
Deep Pools: Ecological Sanctuaries for Steindachneridion melanodermatum, a Large Endemic and Endangered Pimelodid of the Iguaçu River

 , , ,
, , ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
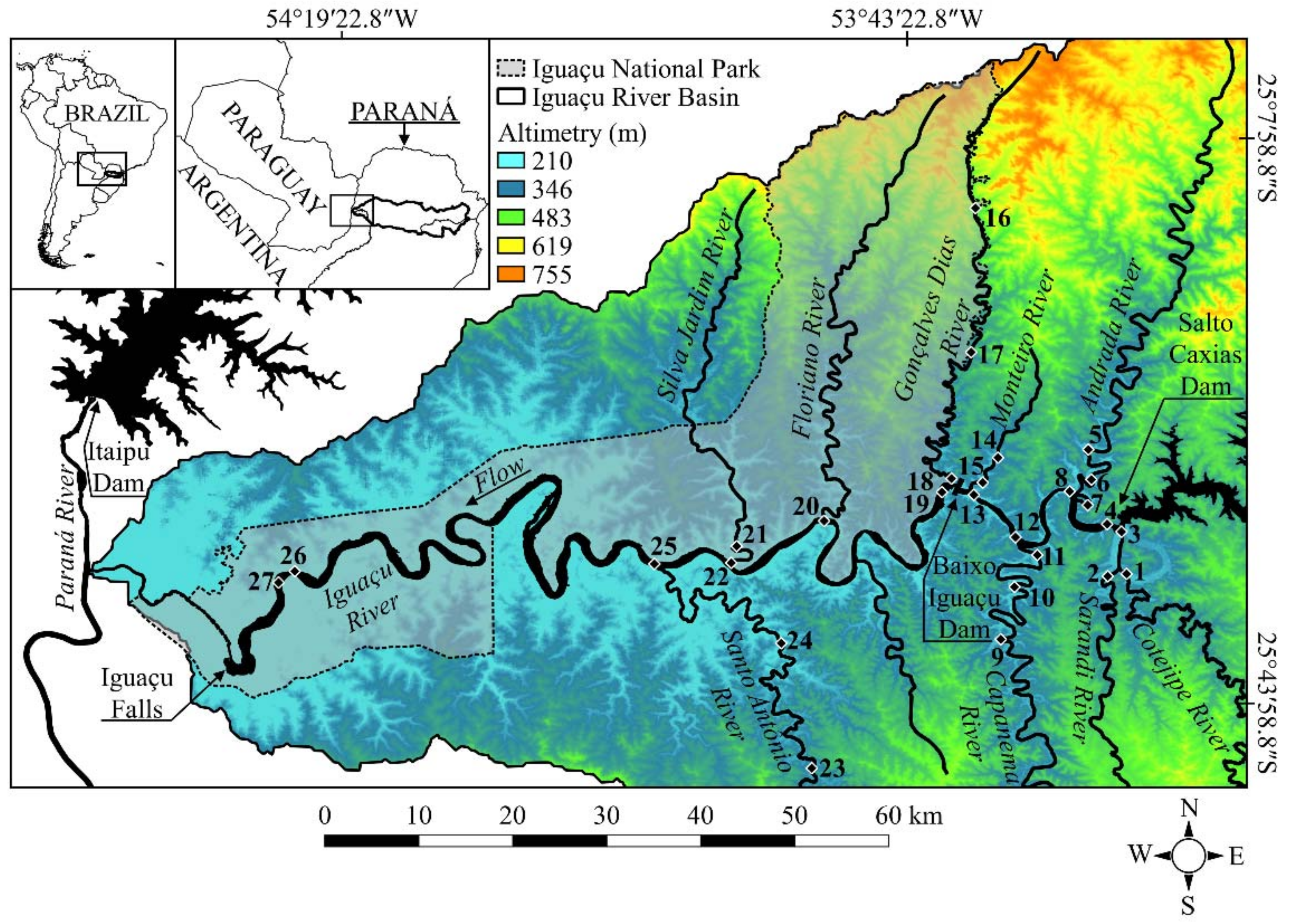
2.1. Study Area
2.2. Sampling and Data Analysis
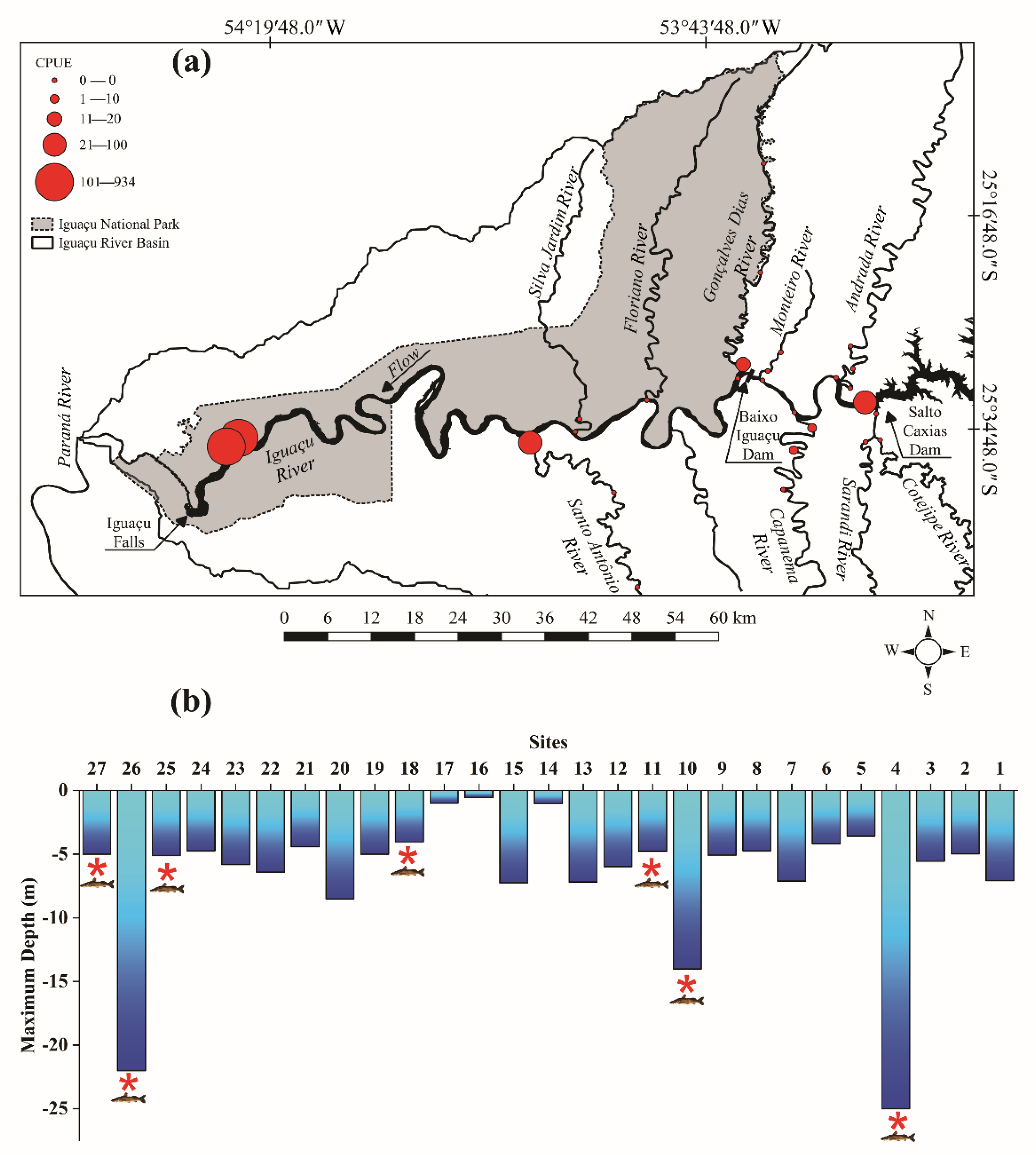
2.2.1. Spatial Distribution and Abundance
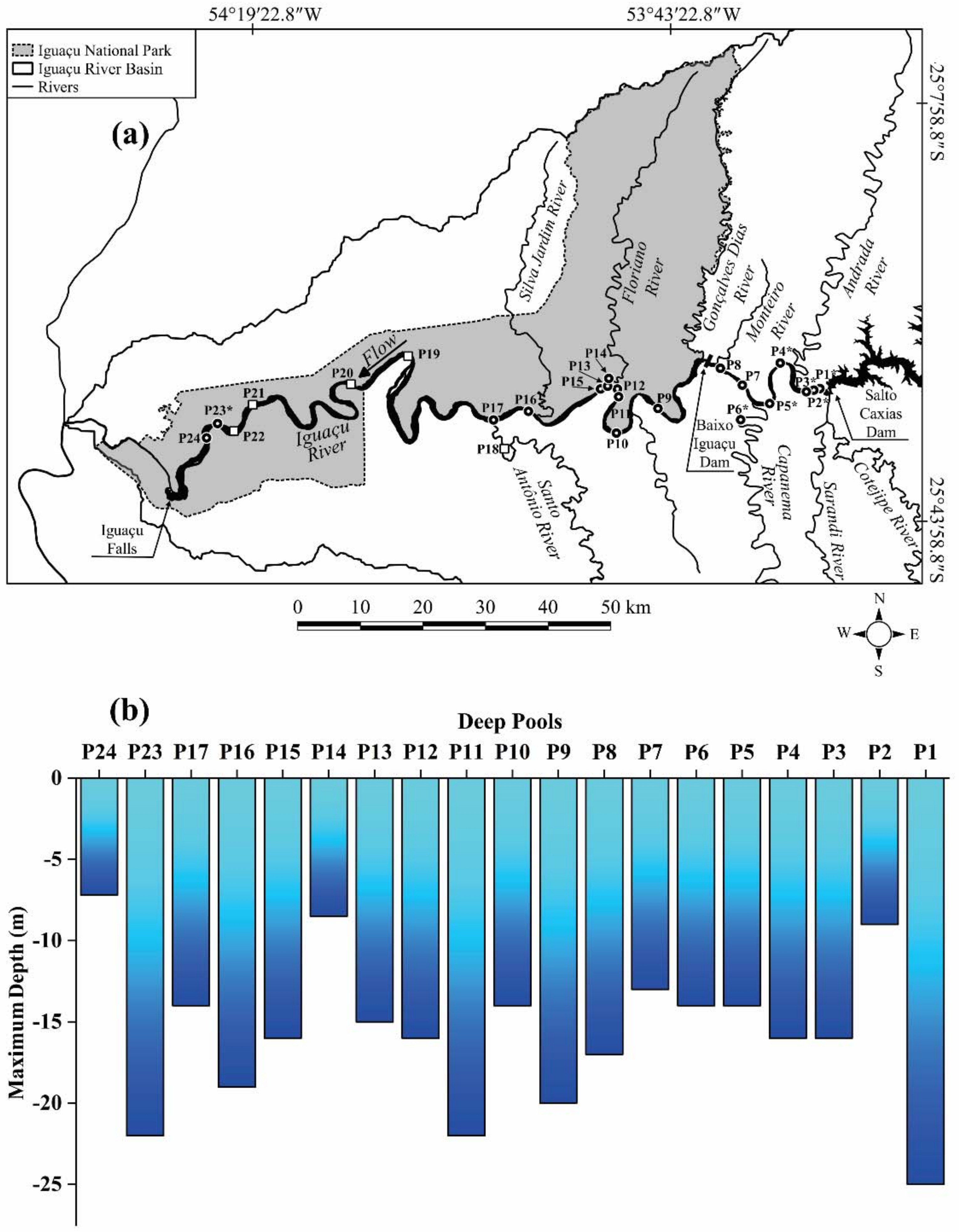
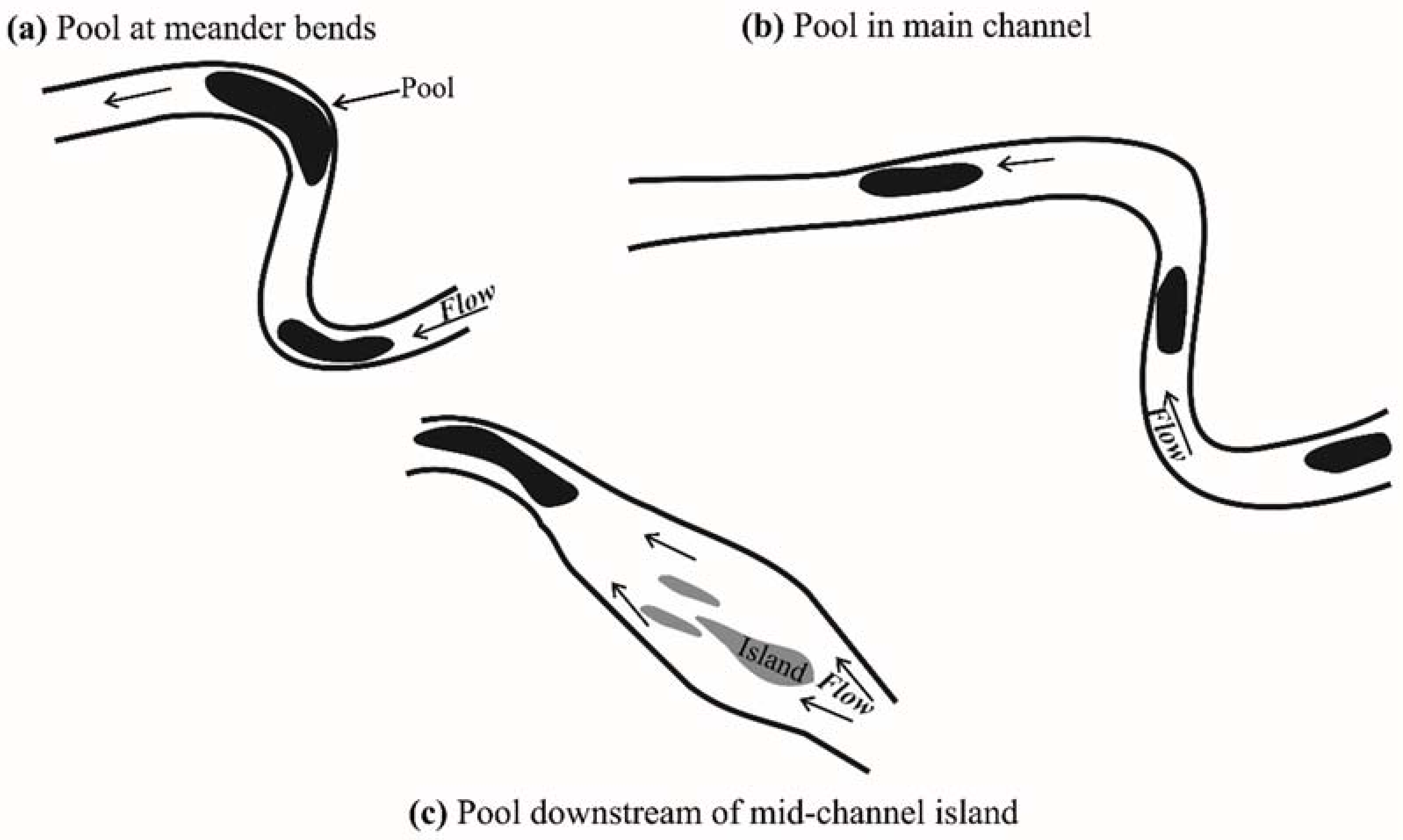
2.2.2. Deep Pools Mapping
3. Results
3.1. Spatial Distribution and Abundance
3.2. Deep Pools Characterization
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosa, R.S.; Lima, F.C.T. Os peixes brasileiros ameaçados de extinção. In Livro Vermelho da Fauna Brasileira Ameaçada de Extinção; Machado, A.B.M., Drummond, G.M., Paglia, A.P., Eds.; Fundação Biodiversitas: Belo Horizonte, Brazil, 2008; pp. 9–275. [Google Scholar]
- Brasil Portaria do Ministério do Meio Ambiente (MMA) [Brasilia 2014]. 445, de 17 de Dezembro de 2014. Reconhece Como espécies de Peixes e Invertebrados Aquáticos da Fauna Brasileira Ameaçadas de Extinção Aquelas Constantes da Lista Nacional Oficial de Espécies da Fauna Ameaçada. Available online: https://www.gov.br/agricultura/pt-br/assuntos/aquicultura-e-pesca/legislacao/legislacao-geral-da-pesca/portaria-mma-no-445-de-17-12-2014.pdf/view (accessed on 5 March 2020).
- Garavello, J.C. Revision of genus Steindachneridion (Siluriformes: Pimelodidae). Neotrop. Ichthyol. 2005, 3, 607–623. [Google Scholar] [CrossRef]
- Agostinho, A.A.; Gomes, L.C. Reservatório de Segredo: Bases Ecológicas Para o Manejo; Eduem: Maringá, Brazil, 1997. [Google Scholar]
- Garavello, J.C.; Pavanelli, C.S.; Suzuki, H.I. Caracterização da ictiofauna do rio Iguaçu. In Reservatório de Segredo: Bases Ecológicas Para o Manejo; Agostinho, A.A., Gomes, L.C., Eds.; Eduem: Maringá, Brazil, 1997; pp. 61–84. [Google Scholar]
- Baumgartner, G.; Pavanelli, C.S.; Baumgartner, D.; Bifi, A.G.; Debona, T.; Frana, V.A. Peixes do Baixo Rio Iguaçu; Eduem: Maringá, Brazil, 2012. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, L.A.M.; Gomes, E.; Artoni, R.F. Um método de reprodução induzida para o surubim Steindachneridion melanodermatum (Siluriformes: Pimelodidae) do Rio Iguaçu. Ciências Biológicas Saúde 2005, 11, 23–27. [Google Scholar] [CrossRef]
- Brehm, M.; Filippin, R.F.; De Moura, R.R. O impacto ambiental causado à ictiofauna do rio Iguaçu pela exploração do potencial hidrelétrico: O caso do surubim do Iguaçu (Steindachneridion melanodermatum). Rev. Bras. Ener. 2016, 22, 30–47. [Google Scholar]
- Assumpção, L.; Makrakis, S.; Silva, P.S.; Makrakis, M.C. Espécies de peixes ameaçadas de extinção no Parque Nacional do Iguaçu. Biodivers. Bras.-BioBrasil 2017, 7, 4–17. [Google Scholar]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lé Vêque, C.R.J.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- De Silva, S.S.; Abery, N.W.; Nguyen, T.T.T. Endemic freshwater finfish of Asia: Distribution and conservation status. Divers. Distrib. 2007, 13, 172–184. [Google Scholar] [CrossRef]
- Letcher, B.H.; Nislow, K.H.; Coombs, J.A.; O’Donnell, M.J.; Dubreuil, T.L. Population response to habitat fragmentation in a stream-dwelling brook trout population. PLoS ONE. 2007, 2, e1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nel, J.L.; Roux, D.J.; Abell, R.; Ashton, P.; Cowling, R.M.; Higgins, J.V.; Thieme, M.; Viers, J.H. Progress and challenges in freshwater conservation planning. Aquat. Conserv. Mar. Freshw. Ecosyst. 2009, 19, 474–485. [Google Scholar] [CrossRef]
- Reis, R.E.; Albert, J.S.; Di Dario, F.; Mincarone, M.M.; Petry, P.; Rocha, L.A. Fish biodiversity and conservation in South America. J. Fish Biol. 2016, 89, 12–47. [Google Scholar] [CrossRef] [Green Version]
- Echevarría, G.; González, N. Fish taxonomic and functional diversity in mesohabitats of the River Kakada, Caura National Park, Venezuela. Nat. Conserv. Res. 2018, 3, 21–39. [Google Scholar] [CrossRef]
- Damme, P.A.V.; Córdova-Clavijo, L.; Baigún, C.; Hauser, M.; Dória, C.R.C.; Duponchelle, F. Upstream dam impacts on gilded catfish Brachyplatystoma rousseauxii (Siluriformes: Pimelodidae) in the Bolivian Amazon. Neotrop. Ichthyol. 2019, 17. [Google Scholar] [CrossRef]
- Schletterer, M.; Shaporenko, S.I.; Kuzovlev, V.V.; Minin, A.E.; Van Geest, G.J.; Middelkoop, H.; Górski, K. The Volga: Management issues in the largest river basin in Europe. River Res. Applic. 2019, 35, 510–519. [Google Scholar] [CrossRef]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [Green Version]
- Dagosta, F.C.P.; Pinna, M.; Peres, C.A.; Tagliacollo, V.A. Existing protected areas provide a poor safety-net for threatened Amazonian fish species. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 1167–1189. [Google Scholar] [CrossRef]
- Cetra, M.; Soares, L.M.S.; Pinheiro, R.F.M. Peixes de riachos e novas Unidades de Conservação no sul da Bahia. Pan-Am. J. Aquat Sci. 2010, 5, 11–21. [Google Scholar]
- Barletta, M.; Jaureguizar, A.J.; Baigun, C.; Fontoura, N.F.; Agostinho, A.A.; Almeida-Val, V.M.F.; Val, A.L.; Torres, R.A.; Jimenes-Segura, L.F.; Giarrizzo, T.; et al. Fish and aquatic habitat conservation in South America: A continental overview with emphasis on neotropical systems. J. Fish Biol. 2010, 76, 2118–2176. [Google Scholar] [CrossRef]
- Instituto Chico Mendes de Conservação da Biodiversidade. Livro Vermelho da Fauna Brasileira Ameaçada de Extinção, 1st ed.; ICMBio/MMA: Brasília, Brazil, 2018; Volume VI-Peixes, pp. 208–222. [Google Scholar]
- Reis, R.E.; Albert, J.S.; Di Dario, F.; Mincarone, M.M.; Petry, P.; Rocha, L.A. Reply to Vitule et al. (2017): Comment on ‘Fish biodiversity and conservation in South America by Reis et al. (2016)’. J. Fish Biol. 2017, 90, 1191–1195. [Google Scholar] [CrossRef] [PubMed]
- Instituto Chico Mendes de Conservação da Biodiversidade, Lista de Espécies Ameaçadas [ICMBio 2014]. Available online: https://www.icmbio.gov.br/portal/especies-ameacadas-destaque (accessed on 6 April 2020).
- Barletta, M.; Cussac, V.E.; Agostinho, A.A.; Baigún, C.; Okada, E.K.; Carlos Catella, A.; Fontoura, N.F.; Pompeu, P.S.; Jimenez-Segura, L.F.; Batista, V.S.; et al. Fisheries ecology in South American river basins. In Freshwater Fisheries Ecology; Craig, J.F., Ed.; Wiley-Blackwell: Oxford, UK, 2015; pp. 311–348. [Google Scholar]
- Campbell, T.; Pin, K.; Ngor, P.B.; Hogan, Z. Conserving Mekong megafishes: Current status and critical threats in Cambodia. Water 2020, 12, 1820. [Google Scholar] [CrossRef]
- He, F.; Bremerich, V.; Zarfl, C.; Geldmann, J.; Langhans, S.D.; David, J.N.W.; Darwall, W.; Tockner, K.; Jähnig, S.C. Freshwater megafauna diversity: Patterns, status and threats. Divers. Distrib. 2018, 24, 1395–1404. [Google Scholar] [CrossRef] [Green Version]
- Carrizo, S.F.; Jähnig, S.C.; Bremerich, V.; Freyhof, J.; Harrison, I.; He, F.; Langhans, S.D.; Tockner, K.; Zarfl, C.; Darwall, W. Freshwater megafauna: Flagships for freshwater biodiversity under threat. BioScience 2017, 67, 919–927. [Google Scholar] [CrossRef]
- He, F.; Zarfl, C.; Bremerich, V.; David, J.N.W.; Hogan, Z.; Kalinkat, G.; Tockner, K.; Jähnig, S.C. The global decline of freshwater megafauna. Glob. Chang. Biol. 2019, 25, 3883–3892. [Google Scholar] [CrossRef] [PubMed]
- Brito, D. Overcoming the linnean shortfall: Data deficiency and biological survey priorities. Basic Appl. Ecol. 2010, 11, 709–713. [Google Scholar] [CrossRef]
- Hogan, Z.S. Ecology and conservation of large-bodied freshwater catfish: A global perspective. Am. Fish. Soc. Symp. 2011, 77, 39–53. [Google Scholar]
- Muchlisin, Z.A. Distributions of the endemic and threatened freshwater fish depik, Rasbora tawarensis Weber and de Beaufort, 1916 in Lake Laut Tawar, Aceh Province, Indonesia-Songklanakarin. J. Sci. Edu. Technol. 2013, 35, 483–488. [Google Scholar]
- Tripe, J.A.; Guy, C.S. Spatial and temporal variation in habitat and fish community characteristics in a Kansas Flint Hills stream. Ecol. Freshw. Fish 1999, 8, 216–226. [Google Scholar] [CrossRef]
- Argent, D.G.; Bishop, J.A.; Stauffer, J.R., Jr.; Carline, R.F.; Myers, W.L. Predicting freshwater fish distributions using landscape-level variables. Fish Res. 2003, 60, 17–32. [Google Scholar] [CrossRef]
- Ko, M.H.; Moon, S.J.; Hong, Y.K.; Lee, G.Y.; Bang, I.C. Distribution status and habitat characteristics of the endangered species, Lethenteron reissner (Petromyzontiformes:Petromyzontidae) in Korea. Korean J. Ichthyol. 2013, 25, 189–199. [Google Scholar]
- Assumpção, L.; Fávaro, L.F.; Makrakis, S.; Silva, P.S.; Pini, S.F.R.; Kashiwaqui, E.A.L.; Makrakis, M.C. Population structure and reproduction of Steindachneridion melanodermatum (Siluriformes: Pimelodidae), a large endemic catfish to Neotropical ecoregion. Mar. Freshw. Res. 2021. [Google Scholar] [CrossRef]
- Pini, S.F.R.; Makrakis, M.C.; Neves, M.P.; Makrakis, S.; Shibatta, O.A.; Kashiwaqui, E.A. Ichthyofauna in the last free-flowing river of the Lower Iguaçu basin: The importance of tributaries for conservation of endemic species. ZooKeys 2021, 1014, 183. [Google Scholar] [CrossRef]
- Baird, I.G.; Flaherty, M.S. Mekong River fish conservation zones in southern Laos: Assessing effectiveness using local ecological knowledge. Environ. Manag. 2005, 36, 439–454. [Google Scholar] [CrossRef]
- Eletrosul, O. O impacto ambiental da ação do homem sobre a natureza-rio Iguaçu, Paraná, Brasil. In Reconhecimento da Ictiofauna, Modificações Ambientais e Usos Múltiplos dos Reservatórios; Florianópolis: Eletrosul, Brazil, 1978. [Google Scholar]
- Júlio-Júnior, H.F.; Bonecker, C.; Agostinho, A.A. Reservatório de segredo e sua inserção na bacia do rio Iguaçu. In Reservatório de Segredo: Bases Ecológicas Para o Manejo; Agostinho, A.A., Gomes, L.C., Eds.; Eduem: Maringá, Brazil, 1997; pp. 1–17. [Google Scholar]
- Maack, R. Geografia Física do Estado do Paraná; Rio de Janeiro: Livraria José Olympio, Brazil, 1981. [Google Scholar]
- Agostinho, A.A.; Gomes, L.C.; Pelicice, F.M. Ecologia e Manejo de Recursos Pesqueiros em Reservatórios do Brasil; Eduem: Maringá, Brazil, 2007. [Google Scholar]
- Celestino, F.E.; Celestino, L.F.; da Silva, F.M.J.; Kashiwaqui, A.L.E.; Makrakis, C.M.; Makrakis, S. Environmental assessment in neotropical watersheds: A multi-factorial approach. Sustainability 2019, 11, 490. [Google Scholar] [CrossRef] [Green Version]
- Gibson-Reinemer, D.K.; Ickes, B.S.; Chick, J.H. Development and assessment of a new method for combining catch per unit effort data from different fish sampling gears: Multigear mean standardization (MGMS). Can. J. Fish. Aquat. Sci. 2016, 74, 8–14. [Google Scholar] [CrossRef]
- Resk, R.P.; Bezerra-Neto, J.F.; Coelho, R.M.P. Nova batimetria e avaliação de parâmetros morfométricos da Lagoa da Pampulha (Belo Horizonte, Brasil). Revista Geogr. 2007, 5, 24–37. [Google Scholar]
- Halls, A.S.; Conlan, I.; Wisesjindawat, W.; Phouthavongs, K.; Viravong, S.; Chan, S.; Vu, V.A. Atlas of deep pools in the Lower Mekong River and some of its tributaries. In MRC Technical Paper No. 31; Mekong River Commission: Phnom Penh, Cambodia, 2013. [Google Scholar]
- Jowett, I.G. A method for objectively identifying pool, run, and riffle habitats from physical measurements. N. Z. J. Mar. Freshw. Res. 1993, 27, 241–248. [Google Scholar] [CrossRef]
- Keppeler, F.; Hallwass, G.; Silvano, R. Influence of protected areas on fish assemblages and fisheries in a large tropical river. Oryx 2017, 51, 268–279. [Google Scholar] [CrossRef] [Green Version]
- Setyawan, E.; Sianipar, A.B.; Erdmann, M.V.; Fischer, A.M.; Haddy, J.A.; Beale, C.S.; Lewis, S.A.; Mambrasar, R. Site fidelity and movement patterns of reef manta rays (Mobula alfredi: Mobulidae) using passive acoustic telemetry in northern Raja Ampat, Indonesia. Nat. Conserv. Res. 2018, 3, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Zhang, H.; Wang, C.; Wu, J.; Wei, Q.; Du, H.; Li, J.; Ye, H. Are river protected areas sufficient for fish conservation? Implications from large-scale hydroacoustic surveys in the middle reach of the Yangtze River. BMC Ecol. 2019, 19, 42. [Google Scholar] [CrossRef] [Green Version]
- Loury, E.K.; Ainsley, S.M. Identifying indicators to evaluate community-managed freshwater protected areas in the lower Mekong basin: A review of marine and freshwater examples. Water 2020, 12, 3530. [Google Scholar] [CrossRef]
- Sweke, E.A.; Assam, J.M.; Chande, A.I.; Mbonde, A.S.; Mosha, M.; Mtui, A. Comparing the performance of protected and unprotected areas in conserving freshwater fish abundance and biodiversity in Lake Tanganyika, Tanzania. Int. J. Ecol. 2016, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, U.K.; Pathak, A.K.; Tyagi, L.K.; Srivastava, S.M.; Singh, S.P.; Dubey, V.K. Biodiversity of freshwater fish of a protected river in India: Comparison with unprotected habitat. Rev. Biol. Trop. 2013, 61, 161–172. [Google Scholar] [CrossRef]
- Jézéquel, C.; Tedesco, P.A.; Darwall, W.; Dias, M.S.; Frederico, R.G.; Hidalgo, M.; Hugueny, B.; Maldonado-Ocampo, J.; Martens, K.; Ortega, H.; et al. Freshwater fish diversity hotspots for conservation priorities in the Amazon Basin. Conserv. Biol. 2020, 34, 4. [Google Scholar] [CrossRef]
- Chan, S.; Putrea, S.; Hortle, K.G. Using local knowledge to inventory deep pools, important fish habitats in Cambodia. MRC Conf. Ser. 2005, 5, 57–76. [Google Scholar]
- Poulsen, A.F.; Valbo-Jørgensen, J. Deep pools in the Mekong River. Catch Cult. 2001, 7, 8–9. [Google Scholar]
- Poulsen, A.F.; Poeu, O.; Virvong, S.; Suntornratana, U.; Tung, N.T. Deep pools as dry season fish habitats in the Mekong Basin. In MRC Technical Paper No. 4; Mekong River Commission: Phnom Penh, Cambodia, 2002. [Google Scholar]
- Chan, S.; Putrea, S.; Hortle, H.G. Using local knowledge to inventory deep pools, important fish habitats in Cambodia. In Proceedings of the 6th Technical Symposium on Mekong Fisheries, Pakse, Laos, 26–28 November 2003; Burnhill, T.J., Hewitt, M.M., Eds.; Mekong River Commission: Vientiane, Laos, 2003; pp. 57–76. [Google Scholar]
- Baird, I.G. Local ecological knowledge and small-scale freshwater fisheries management in the Mekong River in Southern Laos. In Putting Fishers’ Knowledge in Fisheries Science and Management; Haggan, N., Neis, B., Baird, I.G., Eds.; Blackwell Sciences and UNESCO: Oxford, UK, 2005; pp. 247–266. [Google Scholar]
- Baird, I.G. The last anecdote. In Fishers’ Knowledge in Fisheries Science and Management; Haggan, N., Neis, B., Baird, I.G., Eds.; Blackwell Sciences and UNESCO: Oxford, UK, 2005; pp. 433–437. [Google Scholar]
- Stefferud, J.A.; Gido, K.B.; Propst, D.L. Spatially variable response of native fish assemblages to discharge, predators and habitat characteristics in an arid-land river. Freshw. Biol. 2011, 56, 1403–1416. [Google Scholar] [CrossRef]
- Makwinja, R.; Chapotera, M.; Likongwe, P.; Banda, J.; Chijere, A. Location and roles of deep pools in likangala river during 2012 recession period of Lake Chilwa Basin. Int. J. Ecol. 2014, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Zaniboni-Filho, E.; Schulz, U.H. Migratory fishes of the Uruguay River. In Migratory Fishes of the South América: Biology, Fisheries and Conservation Status; Carosfeld, J., Harvey, B., Ross, C., Baer, A., Eds.; World Fisheries Trust: Victoria, Australia, 2003; pp. 161–192. [Google Scholar]
- Welcomme, R.L. River Fisheries; FAO Fisheries Technical Paper; FAO: Rome, Italy, 1985. [Google Scholar]
- Caneppele, D.; Pompeu, P.; Garavello, J. Surubim do Paraíba Steindachneridion parahybae. In Livro Vermelho da Fauna Brasileira Ameaçada de Extinção; Machado, A.B.M., Drummond, G.M., Paglia, A.P., Eds.; Fundação Biodiversitas: Belo Horizonte, Brazil, 2008; pp. 236–238. [Google Scholar]
- Weingartner, M.; Fracalossi, D.M.; Nuñer, A.P.O.; Zaniboni-Filho, E. Conservação Genética. In Reservatório de Machadinho: Peixes, Pesca e Tecnologias de Criação; Nuñer, A.P.O., Zaniboni-Filho, E., Eds.; Orgs. Editora da UFSC: Florianópolis, Brazil, 2012; pp. 241–255. [Google Scholar]
- Reynalte-Tataje, D.A.; Nuñer, A.P.O.; Nunes, M.C.; Garcia, V.; Lopes, C.A.; Zaniboni-Filho, E. Spawning of migratory fish species between two reservoirs of the upper Uruguay River, Brazil. Neotrop. Ichthyol. 2012, 10, 829–835. [Google Scholar] [CrossRef] [Green Version]
- Beux, L.F.; Zaniboni-Filho, E. Produção pesqueira. In Reservatório de Machadinho: Peixes, Pesca e Tecnologias de Criação; Nuñer, A.P.O., Zaniboni-Filho, E., Eds.; EDUFSC: Florianópolis, Brazil, 2012; pp. 63–81. [Google Scholar]
- Schork, G.; Hermes-Silva, S.; Beux, L.F.; Zaniboni-Filho, E.; Nuñer, A.P.O. Diagnóstico da pesca artesanal na usina hidrelétrica de Machadinho, alto rio Uruguai-Brasil. Bol. Inst. Pesca 2012, 38, 97–108. [Google Scholar]
- Wantzen, K.M.; Drago, E.; Silva, C.J. Aquatic habitats of the Upper Paraguay river-floodplain-system and parts of the Pantanal (Brazil). Ecohydrol. Hydrobiol. 2005, 5, 107–126. [Google Scholar]
- Chapman, L.; Kramer, D.; Chapman, C. Population dynamics of the fish Poecilia gillii (Poeciliidae) in pools of an intermittent tropical stream. J. Anim. Ecol. 1991, 60, 441–453. [Google Scholar] [CrossRef]
- Baird, I.G.; Kisouvannalath, P.; Inthaphaisy, V.; Phylavanh, B. The potential for ecological classification as a tool for establishing and monitoring fish conservation zones in the Mekong River. In Environmental Protection and Community Development in Siphandone Wetland Project; Technical Report; CESVI Cooperation and Development: Pakse, Laos, 1998; p. 28. [Google Scholar]
- Timpe, K.; Kaplan, D. The changing hydrology of a dammed Amazon. Sci. Adv. 2017, 3. [Google Scholar] [CrossRef] [Green Version]
- Schletterer, M.; Kuzovlev, V.V.; Zhenikov, Y.N.; Tuhtan, J.A.; Haidvogl, G.; Friedrich, T.; Gorski, K.; Füreder, L. Fish fauna and fisheries of large European rivers: Examples from the Volga and the Danube. Hydrobiologia 2018, 814, 45–60. [Google Scholar] [CrossRef]
- Souza-Cruz-Buenaga, F.V.A.; Espig, S.A.; Castro, T.L.C.; Santos, M.A. Environmental impacts of a reduced flow stretch on hydropower plants. Braz. J. Biol. 2019, 79, 470–487. [Google Scholar] [CrossRef]
- Poddubny, A.G.; Galat, D.L. Habitat associations of upper Volga river fishes: Effects of reservoirs. Regul. Rivers Res. Mgmt. 1995, 11, 67–84. [Google Scholar] [CrossRef]
- Baird, I.G. Strength in diversity: Fish sanctuaries and deep-water pools in Lao PDR. Fish Manag. Ecol. 2006, 13, 1–8. [Google Scholar] [CrossRef]
- Makrakis, M.C.; Miranda, L.E.; Makrakis, S.; Fontes, H.M., Jr.; Morlis, W.G.; Dias, J.H.P.; Garcia, J.O. Diversity in migratory patterns among Neotropical fishes in a highly regulated river basin. J. Fish. Biol. 2012, 81, 866–881. [Google Scholar] [CrossRef]
- Da Silva, P.S.; Makrakis, M.C.; Miranda, L.E.; Makrakis, S.; Assumpção, L.; Paula, S.; Dias, J.H.P.; Marques, H. Importance of reservoir tributaries to spawning of migratory fish in the upper Paraná River. River Res. Appl. 2015, 31, 313–322. [Google Scholar] [CrossRef]
- Da Silva, P.S.; Miranda, L.E.; Makrakis, S.; Assumpção, L.; Dias, J.H.P.; Makrakis, M.C. Tributaries as biodiversity preserves: An ichthyoplankton perspective from the severely impounded Upper Paraná River. Aquat. Conserv Mar. Freshw. Ecosyst. 2019, 29, 258–269. [Google Scholar] [CrossRef]
- Mitamura, H.; Arai, M.; Yamagishi, Y.; Kawabara, Y.; Mitsunaga, Y.; Khachaphichat, M.; Viputhanumas, T. Habitat use and movement of hatchery-reared F2 Mekong giant catfish in the Mae Peum reservoir, Thailand, studied by acoustic telemetry. Fish Sci. 2009, 75, 175–182. [Google Scholar] [CrossRef]
- Saunders, D.L.; Meeuwig, J.J.; Vincent, A.C.J. Freshwater protected areas: Strategies for conservation. Conserv. Biol. 2002, 16, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Sajan, S.; Mercy, T.V.A.; Malika, V. Age, growth and population dynamics of an endangered fish Sahyadria denisonii (Day 1865) from the Western Ghats hotspot of India. Asian Fish. Sci. J. 2015, 28, 130–142. [Google Scholar] [CrossRef]
- Casciotta, J.; Almirón, A.; Ciotek, L.; Giorgis, P.; Říčan, O.; Piálek, L.; Dragová, K.; Croci, Y.; Montes, M.; Iwaszkiw, J.; et al. Visibilizando lo invisible. Un relevamiento de la diversidad de peces del Parque Nacional Iguazú, Misiones, Argentina. Rev. História Nat. 2016, 6, 5–77. [Google Scholar]
- Grill, G.; Lehner, B.; Thieme, M.; Geenen, B.; Tickner, D.; Antonelli, F.; Babu, S.; Borrelli, P.; Cheng, L.; Crochetiere, H.; et al. Mapping the world’s free-flowing rivers. Nature 2019, 569, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Cutler, J.S.; Olivos, J.A.; Sidlauskas, B.; Arismendi, I. Habitat loss due to dam development may affect the distribution of marine-associated fishes in Gabon, Africa. Ecosphere 2020, 11, e03024. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | Environment | River | Site | Latitude | Longitude | Width of the River (m) |
|---|---|---|---|---|---|---|
| Outside | Tributary | Cotejipe River | 1 | 25°35′17.04″ S | 053°29’56.58″ W | 25.4 |
| 3 | 25°33′9.54″ S | 053°29′46.92″ W | 54.2 | |||
| Outside | Tributary | Sarandi River | 2 | 25°35′10.74″ S | 053°30′7.44″ W | 12.6 |
| Outside | Main channel | Iguaçu River | 4 | 25°32′30.18″ S | 053°30′37.98″ W | 373.2 |
| 8 | 25°30′48.00″ S | 053°32′40.62″ W | 501.4 | |||
| 12 | 25°31′2.28″ S | 053°32′34.44″ W | 586.9 | |||
| 13 | 25°29′29.70″ S | 053°31′55.08″ W | 815.6 | |||
| Outside | Tributary | Andrada River | 7 | 25°27′36.18″ S | 053°31′51.69″ W | 120.4 |
| 6 | 25°33′49.14″ S | 053°36′16.92″ W | 59.9 | |||
| 5 | 25°34′16.26″ S | 053°35′52.68″ W | 35.6 | |||
| Outside | Tributary | Capanema River | 11 | 25°36′8.40″ S | 053°36′46.98″ W | 462.7 |
| 10 | 25°39′54.84″ S | 053°37′15.66″ W | 82.7 | |||
| 9 | 25°30′42.58″ S | 053°39′5.76″ W | 52.4 | |||
| Outside | Tributary | Monteiro River | 15 | 25°30′25.38″ S | 053°39′27.24″ W | 231.0 |
| 14 | 25°28′12.96″ S | 053°37′39.00″ W | 9.2 | |||
| INP | Main channel | Iguaçu River | 19 | 25°29’57.54″ S | 053°40’53.52″ W | 750.7 |
| INP | Tributary | Gonçalves Dias River | 18 | 25°29’57.06″ S | 053°40’40.50″ W | 25.9 |
| 17 | 25°21’48.12″ S | 053°39’18.00″ W | 63.5 | |||
| 16 | 25°12’58.98″ S | 053°39’0.06″ W | 13.6 | |||
| INP | Tributary | Floriano River | 20 | 25°32’14.82″ S | 053°48’31.98″ W | 497.5 |
| INP | Tributary | Silva Jardim River | 22 | 25°34’51.24″ S | 053°54’43.68″ W | 26.6 |
| 21 | 25°34’11.09″ S | 053°54’20.36″ W | 40.2 | |||
| Outside | Tributary | Santo Antônio River | 25 | 25°35′17.16″ S | 053°59′25.20″ W | 49.3 |
| 24 | 25°40′25.80″ S | 053°51′15.90″ W | 37.0 | |||
| 23 | 25°48′6.28″ S | 053°49′28.35″ W | 34.3 | |||
| INP | Main channel | Iguaçu River | 26 | 25°35′38.00″ S | 054°21′57.10″ W | 372.3 |
| 27 | 25°37′13.20″ S | 054°23′29.20″ W | 932.6 |
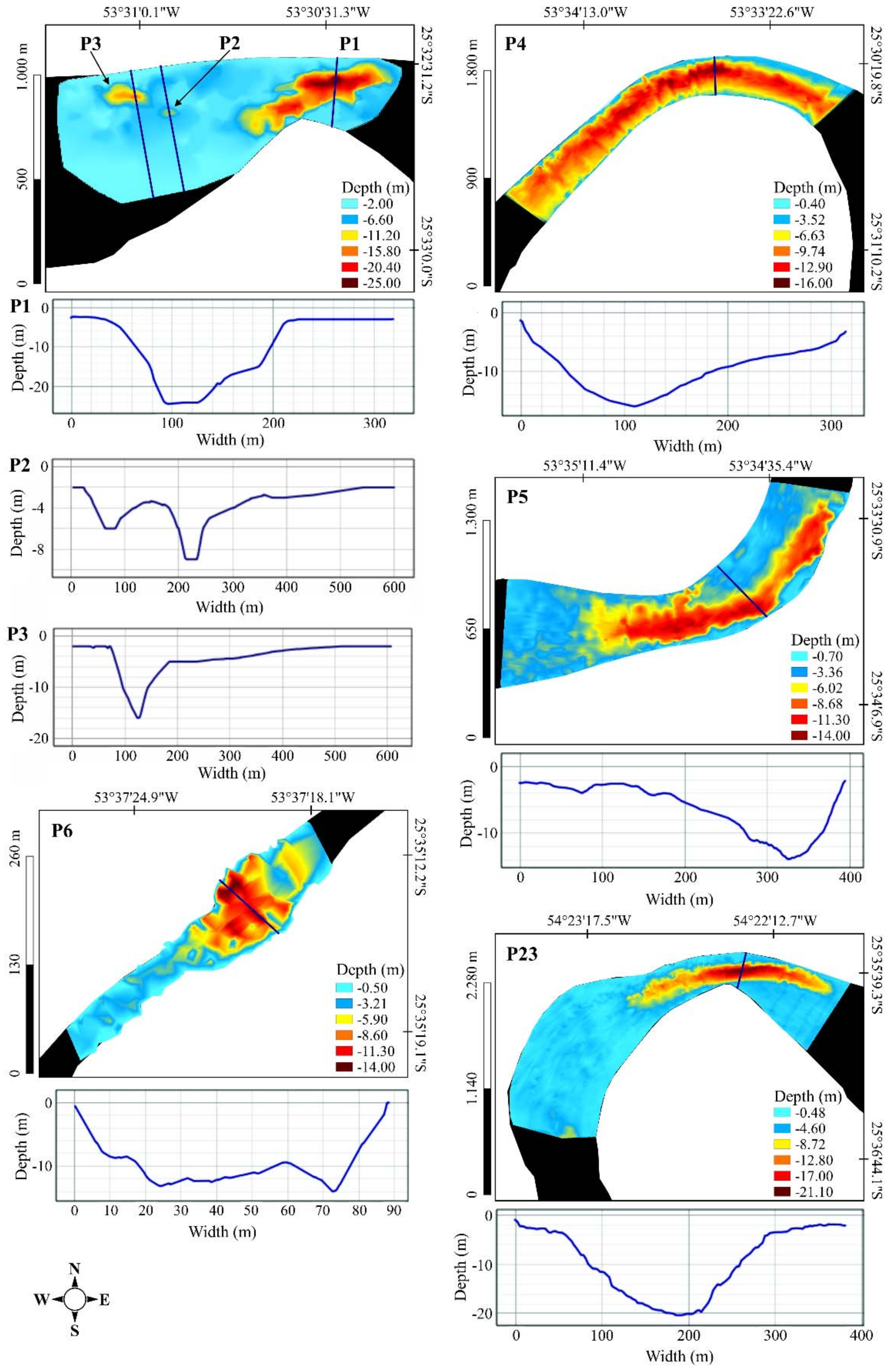
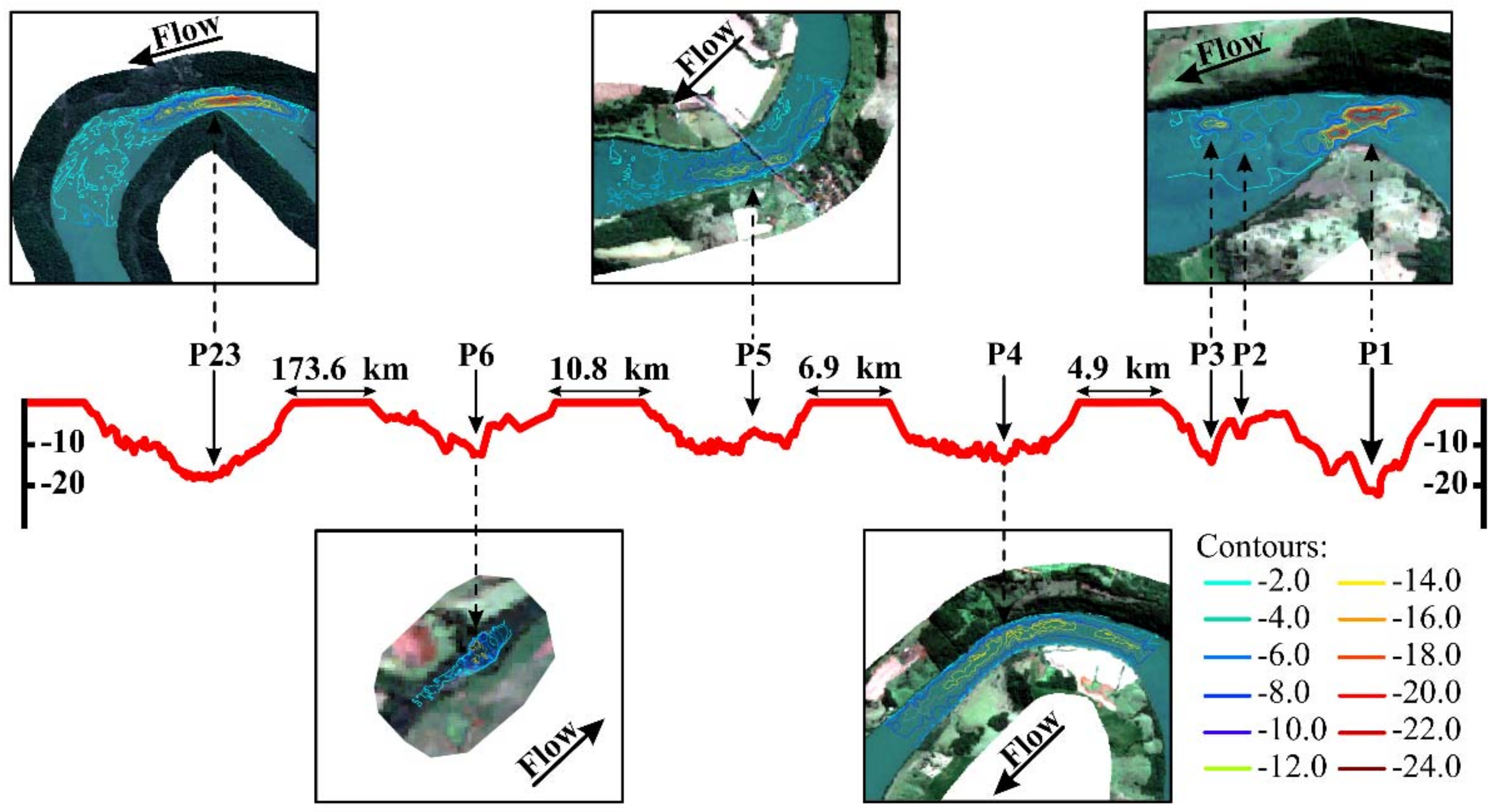
| Deep Pool | Depth (m) | Location | Position | Area (ha) | Deep Pool Area (ha) | Deep Pool Length (m) | Transect Width (m) | Characteristics | |
|---|---|---|---|---|---|---|---|---|---|
| Min | Max | ||||||||
| P1 * | 2.4 | 25.0 | MC | a | 67.06 | 10.186 | 766.511 | 647.03 | Rocky bed, large rocks, and rapids. The river’s right and left banks had a greater concentration of pastures and few forest fragments (Figure 7). |
| P2 * | 0.4 | 16.0 | MC | b | 67.06 | 0.451 | 111.542 | 605.456 | Rapids with a riparian zone that was poorly preserved with forest fragments and a predominance of pasture on both banks (Figure 7). |
| P3 * | 0.9 | 14.0 | MC | b | 67.06 | 1.847 | 277.006 | 615.277 | Rapids with a riparian zone that was poorly preserved with forest fragments and predominance of pasture on both banks (Figure 7). |
| P4 * | 0.4 | 16.0 | MC | a | 93.33 | 47.255 | 2994.951 | 387.32 | Rapids with the left bank impacted by agricultural activities and almost nonexistent riparian forest on both banks (Figure 7). |
| P5 * | 0.9 | 14.0 | MC | a | 87.91 | 31.559 | 1781.068 | 474.60 | Some stretches with low depths, riffles, where the navigation was impossible due to turbulence, other rapids stretches, rocky outcrops, and the presence of a small island in the middle of the river. Human activities, such as agricultural areas and pastures, on both banks (Figure 7). |
| P6 * | 0.5 | 14.0 | TR | b | 2.06 | 1.046 | 244.265 | 60.66 | Riffles upstream of the pool. Human activities, such as agricultural areas, on the right bank, with backwaters in the pool. |
| P7 | 1.0 | 13.0 | MC | b | 85.281 | - | - | - | Rapids stretches near riffles, with an approximate width of 416 m. Both banks with a predominance of pastures and agricultural areas, and a riparian forest that was barely preserved. |
| P8 | 1.0 | 17.0 | MC | b | 23.559 | - | - | - | Rapids stretches located close to shallow water areas, namely, rifles, with exposed rocks before and after the stretch, with an approximate width of 290 m. Right bank had a small section of riparian vegetation that was barely preserved. On the left bank, there were pasture and agricultural areas. |
| P9 | 2.0 | 20.0 | MC | b | 108.833 | - | - | - | Rapids stretch located upstream of an island, with strong running areas and an approximate width of 280 m. Right bank had a predominance of preserved riparian forest. On the left bank, there were pasture, agricultural areas, and small forest fragments that were barely preserved. |
| P10 | 1.5 | 14.0 | MC | a | 1.253 | - | - | - | Rapids stretches with riffles, shallow water, and exposed rocks, as well as a pool that was located in a steep meander, upstream to an island in the middle of the river, with an approximate width of 78 m. Right bank had a predominance of preserved riparian vegetation. On the left bank, there were pasture, agricultural areas, and small forest fragments that were barely preserved. |
| P11 | 1.5 | 22.0 | MC | a | 27.010 | - | - | - | Rapids stretch upstream of an accentuated meander, with an approximate width of 281 m. The right bank had a predominance of preserved riparian forest. On the left bank, there were pasture, agricultural areas, and small forest fragments that were barely preserved. |
| P12 | 1.5 | 16.0 | MC | a | 46.628 | - | - | - | Rapids stretch in a meander with an approximate width of 320 m. The right bank exhibited dense riparian forest. The left bank had a predominance of agricultural area and small forest fragments that were barely preserved. |
| P13 | 1.0 | 15.0 | MC | a | 41.399 | - | - | - | Rapids stretch in a meander with an approximate width of 409 m. The right bank exhibited dense riparian forest. The left bank had a predominance of agricultural area and small forest fragments that were barely preserved. |
| P14 | 1.0 | 8.5 | TR | b | - | - | - | - | Rapids stretch with a dense riparian forest that was preserved on both banks. |
| P15 | 1.5 | 16.0 | MC | a | 97.655 | - | - | - | Rapids stretch upstream of a meander, with an approximate width of 319 m. The right bank had a predominantly riparian forest that was preserved. The left bank presented an agricultural area and small forest fragments that were barely preserved. |
| P16 | 1.0 | 19.0 | MC | b | 176.230 | - | - | - | Rapids stretch that was approximately 268 m in width and located in a meander. The right bank had preserved riparian vegetation. The left bank presented a predominantly agricultural area with poorly preserved forest fragments. |
| P17 | 1.0 | 14.0 | MC | a | 46.976 | - | - | - | Rapids stretch in a meander with an approximate width of 335 m. The right bank had preserved riparian vegetation. The left bank presented a higher concentration of pastures with shrubby vegetation, an agricultural area, and small forest fragments that were barely preserved. |
| P23 * | 0.8 | 22.0 | MC | a, c | 258.39 | 43.693 | 2303.019 | 153.27 | Riffle stretches in some shallow areas, bedrock substrate forming extensive areas with large rocks, backwater areas that included foams on the water surface, and an island upstream of the pool (Taquaras’ Island). The deep pool was in a meander. Both banks had dense riparian forests (Figure 7). |
| P24 | 4.07 | 7.2 | MC | a | - | - | - | - | Intense rapids stretch with an approximate width of 927 m. Stretches of flooded areas on the left bank and preserved dense riparian forest on both banks. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assumpção, L.d.; Makrakis, M.C.; Silva, J.F.M.d.; Moraes, K.A.S.d.; Pini, S.F.R.; Silva, P.S.d.; Kashiwaqui, E.A.L.; Gentil, E.; Souza-Shibatta, L.; Shibatta, O.A.; et al. Deep Pools: Ecological Sanctuaries for Steindachneridion melanodermatum, a Large Endemic and Endangered Pimelodid of the Iguaçu River. Water 2021, 13, 1700. https://doi.org/10.3390/w13121700
Assumpção Ld, Makrakis MC, Silva JFMd, Moraes KASd, Pini SFR, Silva PSd, Kashiwaqui EAL, Gentil E, Souza-Shibatta L, Shibatta OA, et al. Deep Pools: Ecological Sanctuaries for Steindachneridion melanodermatum, a Large Endemic and Endangered Pimelodid of the Iguaçu River. Water. 2021; 13(12):1700. https://doi.org/10.3390/w13121700
Chicago/Turabian StyleAssumpção, Lucileine de, Maristela Cavicchioli Makrakis, Jhony Ferry Mendonça da Silva, Karane Allison Silvestre de Moraes, Suelen Fernanda Ranucci Pini, Patrícia Sarai da Silva, Elaine Antoniassi Luiz Kashiwaqui, Eduardo Gentil, Lenice Souza-Shibatta, Oscar Akio Shibatta, and et al. 2021. "Deep Pools: Ecological Sanctuaries for Steindachneridion melanodermatum, a Large Endemic and Endangered Pimelodid of the Iguaçu River" Water 13, no. 12: 1700. https://doi.org/10.3390/w13121700
APA StyleAssumpção, L. d., Makrakis, M. C., Silva, J. F. M. d., Moraes, K. A. S. d., Pini, S. F. R., Silva, P. S. d., Kashiwaqui, E. A. L., Gentil, E., Souza-Shibatta, L., Shibatta, O. A., & Makrakis, S. (2021). Deep Pools: Ecological Sanctuaries for Steindachneridion melanodermatum, a Large Endemic and Endangered Pimelodid of the Iguaçu River. Water, 13(12), 1700. https://doi.org/10.3390/w13121700