
Environmental Impacts on Zooplankton Functional Diversity in Brackish Semi-Enclosed Gulf


Abstract
:1. Introduction
2. Materials and Methods
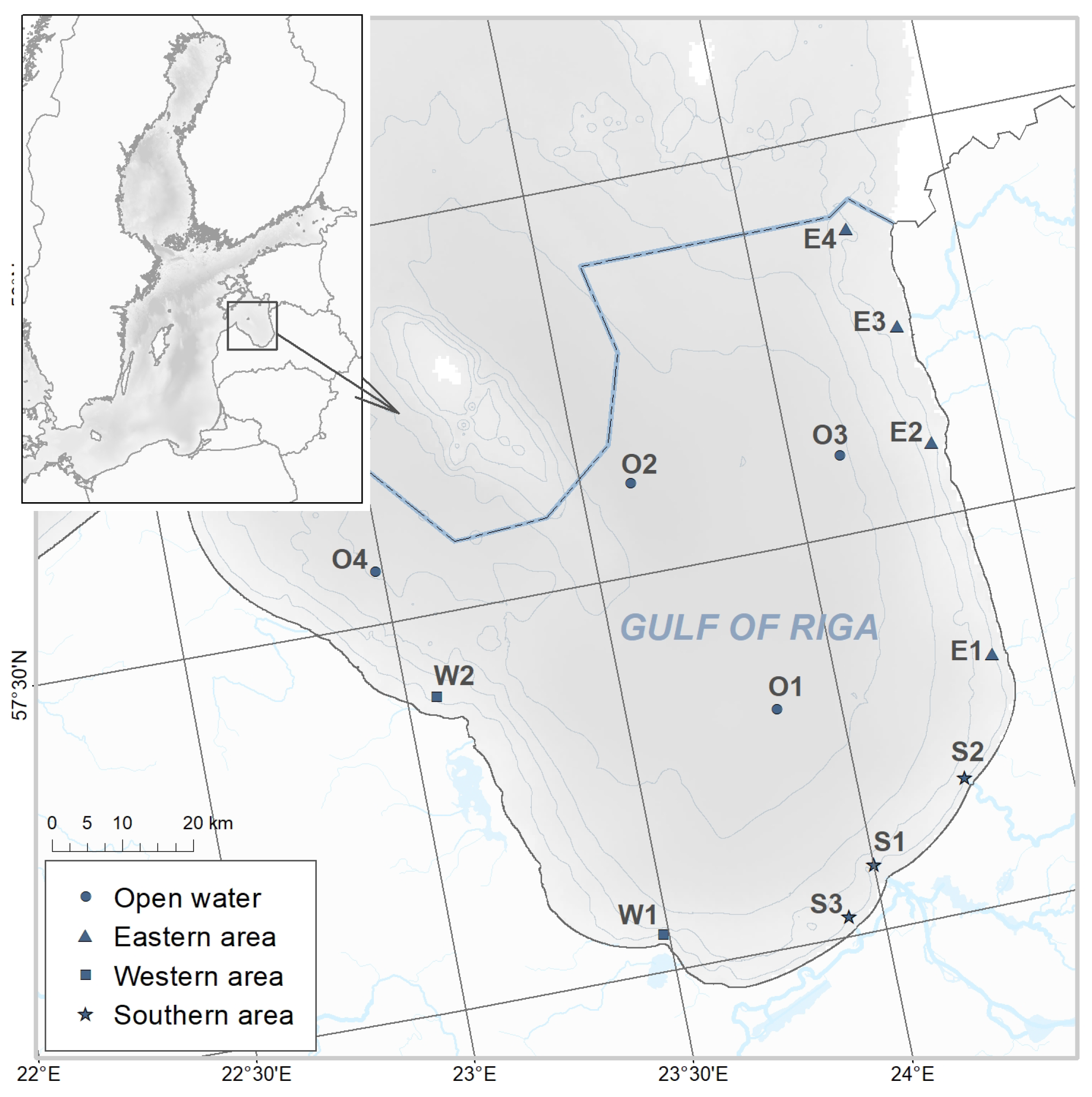
2.1. Study Area and Sampling
2.2. Mesozooplankton Traits and Functional Groups

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Func. Group | Abbr | FM | ML | PS | TG | HS | Dendrogram |
|---|---|---|---|---|---|---|---|
| G1 | KerCoch | F [49] | 1 [50] | 1 [49] | H [49] | 0 |  |
| KerQua | F [49] | 1 [50] | 1 [49] | H [49] | 0 | ||
| KerCruc | F [49] | 1 [50] | 1 [49] | H [49] | 0 | ||
| Amph | F [51] | 1 [41] | 1 [51] | H [51] | 0 | ||
| Poly | F [49] | 1 [52] | 1 [52] | H [52] | 0 | ||
| Biv | F [53] | 1 [54] | 1 [53] | H [54] | 0 | ||
| Bos | F [55] | 1 [41] | 1 [56] | H [55] | 1 | ||
| G2 | Cycl | A [57] | 2 [41] | 2 [56] | C [58] | 0 | |
| G3 | Pleo | M [59,60] | 3 [41] | 2 [59,60] | C [59,60] | 1 | |
| Evad | M [59,60] | 4 [41] | 2 [59,60] | O [59,60] | 1 | ||
| AcSpp | M [57] | 3 [57] | 2 [56] | O [61] | 0 | ||
| AcLon | M [57] | 3 [57] | 2 [56] | O [61] | 0 | ||
| Tem | F [57] | 4 [57] | 2 [56] | O [62] | 0 | ||
| G4 | SySpp | C [49] | 1 [63] | 1 [49] | O [49] | 0 | |
| SyBal | C [49] | 1 [63] | 1 [49] | O [49] | 0 | ||
| CalN | C [64] | 1 [64] | 1 [56] | O [65] | 0 | ||
| CycN | C [64] | 1 [64] | 1 [56] | O [58] | 0 | ||
| G5 | Limn | C [66] | 5 [57] | 3 [56] | C [66] | 2 | |
| Cerc | C [67] | 5 [68] | 3 [56] | C [67] | 1 | ||
| Eury | C [57] | 4 [57] | 2 [56] | O [69] | 2 |
2.3. Functional Diversity Indices
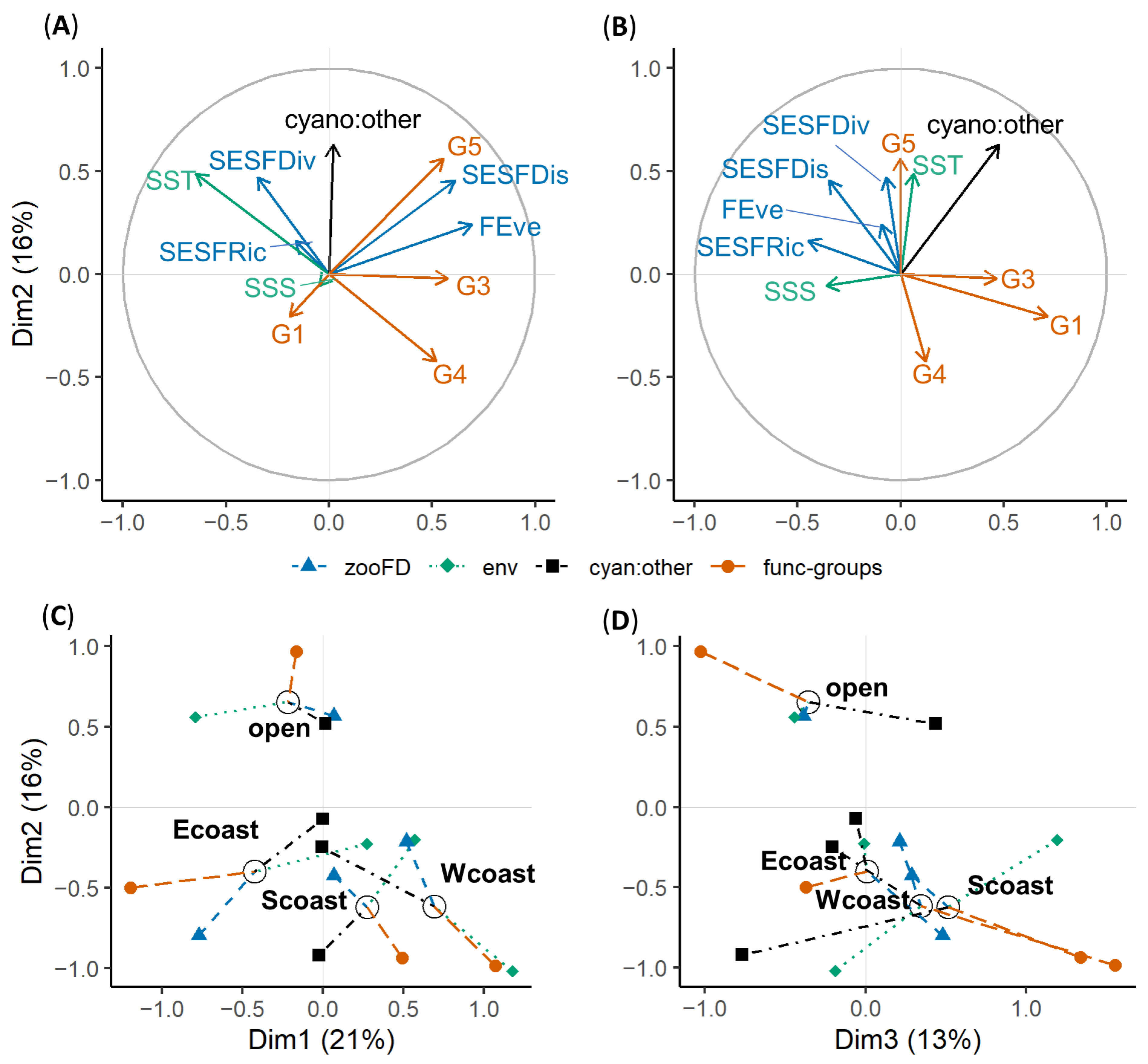
2.4. Data Analysis
3. Results
3.1. Environmental Gradients
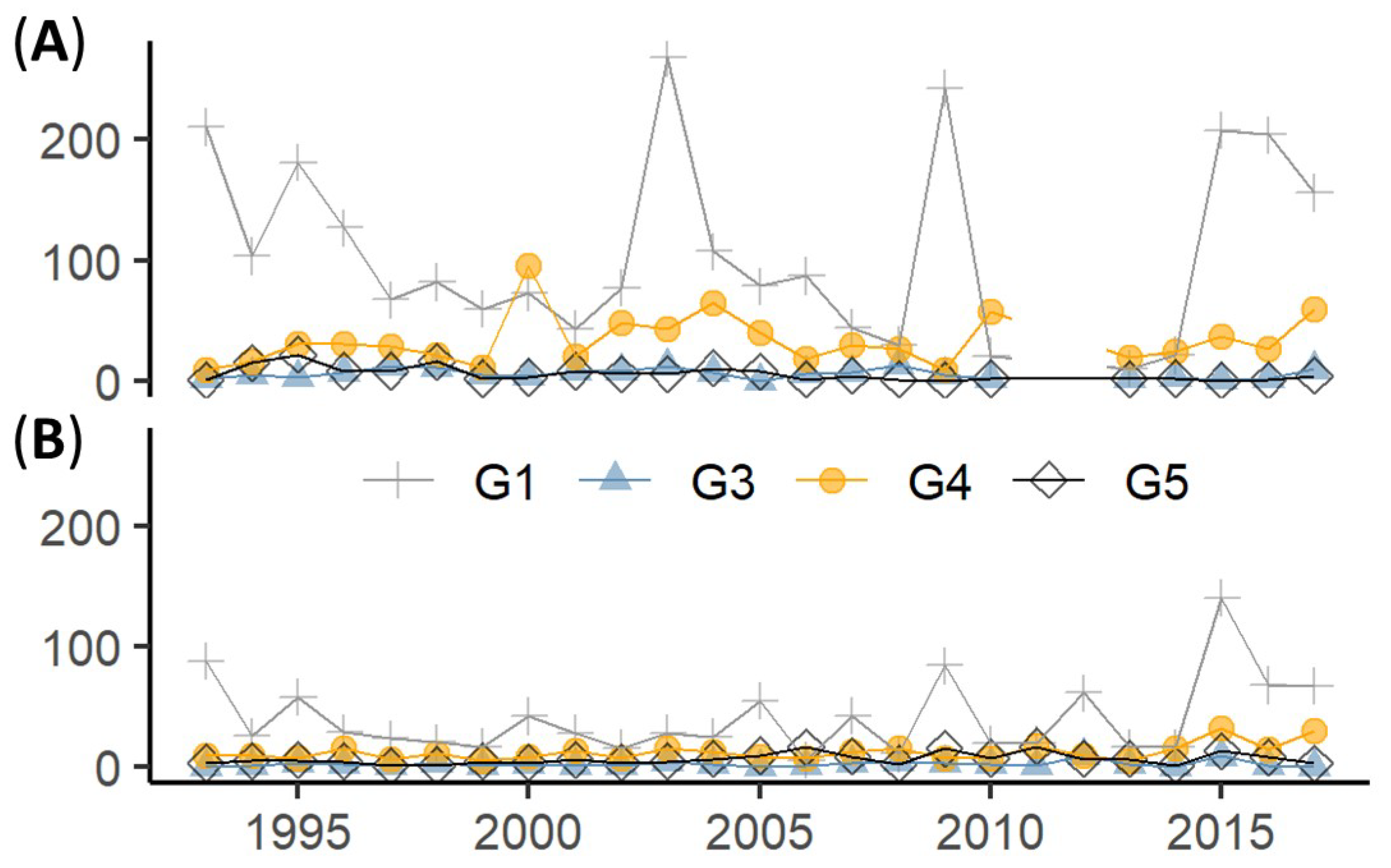
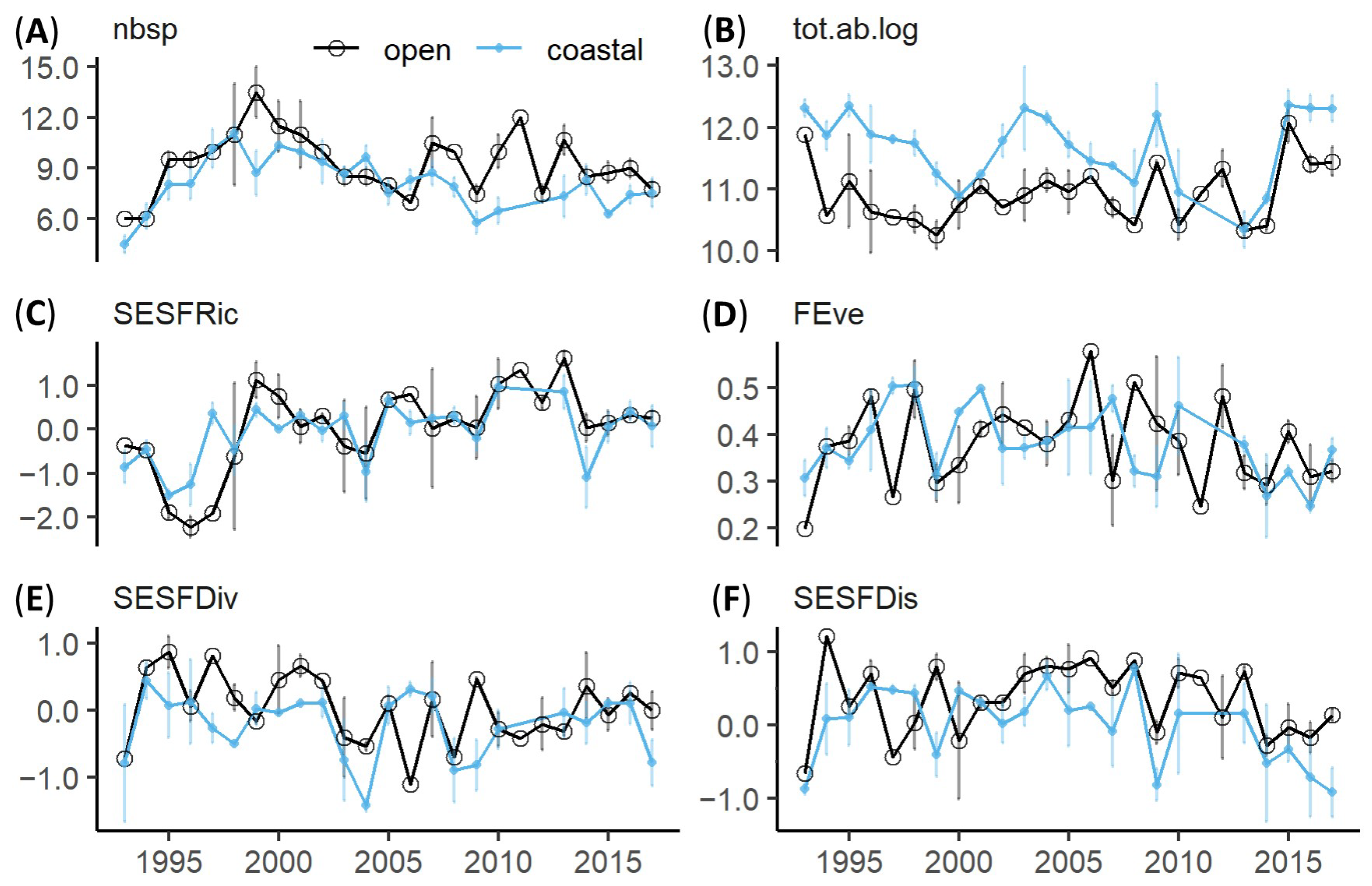
3.2. Long Term Dynamics
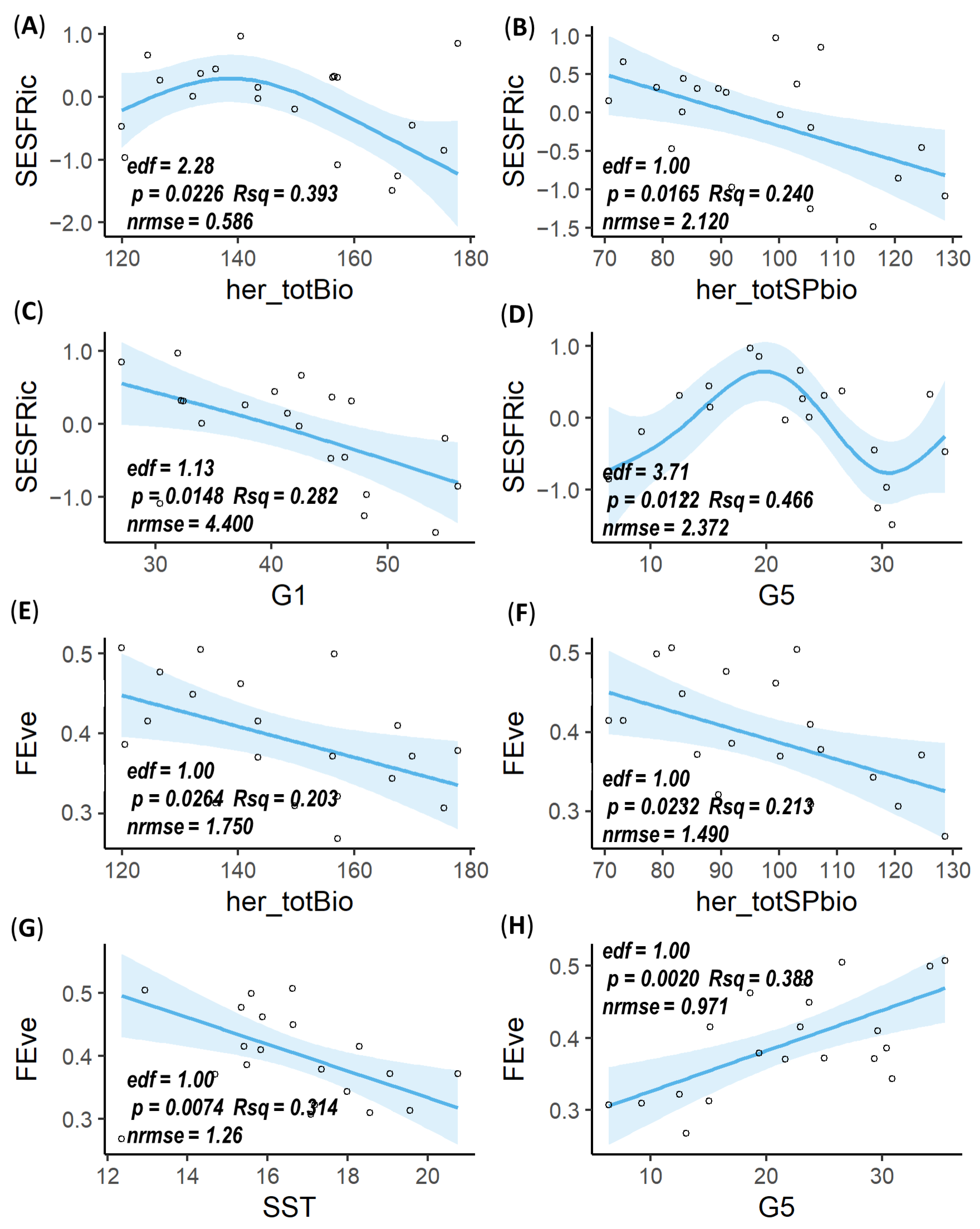
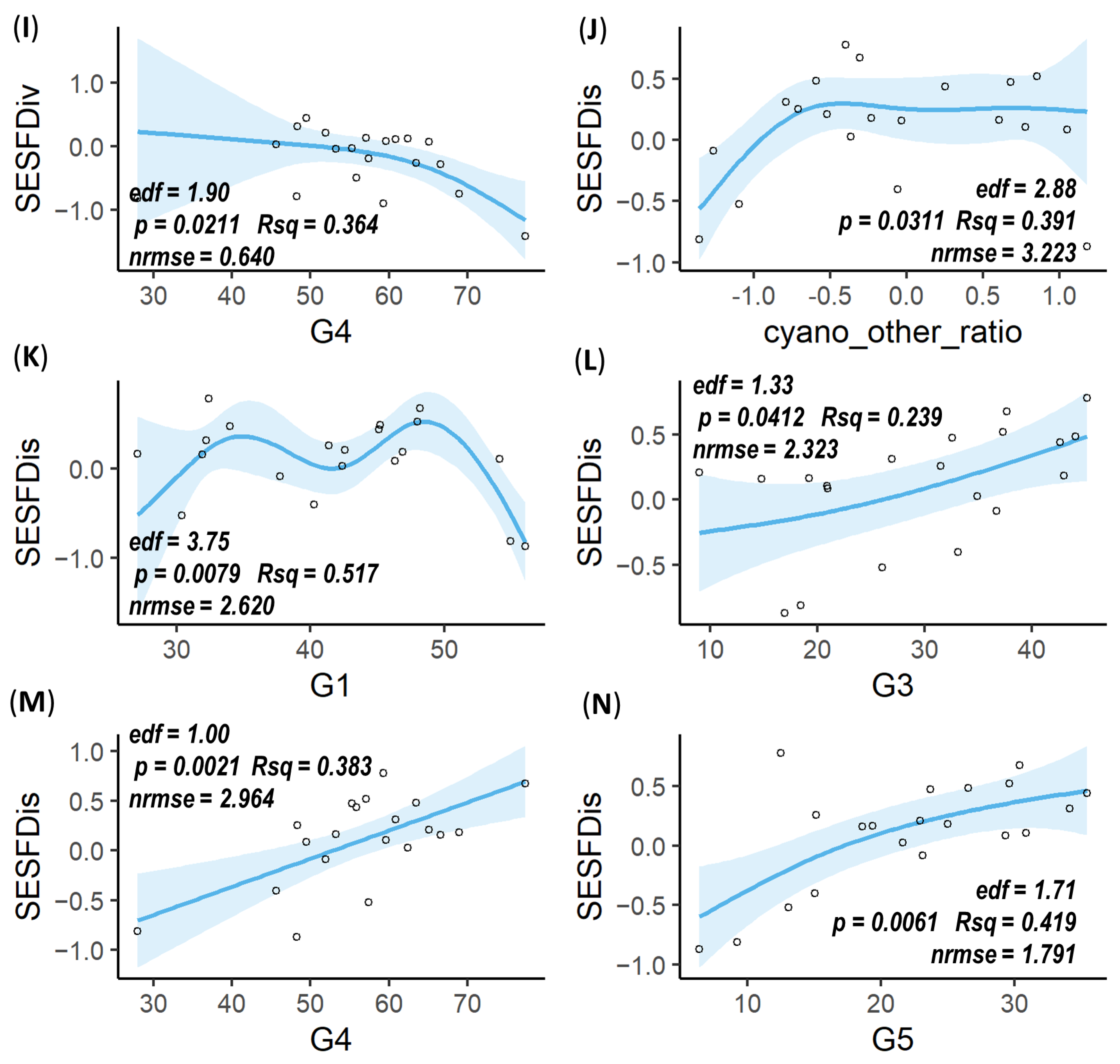
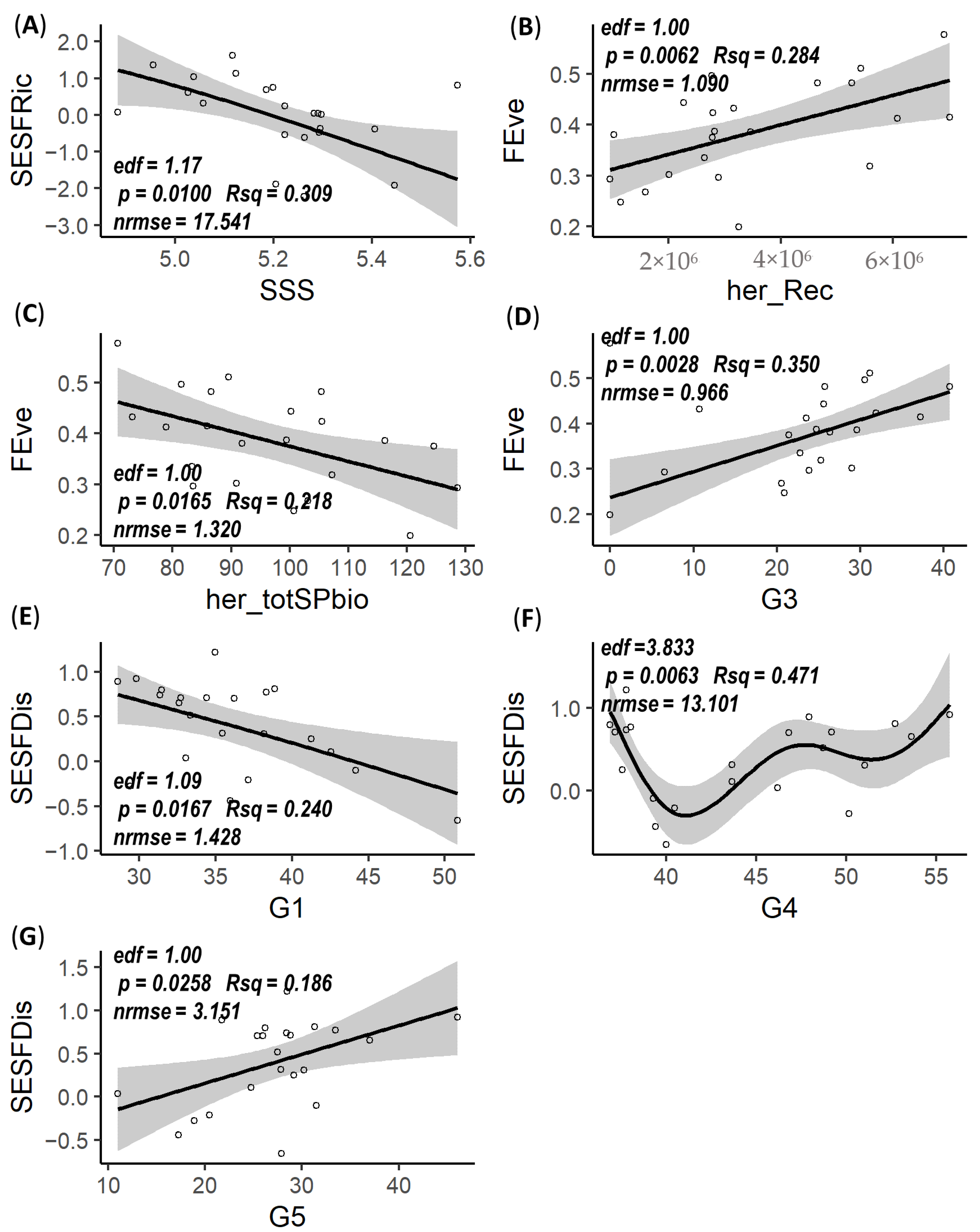
3.3. Mesozooplankton FD Responses to Environmental Factors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chapin, F.S.I.; Reynolds, H.L.; D’Antonio, C.M.; Eckhart, V.M. The functional role of species in terrestrial ecosystems. In Global Change and Terrestrial Ecosystems; Walker, B., Steffen, W., Eds.; Cambridge University Press: Cambridge, UK, 1996; pp. 403–428. [Google Scholar]
- Menge, B.A.; Olson, A.M. Role of scale and environmental factors in regulation of community structure. Trends Ecol. Evol. 1990, 5, 52–57. [Google Scholar] [CrossRef]
- Diaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Functional diversity (FD), species richness and community composition. Ecol. Lett. 2002, 5, 402–411. [Google Scholar] [CrossRef]
- Mason, N.W.; de Bello, F.; Mouillot, D.; Pavoine, S.; Dray, S. A guide for using functional diversity indices to reveal changes in assembly processes along ecological gradients. J. Veg. Sci. 2013, 24, 794–806. [Google Scholar] [CrossRef]
- Pecuchet, L.; Lindegren, M.; Hidalgo, M.; Delgado, M.; Esteban, A.; Fock, H.O.; Gil de Sola, L.; Punzón, A.; Sólmundsson, J.; Payne, M.R. From traits to life-history strategies: Deconstructing fish community composition across European seas. Glob. Ecol. Biogeogr. 2017, 26, 812–822. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F., III; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Mouillot, D.; Graham, N.A.; Villéger, S.; Mason, N.W.; Bellwood, D.R. A functional approach reveals community responses to disturbances. Trends Ecol. Evol. 2013, 28, 167–177. [Google Scholar] [CrossRef]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime shifts, resilience, and biodiversity in ecosystem management. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar] [CrossRef] [Green Version]
- Vallina, S.M.; Cermeno, P.; Dutkiewicz, S.; Loreau, M.; Montoya, J.M. Phytoplankton functional diversity increases ecosystem productivity and stability. Ecol. Model. 2017, 361, 184–196. [Google Scholar] [CrossRef]
- Litchman, E.; Ohman, M.D.; Kiørboe, T. Trait-based approaches to zooplankton communities. J. Plankton Res. 2013, 35, 473–484. [Google Scholar] [CrossRef] [Green Version]
- Hébert, M.P.; Beisner, B.E.; Maranger, R. A meta-analysis of zooplankton functional traits influencing ecosystem function. Ecology 2016, 97, 1069–1080. [Google Scholar] [CrossRef]
- Gomes, L.F.; Pereira, H.R.; Gomes, A.C.A.M.; Vieira, M.C.; Martins, P.R.; Roitman, I.; Vieira, L.C.G. Zooplankton functional-approach studies in continental aquatic environments: A systematic review. Aquat. Ecol. 2019, 8, 191–203. [Google Scholar] [CrossRef]
- Helenius, L.K.; Leskinen, E.; Lehtonen, H.; Nurminen, L. Spatial patterns of littoral zooplankton assemblages along a salinity gradient in a brackish sea: A functional diversity perspective. Estuar. Coast. Shelf Sci. 2017, 198, 400–412. [Google Scholar] [CrossRef]
- Jansson, A.; Klais-Peets, R.; Grinienė, E.; Rubene, G.; Semenova, A.; Lewandowska, A.; Engström-Öst, J. Functional shifts in estuarine zooplankton in response to climate variability. Ecol. Evol. 2020, 10, 11591–11606. [Google Scholar] [CrossRef] [PubMed]
- Lokko, K.; Virro, T.; Kotta, J. Seasonal variability in the structure and functional diversity of psammic rotifer communities: Role of environmental parameters. Hydrobiologia 2017, 796, 287–307. [Google Scholar] [CrossRef]
- Pecuchet, L.; Lindegren, M.; Kortsch, S.; Całkiewicz, J.; Jurgensone, I.; Margonski, P.; Otto, S.A.; Putnis, I.; Strāķe, S.; Nordström, M.C. Spatio-temporal dynamics of multi-trophic communities reveal ecosystem-wide functional reorganization. Ecography 2020, 43, 197–208. [Google Scholar] [CrossRef]
- Ojaveer, H.; Jaanus, A.; Mackenzie, B.R.; Martin, G.; Olenin, S.; Radziejewska, T.; Telesh, I.; Zettler, M.L.; Zaiko, A. Status of biodiversity in the Baltic Sea. PLoS ONE 2010, 5, e12467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snoeijs-Leijonmalm, P.; Andrén, E. Why is the Baltic Sea so special to live in. In Biological Oceanography of the Baltic Sea; Snoeijs-Leijonmalm, P., Schubert, H., Radziejewska, T., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 23–84. [Google Scholar]
- Andersen, J.H.; Axe, P.; Backer, H.; Carstensen, J.; Claussen, U.; Fleming-Lehtinen, V.; Järvinen, M.; Kaartokallio, H.; Knuuttila, S.; Korpinen, S.; et al. Getting the measure of eutrophication in the Baltic Sea: Towards improved assessment principles and methods. Biogeochemistry 2011, 106, 137–156. [Google Scholar] [CrossRef] [Green Version]
- Andrushaitis, A. River load of eutrophying substances and heavy metals into the Gulf of Riga. In Ecosystem of the Gulf of Riga between 1920 and 1990; Ojaveer, E., Ed.; Estonian Academy Publishers: Tallinn, Estonia, 1995; pp. 32–40. [Google Scholar]
- Ojaveer, E.; Lumberg, A.; Ojaveer, H. Highlights of zooplankton dynamics in Estonian waters (Baltic Sea). ICES J. Mar. Sci. 1998, 55, 748–755. [Google Scholar] [CrossRef] [Green Version]
- Ikauniece, A. Long-term abundance dynamics of coastal zooplankton in the Gulf of Riga. Environ. Int. 2001, 26, 175–181. [Google Scholar] [CrossRef]
- Kotta, J.; Kotta, I.; Simm, M.; Põllupüü, M. Separate and interactive effects of eutrophication and climate variables on the ecosystem elements of the Gulf of Riga. Estuar. Coast. Shelf Sci. 2009, 84, 509–518. [Google Scholar] [CrossRef]
- Labuce, A.; Dimante-Deimantoviča, I.; Tunēns, J.; Strāķe, S. Zooplankton indicator-based assessment in relation to site location and abiotic factors: A case study from the Gulf of Riga. Environ. Monit. Assess. 2020, 192, 147. [Google Scholar] [CrossRef]
- Livdāne, L.; Putnis, I.; Rubene, G.; Elferts, D.; Ikauniece, A. Baltic herring prey selectively on older copepodites of Eurytemora affinis and Limnocalanus macrurus in the Gulf of Riga. Oceanologia 2016, 58, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Calbet, A. The trophic roles of microzooplankton in marine systems. ICES J. Mar. Sci. 2008, 65, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.; Gorokhova, E.; Larsson, U.L.F. Annual variability in ciliate community structure, potential prey and predators in the open northern Baltic Sea proper. J. Plankton Res. 2004, 26, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Motwani, N.H.; Duberg, J.; Svedén, J.B.; Gorokhova, E. Grazing on cyanobacteria and transfer of diazotrophic nitrogen to zooplankton in the Baltic Sea. Limnol. Oceanogr. 2018, 63, 672–686. [Google Scholar] [CrossRef]
- Snoeijs-Leijonmalm, P. Patterns of biodiversity. In Biological Oceanography of the Baltic Sea; Snoeijs-Leijonmalm, P., Schubert, H., Radziejewska, T., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 123–191. [Google Scholar]
- Donali, E.; Olli, K.; Heiskanen, A.S.; Andersen, T. Carbon flow patterns in the planktonic food web of the Gulf of Riga, the Baltic Sea: A reconstruction by the inverse method. J. Mar. Syst. 1999, 23, 251–268. [Google Scholar] [CrossRef]
- Filstrup, C.T.; Hillebrand, H.; Heathcote, A.J.; Harpole, W.S.; Downing, J.A. Cyanobacteria dominance influences resource use efficiency and community turnover in phytoplankton and zooplankton communities. Ecol. Lett. 2014, 17, 464–474. [Google Scholar] [CrossRef]
- Hogfors, H.; Motwani, N.H.; Hajdu, S.; El-Shehawy, R.; Holmborn, T.; Vehmaa, A.; Engström-Öst, J.; Brutemark, A.; Gorokhova, E. Bloom-forming cyanobacteria support copepod reproduction and development in the Baltic Sea. PLoS ONE 2014, 9, e112692. [Google Scholar] [CrossRef] [Green Version]
- Kornilovs, G.; Möllmann, C.; Sidrevics, L.; Berzinsh, V. Fish predation modified climate-induced long-term trends of mesozooplankton in a semi-enclosed coastal gulf. ICES CM 2004, 50, 13. [Google Scholar]
- Simm, M.; Ojaveer, E. Dynamics of copepods and fish larvae in Pärnu Bay (NE part of the Gulf of Riga) in the spring–summer period. Proc. Est. Acad. Sci. Biol. Ecol. 2000, 49, 317–326. [Google Scholar]
- Berzinsh, V. Hydrology. In Ecosystem of the Gulf of Riga between 1920 and 1990; Ojaveer, E., Ed.; Estonian Academy Publishers: Tallinn, Estonia, 1995; pp. 7–32. [Google Scholar]
- Skudra, M.; Lips, U. Characteristics and inter-annual changes in temperature, salinity and density distribution in the Gulf of Riga. Oceanologia 2017, 59, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Jurgensone, I.; Carstensen, J.; Ikauniece, A.; Kalveka, B. Long-term Changes and Controlling Factors of Phytoplankton Community in the Gulf of Riga (Baltic Sea). Estuar. Coast. 2011, 34, 1205–1219. [Google Scholar] [CrossRef]
- HELCOM. Manual for Marine Monitoring in the COMBINE Programme of HELCOM. Available online: https://helcom.fi/media/publications/Manual-for-Marine-Monitoring-in-the-COMBINE-Programme-of-HELCOM.pdf (accessed on 8 June 2021).
- Telesh, I.; Heerkloss, R. Atlas of Estuarine Zooplankton of the Southern and Eastern Baltic Sea. Part I: Rotifera; Verlag Dr. Kovač: Hamburg, Germany, 2002; pp. 1–90. [Google Scholar]
- Telesh, I.; Heerkloss, R. Atlas of Estuarine Zooplankton of the Southern and Eastern Baltic Sea. Part II: Crustacea; Verlag Dr. Kovač: Hamburg, Germany, 2004; pp. 1–118. [Google Scholar]
- ICES ID Leaflets for Plankton. 2021. Available online: https://www.ices.dk/publications/library/Pages/default.aspx#Default=%7B%22k%22%3A%22%22%2C%22r%22%3A%5B%7B%22n%22%3A%22owstaxIdPublicationType%22%2C%22t%22%3A%5B%22%5C%22%C7%82%C7%824c307c233061343963306361392d643065372d343661662d393464312d3131363861633234653032357c4944204c6561666c657473%5C%22%22%5D%2C%22o%22%3A%22and%22%2C%22k%22%3Afalse%2C%22m%22%3Anull%7D%2C%7B%22n%22%3A%22ReportAcronymOWSCHCS%22%2C%22t%22%3A%5B%22%5C%22%C7%82%C7%8257475a45%5C%22%22%5D%2C%22o%22%3A%22and%22%2C%22k%22%3Afalse%2C%22m%22%3Anull%7D%5D%7D (accessed on 30 June 2021).
- ICES. Baltic Fisheries Assessment Working Group (WGBFAS), 6–13 April 2018, ICES HQ, Copenhagen, Denmark. 2018. Available online: https://www.ices.dk/sites/pub/Publication%20Reports/Expert%20Group%20Report/acom/2018/WGBFAS/01%20WGBFAS%20Report%202018.pdf (accessed on 8 June 2021).
- Wilke, T.; Ahlrichs, W.H.; Bininda-Emonds, O.R. Is the valid species Synchaeta monopus Plate, 1889 (Rotifera: Monogononta) a product of preparation artefacts? J. Nat. Hist. 2019, 53, 413–423. [Google Scholar] [CrossRef]
- Hébert, M.P.; Beisner, B.E. Functional Trait Approaches for the Study of Metazooplankton Ecology. In Zooplankton Ecology; Teodósio, M.A., Barbosa, A.B., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2020; pp. 3–27. [Google Scholar]
- Kiørboe, T. How zooplankton feed: Mechanisms, traits and trade-offs. Biol. Rev. 2011, 86, 311–339. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Hennig, C. fpc: Flexible Procedures for Clustering, R Package Version 2.2-8; 2020. Available online: https://cran.r-project.org/web/packages/fpc/index.html (accessed on 6 July 2021).
- Ruttner-Kolisko, A. Plankton Rotifers. Biology and Taxonomy; E.Schweizerbart’sche Verlagsbuchhandlung (Nägele u. Obermiller): Stuttgart, Germany, 1974; pp. 1–146. [Google Scholar]
- Berzins, B. Rotatoria (III): Monogonata: Ploima: Brachionidae: Keratella. Available online: https://doi.org/10.17895/ices.pub.5011 (accessed on 8 June 2021).
- Pansch, C.; Schlegel, P.; Havenhand, J. Larval development of the barnacle Amphibalanus improvisus responds variably but robustly to near-future ocean acidification. ICES J. Mar. Sci. 2013, 70, 805–811. [Google Scholar] [CrossRef] [Green Version]
- Burckhardt, R.; Schumann, R.; Bochert, R. Feeding biology of the pelagic larvae of Marenzelleria cf. viridis (Polychaeta: Spionidae) from the Baltic Sea. Aquat. Ecol. 1997, 31, 149–162. [Google Scholar]
- Arapov, J.; Ezgeta-Balić, D.; Peharda, M.; Ninčević Gladan, Ž. Bivalve feeding—How and what they eat? Croat. J. Fish. 2010, 68, 105–116. [Google Scholar]
- Raby, D.; Lagadeuc, Y.; Dodson, J.J.; Mingelbier, M. Relationship between feeding and vertical distribution of bivalve larvae in stratified and mixed waters. Mar. Ecol. Prog. Ser. 1994, 103, 275–284. [Google Scholar] [CrossRef]
- Sommer, U.; Sommer, F. Cladocerans versus copepods: The cause of contrasting top–down controls on freshwater and marine phytoplankton. Oecologia 2006, 147, 183–194. [Google Scholar] [CrossRef]
- Hansen, B.; Bjornsen, P.K.; Hansen, P.J. The size ratio between planktonic predators and their prey. Limnol. Oceanogr. 1994, 39, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Brun, P.; Payne, M.R.; Kiørboe, T. A trait database for marine copepods. Earth Syst. Scit. Data 2017, 9, 99–113. [Google Scholar] [CrossRef] [Green Version]
- García, C.E.; Nandini, S.; Sarma, S. Demographic characteristics of the copepod Acanthocyclops americanus (Sars, 1863) (Copepoda: Cyclopoida) fed mixed algal (Scenedesmus acutus)-rotifer (Brachionus havanaensis) diet. Hydrobiologia 2011, 666, 59–69. [Google Scholar] [CrossRef]
- Egloff, D.A.; Fofonoff, P.W.; Onbé, T. Reproductive biology of marine cladocerans. Adv. Mar. Biol. 1997, 31, 79–167. [Google Scholar]
- Katechakis, A.; Stibor, H. Feeding selectivities of the marine cladocerans Penilia avirostris, Podon intermedius and Evadne nordmanni. Mar. Biol. 2004, 145, 529–539. [Google Scholar] [CrossRef]
- Tiselius, P. Contribution of aloricate ciliates to the diet of Acartia clausi and Centropages hamatus in coastal waters. Mar. Ecol. Prog. Ser. 1989, 56, 49–56. [Google Scholar] [CrossRef]
- Gentsch, E.; Kreibich, T.; Hagen, W.; Niehoff, B. Dietary shifts in the copepod Temora longicornis during spring: Evidence from stable isotope signatures, fatty acid biomarkers and feeding experiments. J. Plankton Res. 2009, 31, 45–60. [Google Scholar] [CrossRef] [Green Version]
- Hollowday, E. Family Synchaetidae Hudson & Gosse, 186. In Guides to the Identification of the Microinvertebrates. ROTIFERA; Nograndy, T., Segers, H., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2002; Volume 6, pp. 87–264. [Google Scholar]
- Titelman, J.; Kiørboe, T. Motility of copepod nauplii and implications for food encounter. Mar. Ecol. Prog. Ser. 2003, 247, 123–135. [Google Scholar] [CrossRef] [Green Version]
- Stoecker, D.K.; Egloff, D.A. Predation by Acartia tonsa Dana on planktonic ciliates and rotifers. J. Exp. Mar. Biol. Ecol. 1987, 110, 53–68. [Google Scholar] [CrossRef]
- Warren, G.J. Predaceous feeding habits of Limnocalanus macrurus. J. Plankton Res. 1985, 7, 537–552. [Google Scholar] [CrossRef]
- Rivier, I.K. The predatory Cladocera (Onychopoda: Podonidae, Polyphemidae, Cercopagidae) and Leptodorida of the World. In Guides to the Identification of the Micro-Invertebrates of the Continental Waters of the World; Dumont, H., Ed.; SPB Academic Pub: The Hague, The Netherlands, 1998; pp. 1–213. [Google Scholar]
- Aladin, N.V.; Panov, V.E. Comparative biology of the predatory cladoceran Cercopagis pengoi from Lake Ontario, Baltic Sea and Caspian Sea. Arch. Hydrobiol. 2000, 1, 23–50. [Google Scholar]
- Revis, N.; Castel, J.; Tackx, M. Some reflections on the structure of the mandibleplate of Eurytemora affinis (Copepoda, Calanoida). Hydrobiol. Bull. 1991, 25, 45–50. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology, R Package Version 1.0-12; 2014. Available online: https://rdrr.io/cran/FD/ (accessed on 6 July 2021).
- Mason, N.; Mouillot, D.; Lee, W.; Wilson, J. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 1, 112–118. [Google Scholar] [CrossRef]
- Rao, C.R. Diversity and dissimilarity coefficients: A unified approach. Theor. Popul. Biol. 1982, 21, 24–43. [Google Scholar] [CrossRef]
- Swenson, N.G. Functional and Phylogenetic Ecology in R; Springer: New York, NY, USA, 2014; pp. 1–209. [Google Scholar]
- Kembel, S.; Cowan, P.; Helmus, M.; Cornwell, W.; Morlon, H.; Ackerly, D.; Blomberg, S.; Webb, C. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: A Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Peterson, R.A.; Cavanaugh, J.E. Ordered quantile normalization: A semiparametric transformation built for the cross-validation era. J. Appl. Stat. 2020, 47, 2312–2327. [Google Scholar] [CrossRef]
- Otto, S.A.; Plonus, R.; Funk, S.; Keth, A. INDperform: Evaluation of Indicator Performances for Assessing Ecosystem States, R Package Version 0.2.2.9000; 2020. Available online: https://rdrr.io/cran/INDperform/ (accessed on 6 July 2021).
- Gasiūnaitė, Z.R. Coupling of the limnetic and brackishwater plankton crustaceans in the Curonian Lagoon (Baltic Sea). Int. Rev. Hydrobiol. 2000, 85, 653–661. [Google Scholar] [CrossRef]
- Telesh, I.V. Plankton of the Baltic estuarine ecosystems with emphasis on Neva Estuary: A review of present knowledge and research perspectives. Mar. Pollut. Bull. 2004, 49, 206–219. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, D.M. The disturbing history of intermediate disturbance. Oikos 1999, 84, 145–147. [Google Scholar] [CrossRef] [Green Version]
- Lips, U.; Zhurbas, V.; Skudra, M.; Väli, G. A numerical study of circulation in the Gulf of Riga, Baltic Sea. Part I: Whole-basin gyres and mean currents. Cont. Shelf Res. 2016, 112, 1–13. [Google Scholar] [CrossRef]
- Forster, J.; Hirst, A.G. The temperature-size rule emerges from ontogenetic differences between growth and development rates. Funct. Ecol. 2012, 26, 483–492. [Google Scholar] [CrossRef]
- Kenitz, K.M.; Visser, A.W.; Mariani, P.; Andersen, K.H. Seasonal succession in zooplankton feeding traits reveals trophic trait coupling. Limnol. Oceanogr. 2017, 62, 1184–1197. [Google Scholar] [CrossRef] [Green Version]
- Kiørboe, T.; Visser, A.; Andersen, K.H. A trait-based approach to ocean ecology. ICES J. Mar. Sci. 2018, 75, 1849–1863. [Google Scholar] [CrossRef]
- McManus, M.A.; Woodson, C.B. Plankton distribution and ocean dispersal. J. Exp. Biol. 2012, 215, 1008–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahru, M.; Nômmann, S.; Simm, M.; Vilbaste, K. Plankton Distributions and Processes in the Baltic Boundary Zones. In Marine Interfaces Ecohydrodynamics; Nihoul, J., Ed.; Elsevier: Oxford, UK, 1986; pp. 273–294. [Google Scholar]
- Winder, M.; Varpe, Ø. Interactions in Plankton Food Webs: Seasonal Succession and Phenology of Baltic Sea Zooplankton. In Zooplankton Ecology; Teodósio, M.A., Barbosa, A.B., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2020; pp. 162–191. [Google Scholar]
- Viitasalo, M.; Katajisto, T. Mesozooplankton resting eggs in the Baltic Sea: Identification and vertical distribution in laminated and mixed sediments. Mar. Biol. 1994, 120, 455–466. [Google Scholar] [CrossRef]
- Carman, R.; Aigars, J.; Larsen, B. Carbon and nutrient geochemistry of the surface sediments of the Gulf of Riga, Baltic Sea. Mar. Geol. 1996, 134, 57–76. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Harmful cyanobacterial blooms: Causes, consequences, and controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef]
- Tillmanns, A.R.; Wilson, A.E.; Pick, F.R.; Sarnelle, O. Meta-analysis of cyanobacterial effects on zooplankton population growth rate: Species-specific responses. Fund. Appl. Limnol. 2008, 171, 285–295. [Google Scholar] [CrossRef]
- Finkel, Z.V.; Beardall, J.; Flynn, K.J.; Quigg, A.; Rees, T.A.V.; Raven, J.a. Phytoplankton in a changing world: Cell size and elemental stoichiometry. J. Plankton Res. 2010, 32, 119–137. [Google Scholar] [CrossRef] [Green Version]
- Jónasdóttir, S. Fatty Acid Profiles and Production in Marine Phytoplankton. Mar. Drugs 2019, 17, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wejnerowski, L.; Cerbin, S.; Dziuba, M.K. Thicker filaments of Aphanizomenon gracile are more harmful to Daphnia than thinner Cylindrospermopsis raciborskii. Zool. Stud. 2015, 54, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorokhova, E.; El-Shehawy, R.; Lehtiniemi, M.; Garbaras, A. How Copepods Can Eat Toxins Without Getting Sick: Gut Bacteria Help Zooplankton to Feed in Cyanobacteria Blooms. Front. Microbiol. 2021, 11, 589816. [Google Scholar] [CrossRef]
- Krztoń, W.; Kosiba, J.; Pociecha, A.; Wilk-Woźniak, E. The effect of cyanobacterial blooms on bio- and functional diversity of zooplankton communities. Biodivers Conserv. 2019, 28, 1815–1835. [Google Scholar] [CrossRef] [Green Version]
- Landry, M.R.; Calbet, A. Microzooplankton production in the oceans. ICES J. Mar. Sci. 2004, 61, 501–507. [Google Scholar] [CrossRef] [Green Version]
- Möllmann, C.; Diekmann, R.; Müller-Karulis, B.; Kornilovs, G.; Plikshs, M.; Axe, P. Reorganization of a large marine ecosystem due to atmospheric and anthropogenic pressure: A discontinuous regime shift in the Central Baltic Sea. Glob. Chang. Biol. 2009, 15, 1377–1393. [Google Scholar] [CrossRef]
- Kortsch, S.; Frelat, R.; Pecuchet, L.; Olivier, P.; Putnis, I.; Bonsdorff, E.; Ojaveer, H.; Jurgensone, I.; Strāķe, S.; Rubene, G.; et al. Disentangling temporal food web dynamics facilitates understanding of ecosystem functioning. J. Anim. Ecol. 2021, 90, 1205–1216. [Google Scholar] [CrossRef]
- Ojaveer, H.; Kotta, J.; Põllumäe, A.; Põllupüü, M.; Jaanus, A.; Vetemaa, M. Alien species in a brackish water temperate ecosystem: Annual-scale dynamics in response to environmental variability. Environ. Res. 2011, 111, 933–942. [Google Scholar] [CrossRef]
- Ojaveer, H.; Simm, M.; Lankov, A. Population dynamics and ecological impact of the non-indigenous Cercopagis pengoi in the Gulf of Riga (Baltic Sea). Hydrobiologia 2004, 522, 261–269. [Google Scholar] [CrossRef]
- Gorokhova, E.; Fagerberg, T.; Hansson, S. Predation by herring (Clupea harengus) and sprat (Sprattus sprattus) on Cercopagis pengoi in a western Baltic Sea bay. ICES J. Mar. Sci. 2004, 61, 959–965. [Google Scholar] [CrossRef] [Green Version]
- Gorokhova, E.; Hansson, S.; Höglander, H.; Andersen, C.M. Stable isotopes show food web changes after invasion by the predatory cladoceran Cercopagis pengoi in a Baltic Sea bay. Oecologia 2005, 143, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Ojaveer, E. Ecosystems and Living Resources of the Baltic Sea: Their Assessment and Management; Springer: Cham, Switzerland, 2017; pp. 1–283. [Google Scholar]
- Arula, T.; Kotta, J.; Lankov, A.; Simm, M.; Põlme, S. Diet composition and feeding activity of larval spring-spawning herring: Importance of environmental variability. J. Sea Res. 2012, 68, 33–40. [Google Scholar] [CrossRef]
- Arrhenius, F.; Hansson, S. Food consumption of larval, young and adult herring and sprat in the Baltic Sea. Mar. Ecol. Prog. Ser. 1993, 96, 125. [Google Scholar] [CrossRef]







| Year | O1 | O2 | O3 | O4 | E4 | E3 | E2 | E1 | S2 | S1 | S3 | W1 | W2 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1993 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 0 | 0 |
| 1994 | 7 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 3 | 3 | 3 | 3 | 0 |
| 1995 | 4 | 1 | 1 | 1 | 0 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 0 |
| 1996 | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 |
| 1997 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 2 | 3 | 3 | 3 | 2 | 0 |
| 1998 | 4 | 1 | 0 | 0 | 0 | 0 | 0 | 3 | 3 | 5 | 2 | 3 | 0 |
| 1999 | 6 | 2 | 0 | 0 | 0 | 1 | 0 | 3 | 1 | 6 | 3 | 3 | 0 |
| 2000 | 2 | 3 | 1 | 1 | 1 | 0 | 0 | 3 | 3 | 4 | 3 | 3 | 0 |
| 2001 | 6 | 2 | 1 | 1 | 1 | 0 | 0 | 3 | 3 | 6 | 3 | 3 | 0 |
| 2002 | 6 | 3 | 1 | 1 | 1 | 0 | 0 | 3 | 3 | 7 | 2 | 3 | 1 |
| 2003 | 6 | 3 | 1 | 1 | 1 | 0 | 0 | 3 | 2 | 7 | 2 | 3 | 1 |
| 2004 | 3 | 1 | 1 | 1 | 1 | 0 | 0 | 3 | 1 | 4 | 2 | 2 | 0 |
| 2005 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 |
| 2006 | 2 | 0 | 1 | 1 | 1 | 0 | 0 | 2 | 1 | 2 | 1 | 2 | 0 |
| 2007 | 3 | 1 | 1 | 1 | 1 | 0 | 0 | 3 | 1 | 4 | 2 | 2 | 0 |
| 2008 | 3 | 1 | 1 | 1 | 1 | 0 | 0 | 3 | 1 | 4 | 2 | 2 | 0 |
| 2009 | 3 | 1 | 1 | 1 | 1 | 0 | 0 | 3 | 1 | 4 | 2 | 2 | 0 |
| 2010 | 2 | 2 | 0 | 2 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 |
| 2011 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2012 | 3 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 2013 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0 |
| 2014 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 |
| 2015 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 |
| 2016 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 |
| 2017 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 5 | 1 | 4 |
| Taxonomic Affiliation | Abbreviation | Group |
|---|---|---|
| Bosmina (Eubosmina) coregoni | Bos | Cladocera |
| Cercopagis (Cercopagis) pengoi | Cerc | Cladocera |
| Evadne normanni | Evad | Cladocera |
| Pleopis polyphemoides | Pleo | Cladocera |
| Acartia spp. | AcSpp | Copepoda |
| Acartia longiremis | AcLon | Copepoda |
| Cyclopoida | Cycl | Copepoda |
| Eurytemora affinis affinis | Eury | Copepoda |
| Limnocalanus macrurus | Limn | Copepoda |
| Temora longicornis | Tem | Copepoda |
| Calanoida Copepoda nauplii | CalN | Copepoda |
| Cyclopoida Copepoda nauplii | CylN | Copepoda |
| Keratella cochlearis | KerCoch | Rotifera |
| Keratella quadrata | KerQua | Rotifera |
| Keratella cruciformis | KerCruc | Rotifera |
| Synchaeta monopus (but see [44]) | SySpp | Rotifera |
| Synchaeta baltica | SyBal | Rotifera |
| Amphibalanus larvae | Amph | Meroplankton |
| Bivalvia larvae | Biv | Meroplankton |
| Gastropoda larvae | Gast | Meroplankton |
| Polychaeta larvae | Poly | Meroplankton |
| Parameter | Dim1 | Dim2 | Dim3 | Dim4 | Dim5 |
|---|---|---|---|---|---|
| SESFRic | 1.0 | 1.4 | 12.7 | 4.9 | 45.6 |
| FEve | 19.4 | 3.0 | 0.5 | 0.0 | 4.2 |
| SESFDiv | 4.9 | 11.5 | 0.3 | 0.7 | 27.3 |
| SESFDis | 15.0 | 10.7 | 7.7 | 1.9 | 0.1 |
| SSS | 0.1 | 0.2 | 10.4 | 59.3 | 0.7 |
| SST | 21.1 | 15.5 | 0.3 | 9.6 | 1.5 |
| Cyano:other | 0.0 | 29.6 | 20.7 | 20.6 | 12.4 |
| G1 | 1.5 | 2.2 | 32.6 | 0.4 | 0.3 |
| G3 | 13.5 | 0.0 | 13.8 | 0.2 | 2.3 |
| G4 | 11.0 | 9.4 | 1.0 | 0.0 | 5.0 |
| G5 | 12.5 | 16.6 | 0.0 | 2.2 | 0.6 |
| expl.variance | 20.7 | 16.1 | 13.1 | 12.0 | 8.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labuce, A.; Ikauniece, A.; Jurgensone, I.; Aigars, J. Environmental Impacts on Zooplankton Functional Diversity in Brackish Semi-Enclosed Gulf. Water 2021, 13, 1881. https://doi.org/10.3390/w13141881
Labuce A, Ikauniece A, Jurgensone I, Aigars J. Environmental Impacts on Zooplankton Functional Diversity in Brackish Semi-Enclosed Gulf. Water. 2021; 13(14):1881. https://doi.org/10.3390/w13141881
Chicago/Turabian StyleLabuce, Astra, Anda Ikauniece, Iveta Jurgensone, and Juris Aigars. 2021. "Environmental Impacts on Zooplankton Functional Diversity in Brackish Semi-Enclosed Gulf" Water 13, no. 14: 1881. https://doi.org/10.3390/w13141881
APA StyleLabuce, A., Ikauniece, A., Jurgensone, I., & Aigars, J. (2021). Environmental Impacts on Zooplankton Functional Diversity in Brackish Semi-Enclosed Gulf. Water, 13(14), 1881. https://doi.org/10.3390/w13141881