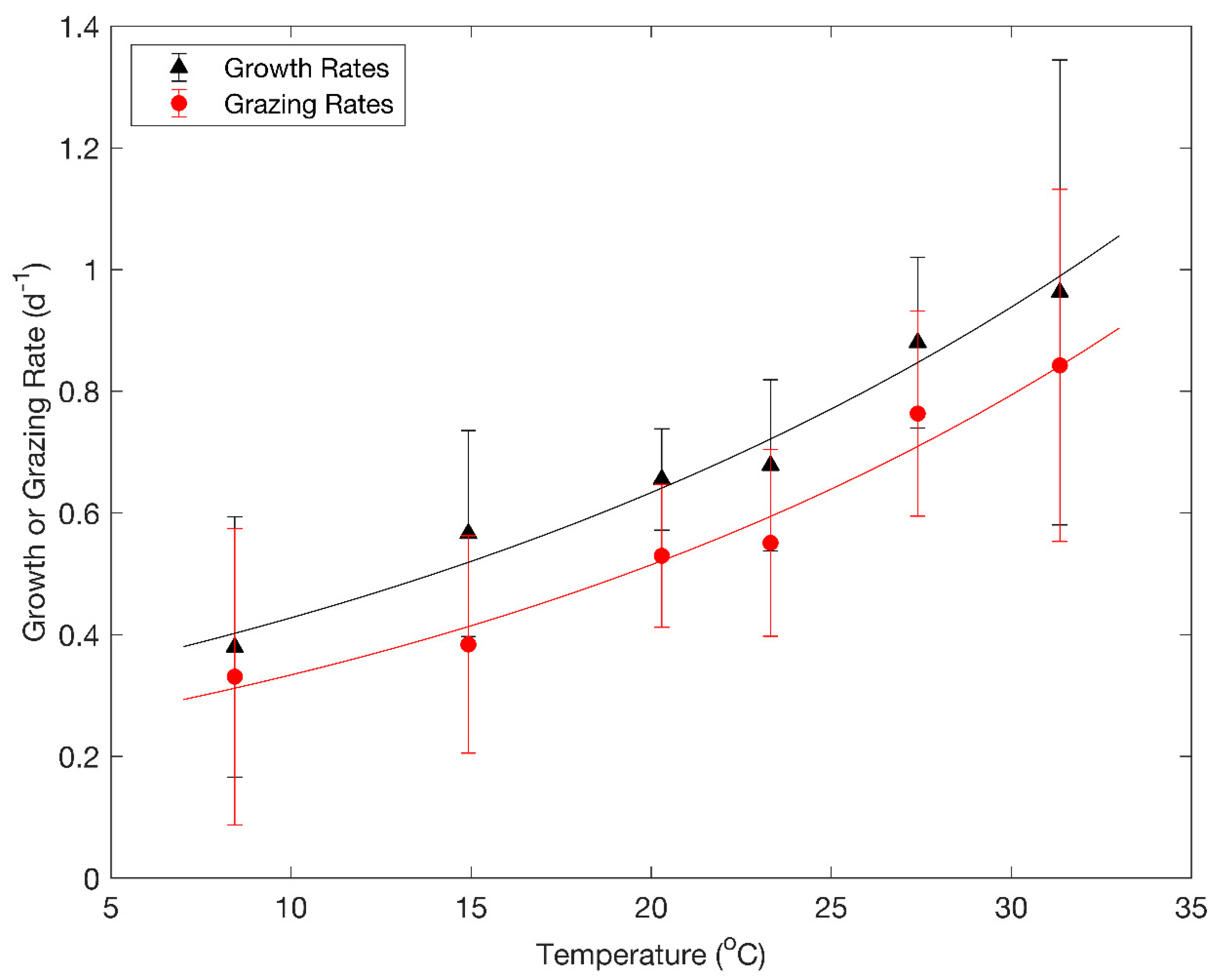
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Date | Initial Chlorophyll (mg L−1) | Temp. (°C) | μ (d−1) 1 | g (d−1) 2 | Gmax (mg chl a L−1 d−1) 3 | K (mg chl a L−1) 4 | R2 5 |
|---|---|---|---|---|---|---|---|
| 10/15/18 | 372 | 26.2 | 0.95 | 1.10 | 408 | 21 | 0.895 |
| 10/26/18 | 156 | 20.3 | 2.1 | 2.00 | 312 | 8 | 0.876 |
| 11/6/18 | 148 | 22.2 | 2.65 | 2.59 | 384 | 3 | 0.967 |
| 11/12/18 | 137 | 16.8 | 0.75 | 0.70 | 96 | 1.5 | 0.839 |
| 11/18/18 | 96.7 | 11 | 1.5 | 1.49 | 144 | 1 | 0.915 |
| 12/3/18 | 87 | 14 | 0.2 | 0.28 | 24 | 5 | 0.722 |
| 1/17/19 | 65.5 | 12.8 | 2.6 | 2.56 | 168 | 4.5 | 0.958 |
| 2/22/19 | 40.1 | 20.3 | 0.6 | 0.60 | 24 | 1 | 0.946 |
| 4/26/19 | 2012 | 23.6 | 0.35 | 0.62 | 1248 | 400 | 0.834 |
| 6/27/19 | 232 | 30.5 | 0.35 | 0.31 | 72 | 102 | 0.858 |
| 9/10/19 | 376 | 33 | 0.3 | 0.38 | 144 | 67 | 0.966 |
| 9/19/19 | 337 | 30.3 | 1.35 | 1.42 | 480 | 69.5 | 0.999 |
| 9/22/19 | 114 | 31.6 | 1.85 | 1.26 | 144 | 9.5 | 0.989 |
| 11/12/19 | 177 | 11 | 0.4 | 0.54 | 96 | 30.5 | 1.0 |
| 2/26/20 | 56.1 | 15.7 | 0.4 | 0.43 | 24 | 8.5 | 0.63 |
| 3/16/20 | 60.7 | 24.2 | 0.2 | 0.40 | 24 | 4.5 | 0.86 |
| Temperature (°C) | Phytoplankton Growth Rate (d−1) | Zooplankton Grazing Rate (d−1) | Reference |
|---|---|---|---|
| 22 | 1.23 | 1.79 | [18] |
| 9 | 1.28 | 1.62 | [18] |
| 11 | 0.7 | 0.1 | [15] |
| 11 | 0.4 | −0.3 | [15] |
| 10 | 0.9 | 0.2 | [15] |
| 19 | 1.2 | 0.3 | [15] |
| 16 | 0 | −0.1 | [15] |
| 15 | −1.3 | −1.5 | [15] |
| 16 | −0.7 | −1.3 | [15] |
| 21 | −0.2 | −1.1 | [15] |
| 20 | 0.8 | 0.4 | [15] |
| 22 | 0.9 | 0 | [15] |
| 25 | 0.4 | 0.3 | [15] |
| 22 | 1 | 0.5 | [15] |
| 20 | 1.1 | 0.8 | [15] |
| 16 | 0.9 | 0.6 | [15] |
| 13 | 0.9 | 0.7 | [15] |
| 6 | −0.8 | −0.9 | [15] |
| 18.6 | 1.33 | −1.83 | [19] |
| 18.6 | 0.19 | −0.35 | [19] |
| 21 | −0.1 | 0.03 | [16] |
| 21 | 0 | 0 | [16] |
| 17 | −0.05 | 0.3 | [16] |
| 21 | 0.34 | 0.07 | [16] |
| 21 | 0.96 | 1.34 | [16] |
| 17 | 0.87 | 0.51 | [16] |
| 21 | 0.26 | 0.21 | [16] |
| 21 | 0.1 | 0.17 | [16] |
| 17 | 1.42 | 0.27 | [16] |
| 21 | 0.43 | 0.33 | [16] |
| 21 | 0.16 | 0.4 | [16] |
| 17 | 0.6 | 0 | [16] |
| 21 | 0.32 | 0 | [16] |
| 21 | 0.16 | 0.11 | [16] |
| 17 | 0.91 | 0.27 | [16] |
| 21 | 0.5 | 0.25 | [16] |
| 21 | 0.18 | 0.1 | [16] |
| 17 | 1.34 | 0.69 | [16] |
| 21 | 0.51 | 0.53 | [16] |
| 21 | 0.32 | 0.45 | [16] |
| 17 | 0.29 | 1.13 | [16] |
| 19.2 | 0.19 | 0.32 | [20] |
| 19.2 | 1.87 | 2.79 | [20] |
| 19.2 | 1.51 | 1.65 | [20] |
| 19.2 | 1.7 | 1.01 | [20] |
| 15 | 0.09 | 0.07 | [21] |
| 20 | 0.37 | 0.26 | [21] |
| 20.2 | 0.78 | 0.87 | [21] |
| 15 | 0.24 | 1.01 | [21] |
| 20 | 1.22 | 1.34 | [21] |
| 20.2 | 1.66 | 1.92 | [21] |
| 20.7 | 0.38 | 0.27 | [17] |
| 18.6 | 0.73 | 0.62 | [17] |
| 22.1 | 1.15 | 0.66 | [17] |
| 21.4 | 0.69 | 0.42 | [17] |
| 23.2 | 0.43 | 0.21 | [17] |
| 24 | 0.37 | 0.28 | [17] |
| 21.3 | 0.52 | 0.42 | [17] |
| 19.6 | 0.23 | 0.16 | [17] |
| 20.7 | 0.5 | 0.04 | [17] |
| 18.6 | 0.34 | 0.06 | [17] |
| 22.1 | 0.19 | 0.02 | [17] |
| 21.4 | 0.3 | 0.04 | [17] |
| 23.2 | 0.25 | 0.059 | [17] |
| 24 | 0.18 | 0.07 | [17] |
| 21.3 | 0.44 | 0.04 | [17] |
| 19.6 | 0.23 | 0.03 | [17] |
| 18 | 0.58 | 0.52 | [22] |
| 18 | 2.32 | 2.29 | [22] |
| 21 | 1.07 | 1.93 | [23] |
| 21 | 0.74 | 0.88 | [23] |
| 21 | 0.33 | 0.28 | [23] |
| 21 | 0.5 | 1.6 | [23] |
| 21 | 0.31 | 0.67 | [23] |
| 21 | 0.68 | 0.18 | [23] |
| 8.5 | 0.312 | 0.264 | [24] |
| 8.5 | 0.264 | 0.144 | [24] |
| 8.5 | 0.144 | 0.264 | [24] |
| 8.5 | 0.48 | 0.528 | [24] |
| 8.5 | 0.456 | 0.528 | [24] |
| 28 | 0.61 | 0.58 | [26] |
| 28 | 1.08 | 0.61 | [26] |
| 24 | 0.25 | 0.37 | [25] |
| 24 | 0.47 | 0.68 | [25] |
| 24 | 0.8 | 0.3 | [25] |
| 24 | 0.74 | 0.69 | [25] |
| 24 | 0.65 | 0.38 | [25] |
References
- Eppley, R.W. Temperature and phytoplankton growth in the sea. Fish. Bull. 1972, 70, 1063–1085. [Google Scholar]
- Goldman, J.C.; Carpenter, E.J. Kinetic approach to effect of temperature on algal growth. Limnol. Oceanogr. 1974, 19, 756–766. [Google Scholar] [CrossRef]
- Bissinger, J.E.; Montagnes, D.J.S.; Harples, J.; Atkinson, D. Predicting marine phytoplankton maximum growth rates from temperature: Improving on the Eppley curve using quantile regression. Limnol. Oceanogr. 2008, 53, 487–493. [Google Scholar] [CrossRef]
- Kremer, C.T.; Thomas, M.K.; Litchman, E. Temperature- and size-scaling of phytoplankton population growth rates: Reconciling the Eppley curve and the metabolic theory of ecology. Limnol. Oceanogr. 2017, 62, 1658–1670. [Google Scholar] [CrossRef]
- Norris, B.; Laws, E.A. Nutrients and Phytoplankton in a Shallow, Hypereutrophic Urban Lake: Prospects for Restoration. Water 2017, 9, 11. [Google Scholar]
- Landry, M.R.; Hassett, R.P. Estimating the grazing impact of marine micro-zooplankton. Mar. Biol. 1982, 67, 283–288. [Google Scholar] [CrossRef]
- Evans, G.T.; Paranjape, M.A. Precision of estimates of phytoplankton growth and microzooplankton grazing when the functional-response of grazers may be nonlinear. Mar. Ecol. Prog. Ser. 1992, 80, 285–290. [Google Scholar] [CrossRef]
- Gallegos, C.L. Microzooplankton grazing on phytoplankton in the Rhode River, Maryland - nonlinear feeding kinetics. Mar. Ecol. Prog. Ser. 1989, 57, 23–33. [Google Scholar] [CrossRef]
- Moigis, A.G. The clearance rate of microzooplankton as the key element for describing estimated non-linear dilution plots demonstrated by a model. Mar. Biol. 2006, 149, 743–762. [Google Scholar] [CrossRef]
- Holling, C.S. The components of predation as revealed by a study of small-mammal predation of the European pine sawfly. Can. Entomol. 1959, 91, 293–320. [Google Scholar] [CrossRef]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Chen, B.; Laws, E.A.; Liu, H.; Huang, B. Estimating microzooplankton grazing half-saturatiion constants from dilution experiments with nonlinear feeding kinetics. Limnol. Oceanogr. 2014, 59, 639–644. [Google Scholar] [CrossRef] [Green Version]
- Atlas, D.; Bannister, T.T. Dependence of mean spectral extinction coefficient of phytoplankton on depth, water color, and species. Limnol. Oceanogr. 1980, 25, 157–159. [Google Scholar] [CrossRef]
- Holm-Hansen, O.; Riemann, B. Chlorophyll a determination: Improvements in methodology. Oikos 1978, 30, 438–447. [Google Scholar] [CrossRef]
- Boyer, J.; Rollwagen-Bollens, G.; Bollens, S.M. Microzooplankton grazing before, during and after a cyanobacterial bloom in Vancouver Lake, Washington, USA. Aquat. Microb. Ecol. 2011, 64, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Lavrentyev, P.J.; Vanderploeg, H.A.; Franzé, G.; Chacin, D.H.; Liebig, J.R.; Johengen, T.H. Microzooplankton distribution, dynamics, and trophic interactions relative to phytoplankton and quagga mussels in Saginaw Bay, Lake Huron. J.Great Lakes Res. 2014, 40, 95–105. [Google Scholar] [CrossRef]
- Tadonleke, R.D.; Sime-Ngando, T. Rates of growth and microbial grazing mortality of phytoplankton in a recent artificial lake. Aquat. Microb. Ecol. 2000, 22, 301–313. [Google Scholar] [CrossRef]
- Adrian, R.; Wickham, S.A.; Butler, N.M. Trophic interactions between zooplankton and the microbial community in contrasting food webs: The epilimnion and deep chlorophyll maximum of a mesotrophic lake. Aquat. Microb. Ecol. 2001, 24, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Griniene, E.; Sulcius, S.; Kuosa, H. Size-selective microzooplankton grazing on the phytoplankton in the Curonian Lagoon (SE Baltic Sea). Oceanologia 2016, 58, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Staniewski, M.A.; Short, C.M.; Short, S.M. Contrasting Community versus Population-Based Estimates of Grazing and Virus-Induced Mortality of Phytoplankton. Microb. Ecol. 2012, 64, 25–38. [Google Scholar] [CrossRef]
- Sterner, R.W.; Chrzanowski, T.H.; Elser, J.J.; George, N.B. Sources of nitrogen and phosphorus supporting the growth of bacterioplankton and phytoplankton in an oligotrophic Canadian shield lake. Limnol. Oceanogr. 1995, 40, 242–249. [Google Scholar] [CrossRef]
- Tijdens, M.; Van de Waal, D.B.; Slovackova, H.; Hoogveld, H.L.; Gons, H.J. Estimates of bacterial and phytoplankton mortality caused by viral lysis and microzooplankton grazing in a shallow eutrophic lake. Freshw. Biol. 2008, 53, 1126–1141. [Google Scholar] [CrossRef]
- Twiss, M.R.; Campbell, P.G.C.; Auclair, J.C. Regeneration, recycling, and trophic transfer of trace metals by microbial food-web organisms in the pelagic surface waters of Lake Erie. Limnol. Oceanogr. 1996, 41, 1425–1437. [Google Scholar] [CrossRef]
- Weisse, T.; Müller, H.; Pinto-Coelho, R.M.; Schweizer, A.; Springmann, D.; Baldringer, G. Response of the microbial loop to the phytoplankton spring bloom in a large prealpine lake. Limnol. Oceanogr. 1990, 35, 781–794. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.W.; Koch, F.; Marcoval, M.A.; Wilhelm, S.W.; Gobler, C.J. Mesozooplankton and microzooplankton grazing during cyanobacterial blooms in the western basin of Lake Erie. Harmful Algae 2012, 15, 26–35. [Google Scholar] [CrossRef]
- Collos, Y.; Vaquer, A.; Johnston, A.M.; Pons, V.; Bibent, B.; Richard, S. Carbon fixation, ammonium uptake and regeneration in an equatorial lake: Biological versus physical control. J. Plankton Res. 2001, 23, 263–270. [Google Scholar] [CrossRef]
- Chisholm, S.W. Phytoplankton size. In Primary Productivity and Biogeochemical Cycles in the Sea; Falkowski, P.G., Woodhead, A.D., Eds.; Plenum Press: New York, NY, USA, 1992; pp. 213–237. [Google Scholar]
- Laws, E.A. The importance of respiration losses in controlling the size distribution of marine phytoplankton. Ecology 1975, 56, 419–426. [Google Scholar] [CrossRef]
- Maranon, E. Inter-specific scaling of phytoplankton production and cell size in the field. J. Plankton Res. 2008, 30, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Maranon, E. Cell Size as a Key Determinant of Phytoplankton Metabolism and Community Structure. Annu. Rev. Mar. Sci. 2015, 7, 241–264. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.P.; Gillooly, J.F.; Brown, J.H. Linking the global carbon cycle to individual metabolism. Funct. Ecol. 2005, 19, 202–213. [Google Scholar] [CrossRef]
- Lopez-Urrutia, A.; San Martin, E.; Harris, R.P.; Irigoien, X. Scaling the metabolic balance of the oceans. PNAS 2006, 103, 8739–8744. [Google Scholar] [CrossRef] [Green Version]
- Rose, J.M.; Caron, D.A. Does low temperature constrain the growth rates of heterotrophic protists? Evidence and implications for algal blooms in cold waters. Limnol. Oceanogr. 2007, 52, 886–895. [Google Scholar] [CrossRef]
- Doney, S.C.; Glover, D.M.; Najjar, R.G. A new coupled, one-dimensional biological-physical model for the upper ocean: Applications to the JGOFS Bermuda Atlantic Time-series Study (BATS) site. Deep Sea Res. Part II 1996, 43, 591–624. [Google Scholar] [CrossRef]
- Laws, E.A.; Falkowski, P.G.; Smith, W.O.J.; Ducklow, H.; McCarthy, J.J. Temperature effects on export production in the open ocean. Glob. Biogeochem. Cycles 2000, 14, 1231–1246. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.R.; Totterdell, I.J. Production and export in a global ocean ecosystem model. Deep Sea Res. Part I Oceanogr. Res. Pap. 2001, 48, 1169–1198. [Google Scholar] [CrossRef]
- Taucher, J.; Oschlies, A. Can we predict the direction of marine primary production change under global warming? Geophys. Res. Lett. 2011, 38, 6. [Google Scholar] [CrossRef] [Green Version]
- Toseland, A.; Daines, S.J.; Clark, C.R.; Kirkham, A.; Strauss, J.; Uhlig, C.; Lenton, T.M.; Valentin, K.; Pearson, G.A.; Moulton, V.; et al. The impact of temperature on marine phytoplankton resource allocation and metabolism. Nat. Clim. Chang. 2013, 3, 979–984. [Google Scholar] [CrossRef]
- Stock, C.A.; Dunne, J.P.; John, J.G. Global-scale carbon and energy flows through the marine planktonic food web: An analysis with a coupled physical-biological model. Prog. Oceanogr. 2014, 120, 1–28. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pulsifer, J.; Laws, E. Temperature Dependence of Freshwater Phytoplankton Growth Rates and Zooplankton Grazing Rates. Water 2021, 13, 1591. https://doi.org/10.3390/w13111591
Pulsifer J, Laws E. Temperature Dependence of Freshwater Phytoplankton Growth Rates and Zooplankton Grazing Rates. Water. 2021; 13(11):1591. https://doi.org/10.3390/w13111591
Chicago/Turabian StylePulsifer, Jennifer, and Edward Laws. 2021. "Temperature Dependence of Freshwater Phytoplankton Growth Rates and Zooplankton Grazing Rates" Water 13, no. 11: 1591. https://doi.org/10.3390/w13111591
APA StylePulsifer, J., & Laws, E. (2021). Temperature Dependence of Freshwater Phytoplankton Growth Rates and Zooplankton Grazing Rates. Water, 13(11), 1591. https://doi.org/10.3390/w13111591