
Soil-Related Predictors for Distribution Modelling of Four European Crayfish Species



Abstract
:1. Introduction
2. Materials and Methods
2.1. Crayfish Data Collection
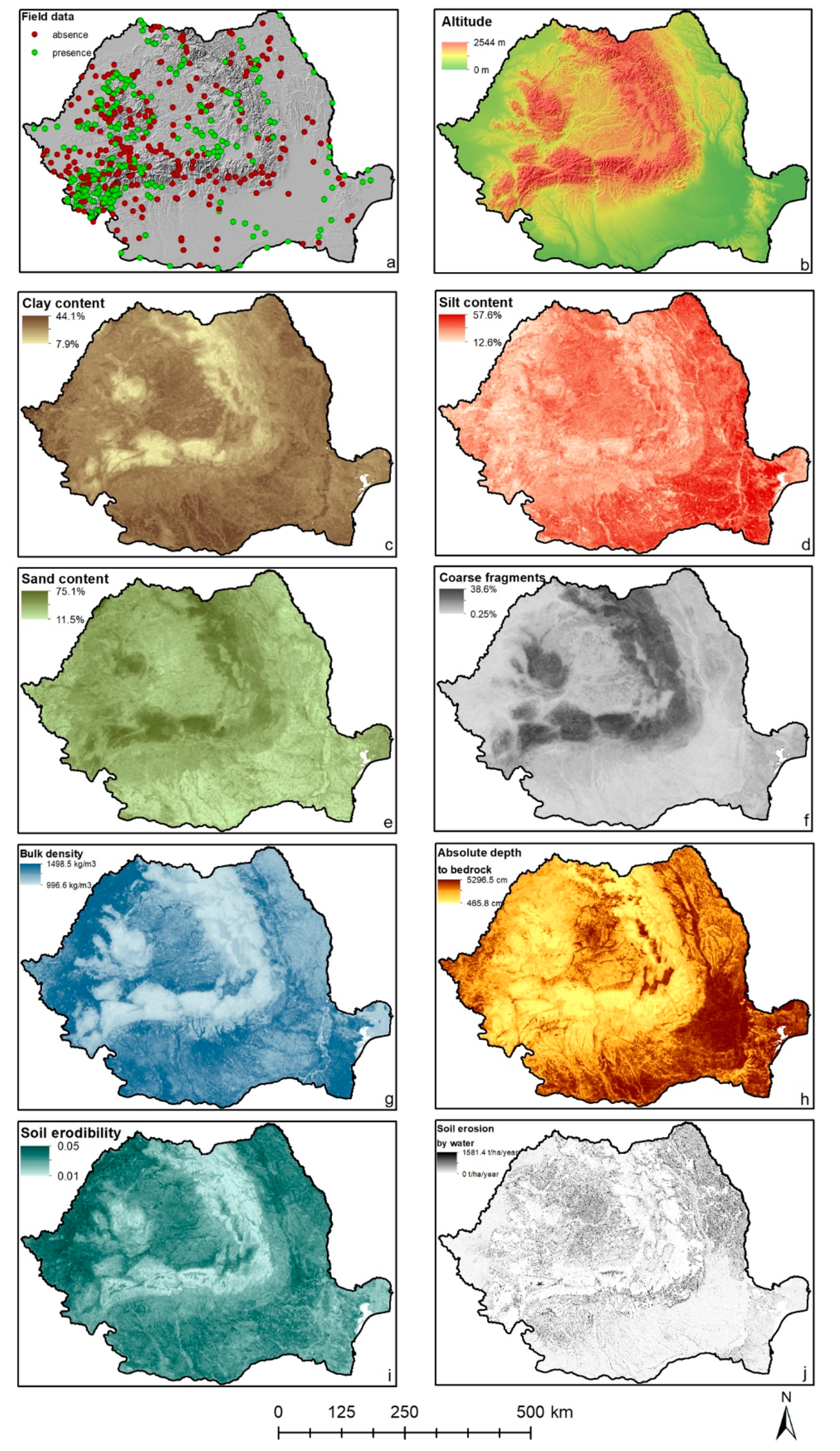
2.2. Soil Data
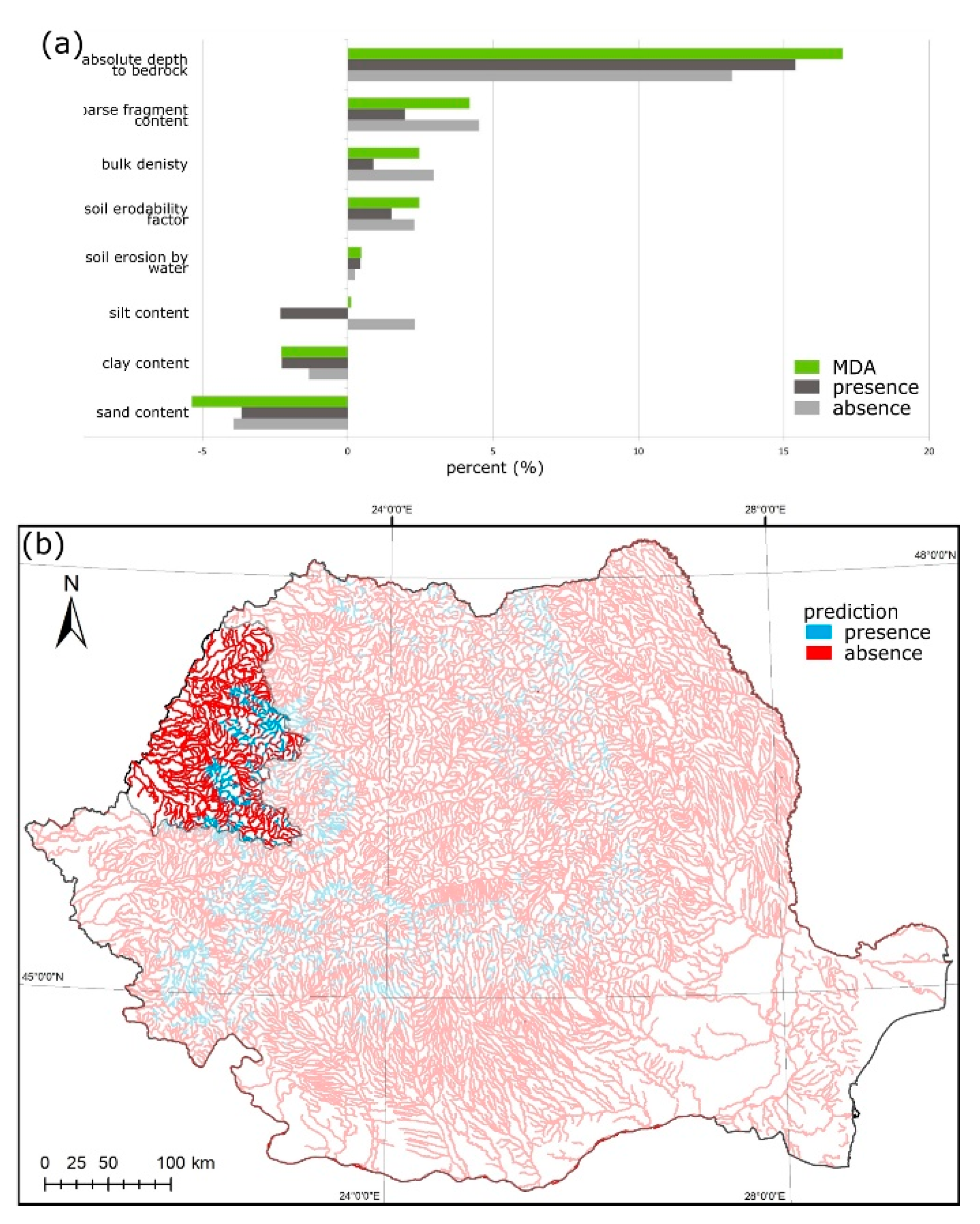
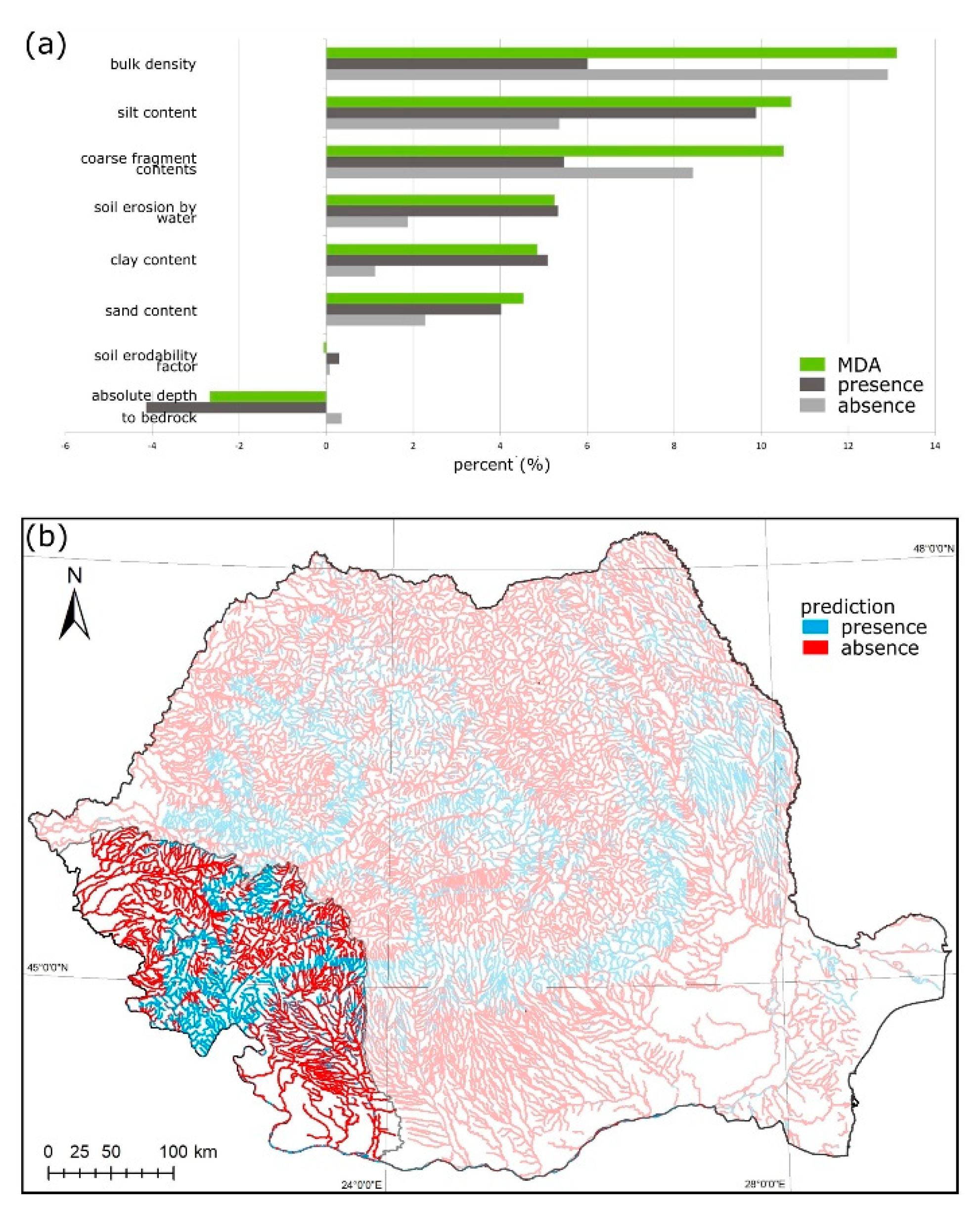
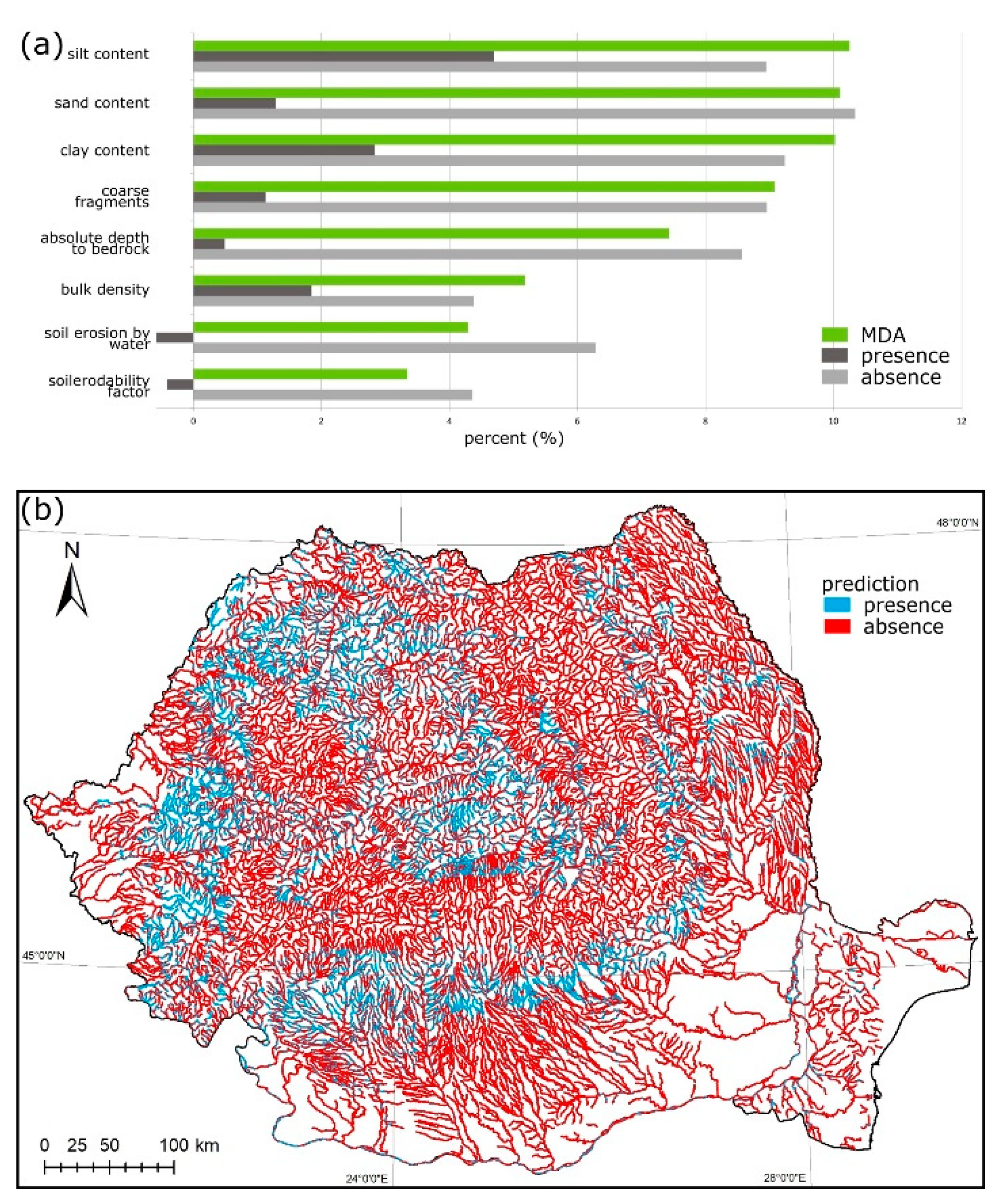
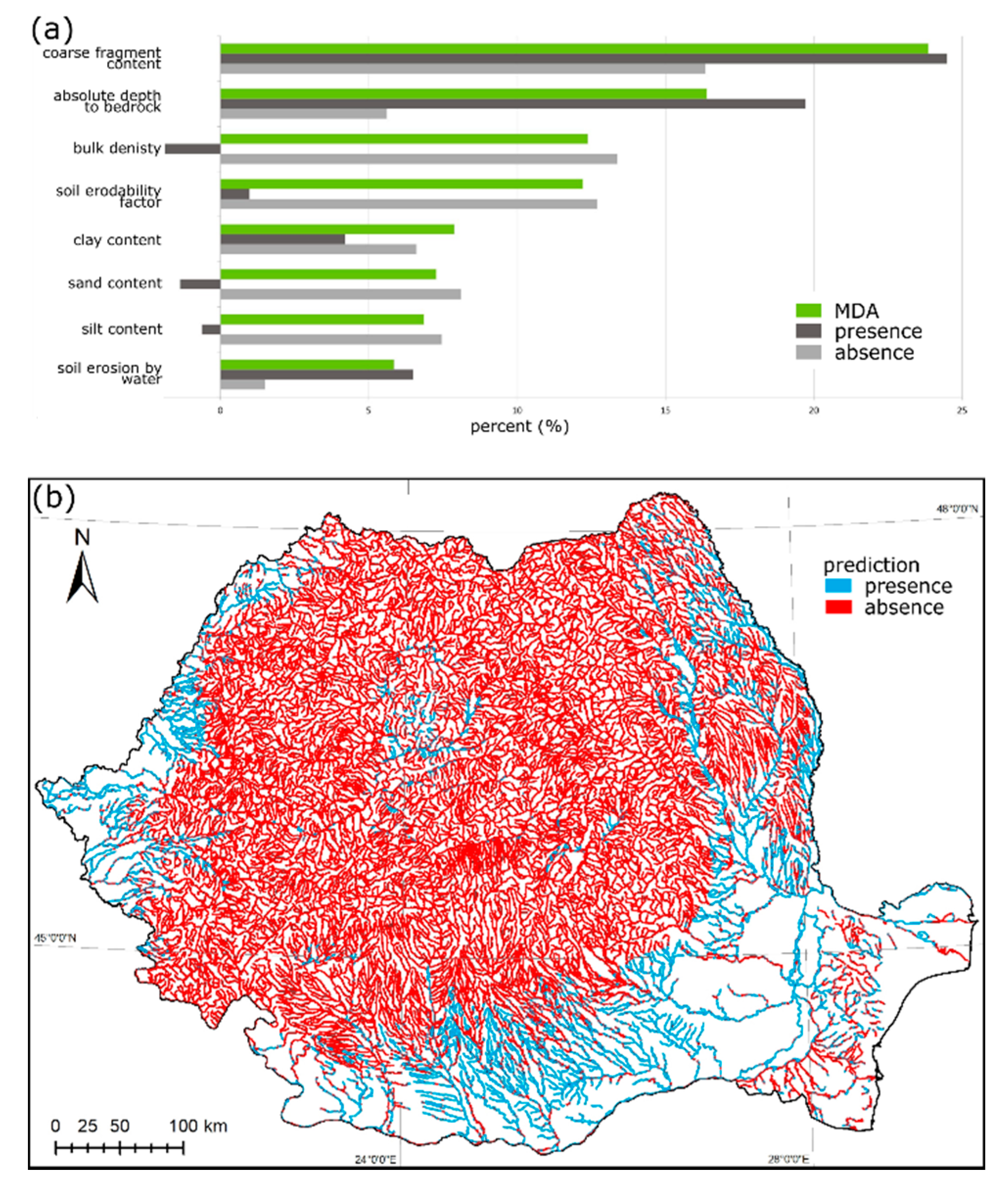
2.3. Variable Importance Analysis
2.4. Species Distribution Maps
2.5. Results Validation
3. Results
3.1. Austropotamobius bihariensis
3.2. Austropotamobius torrentium
3.3. Astacus astacus
3.4. Pontastacus leptodactylus
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chiang, F.; Mazdiyasni, O.; AghaKouchak, A. Evidence of anthropogenic impacts on global drought frequency, duration, and intensity. Nat. Commun. 2021, 12, 2754. [Google Scholar] [CrossRef]
- Demers, A.; Souty-Grosset, C.; Trouilhé, M.C.; Füreder, L.; Renai, B.; Gherardi, F. Tolerance of three European native species of crayfish to hypoxia. Hydrobiologia 2006, 560, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Füreder, L.; Reynolds, J.D. Is Austropotamobius pallipes a good bioindicator*? Bull. Français Pêche Piscic. 2003, 370–371, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Kubec, J.; Veselý, L.; Hossain, M.S.; Buřič, M.; McClain, R.; Kouba, A. High air humidity is sufficient for successful egg incubation and early post-embryonic development in the marbled crayfish (Procambarus virginalis). Freshw. Biol. 2019, 64, 1603–1612. [Google Scholar] [CrossRef] [Green Version]
- Kouba, A.; Tíkal, J.; Císař, P.; Veselý, L.; Fořt, M.; Příborský, J.; Patoka, J.; Buřič, M. The significance of droughts for hyporheic dwellers: Evidence from freshwater crayfish. Sci. Rep. 2016, 6, 26569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streissl, F.; Hödl, W. Habitat and shelter requirements of the stone crayfish, Austropotamobius torrentium Schrank. Hydrobiologia 2002, 477, 195–199. [Google Scholar] [CrossRef]
- Araújo, M.B.; Guisan, A. Five (or so) challenges for species distribution modelling. J. Biogeogr. 2006, 33, 1677–1688. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Modell. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Pârâu, L.G.; Wink, M. Common patterns in the molecular phylogeography of western palearctic birds: A comprehensive review. J. Ornithol. 2021, 1, 1–23. [Google Scholar] [CrossRef]
- Deblauwe, V.; Droissart, V.; Bose, R.; Sonké, B.; Blach-Overgaard, A.; Svenning, J.-C.; Wieringa, J.J.; Ramesh, B.R.; Stévart, T.; Couvreur, T.L.P. Remotely sensed temperature and precipitation data improve species distribution modelling in the tropics. Glob. Ecol. Biogeogr. 2016, 25, 443–454. [Google Scholar] [CrossRef]
- Waltari, E.; Schroeder, R.; McDonald, K.; Anderson, R.P.; Carnaval, A. Bioclimatic variables derived from remote sensing: Assessment and application for species distribution modelling. Methods Ecol. Evol. 2014, 5, 1033–1042. [Google Scholar] [CrossRef]
- Şandric, I.; Satmari, A.; Zaharia, C.; Petrovici, M.; Cîmpean, M.; Battes, K.-P.; David, D.-C.; Pacioglu, O.; Weiperth, A.; Gál, B.; et al. Integrating catchment land cover data to remotely assess freshwater quality: A step forward in heterogeneity analysis of river networks. Aquat. Sci. 2019, 81, 26. [Google Scholar] [CrossRef]
- Domisch, S.; Amatulli, G.; Jetz, W. Near-global freshwater-specific environmental variables for biodiversity analyses in 1 km resolution. Sci. Data 2015, 2, 150073. [Google Scholar] [CrossRef] [Green Version]
- Beck, H.E.; Zimmermann, N.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Wood, E.F. Present and future köppen-geiger climate classification maps at 1-km resolution. Sci. Data 2018, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Bearup, D.; Liao, J. Habitat loss alters effects of intransitive higher-order competition on biodiversity: A new metapopulation framework. Proc. R. Soc. B Biol. Sci. 2020, 287, 20201571. [Google Scholar] [CrossRef]
- Brooks, T.M.; Pimm, S.L.; Akçakaya, H.R.; Buchanan, G.M.; Butchart, S.H.M.; Foden, W.; Hilton-Taylor, C.; Hoffmann, M.; Jenkins, C.N.; Joppa, L.; et al. Measuring Terrestrial Area of Habitat (AOH) and Its Utility for the IUCN Red List. Trends Ecol. Evol. 2019, 34, 977–986. [Google Scholar] [CrossRef] [Green Version]
- Crandall, K.A.; De Grave, S. An updated classification of the freshwater crayfishes (Decapoda: Astacidea) of the world, with a complete species list. J. Crustac. Biol. 2017, 37, 615–653. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, H.H. Biology of Freshwater Crayfish. J. Crustac. Biol. 2002, 22, 969. [Google Scholar] [CrossRef] [Green Version]
- Bouchard, R.W. Taxonomy, ecology and phylogeny of the subgenus Depressicambarus, with the description of a new species from Florida and redescriptions of Cambarus graysoni, Cambarus latimanus and Cambarus striatus (Decapoda: Cambaridae). Bull. Alabama Museum Nat. Hist. 1978, 28, 27–60. [Google Scholar]
- Caine, E.A. Comparative Ecology of Epigean and Hypogean Crayfish (Crustacea: Cambaridae) from Northwestern Florida. Am. Midl. Nat. 1978, 99, 315. [Google Scholar] [CrossRef]
- Taylor, C.A.; Schuster, G.A.; Cooper, J.E.; DiStefano, R.J.; Eversole, A.G.; Hamr, P.; Hobbs, H.H.; Robison, H.W.; Skelton, C.E.; Thoma, R.F. A Reassessment of the Conservation Status of Crayfishes of the United States and Canada after 10+ Years of Increased Awareness. Fisheries 2007, 32, 372–389. [Google Scholar] [CrossRef]
- Chucholl, C.; Schrimpf, A. The decline of endangered stone crayfish (Austropotamobius torrentium) in southern Germany is related to the spread of invasive alien species and land-use change. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 44–56. [Google Scholar] [CrossRef]
- Klobučar, G.I.V.; Podnar, M.; Jelić, M.; Franjević, D.; Faller, M.; Štambuk, A.; Gottstein, S.; Simić, V.; Maguire, I. Role of the Dinaric Karst (western Balkans) in shaping the phylogeographic structure of the threatened crayfish Austropotamobius torrentium. Freshw. Biol. 2013, 58, 1089–1105. [Google Scholar] [CrossRef]
- Pârvulescu, L.; Zaharia, C.; Satmari, A.; Drăguţ, L. Is the distribution pattern of the stone crayfish in the Carpathians related to karstic refugia from Pleistocene glaciations? Freshw. Sci. 2013, 32, 1410–1419. [Google Scholar] [CrossRef] [Green Version]
- Pârvulescu, L.; Pérez-Moreno, J.L.; Panaiotu, C.; Drăguț, L.; Schrimpf, A.; Popovici, I.-D.; Zaharia, C.; Weiperth, A.; Gál, B.; Schubart, C.D.; et al. A journey on plate tectonics sheds light on European crayfish phylogeography. Ecol. Evol. 2019, 9, 1957–1971. [Google Scholar] [CrossRef] [Green Version]
- Pârvulescu, L.; Zaharia, C. Distribution and ecological preferences of noble crayfish in the Carpathian Danube basin: Biogeographical insights into the species history. Hydrobiologia 2014, 726, 53–63. [Google Scholar] [CrossRef]
- Pârvulescu, L.; Zaharia, C. Current limitations of the stone crayfish distribution in Romania: Implications for its conservation status. Limnologica 2013, 43, 143–150. [Google Scholar] [CrossRef]
- Trouilhé, M.-C.; Souty-Grosset, C.; Grandjean, F.; Parinet, B. Physical and chemical water requirements of the white-clawed crayfish (Austropotamobius pallipes) in western France. Aquat. Conserv. Mar. Freshw. Ecosyst. 2007, 17, 520–538. [Google Scholar] [CrossRef]
- Pârvulescu, L.; Zaharia, C.; Groza, M.-I.; Csillik, O.; Satmari, A.; Drăguţ, L. Flash-flood potential: A proxy for crayfish habitat stability. Ecohydrology 2016, 9, 1507–1516. [Google Scholar] [CrossRef]
- Dudley, M.P.; Solomon, K.; Wenger, S.; Jackson, C.R.; Freeman, M.; Elliott, K.J.; Miniat, C.F.; Pringle, C.M. Do crayfish affect stream ecosystem response to riparian vegetation removal? Freshw. Biol. 2021, 66, 1423–1435. [Google Scholar] [CrossRef]
- Pârvulescu, L.; Iorgu, E.-I.; Zaharia, C.; Ion, M.C.; Satmari, A.; Krapal, A.-M.; Popa, O.-P.; Miok, K.; Petrescu, I.; Popa, L.-O. The future of endangered crayfish in light of protected areas and habitat fragmentation. Sci. Rep. 2020, 10, 14870. [Google Scholar] [CrossRef]
- Pacioglu, O.; Theissinger, K.; Alexa, A.; Samoilă, C.; Sîrbu, O.I.; Schrimpf, A.; Zubrod, J.P.; Schulz, R.; Pîrvu, M.; Lele, S.F.; et al. Multifaceted implications of the competition between native and invasive crayfish: A glimmer of hope for the native’s long-term survival. Biol. Invasions 2020, 22, 827–842. [Google Scholar] [CrossRef]
- Pârvulescu, L.; Stoia, D.I.; Miok, K.; Constan Ion, M.; Estera Puha, A.; Sterie, M.; Vereș, M.; Marcu, I.; Muntean, D.; Aburel, O.M. Force and Boldness: Cumulative Assets of a Successful Crayfish Invader. Front. Ecol. Evol. 2021, 9, 1–12. [Google Scholar] [CrossRef]
- Dannewitz, J.; Palm, S.; Edsman, L. Colonization history and human translocations explain the population genetic structure of the noble crayfish (Astacus astacus) in Fennoscandia: Implications for the management of a critically endangered species. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, aqc.3632. [Google Scholar] [CrossRef]
- Mazza, G.; Scalici, M.; Inghilesi, A.; Aquiloni, L.; Pretto, T.; Monaco, A.; Tricarico, E. The Red Alien vs. the Blue Destructor: The Eradication of Cherax destructor by Procambarus clarkii in Latium (Central Italy). Diversity 2018, 10, 126. [Google Scholar] [CrossRef] [Green Version]
- Graham, Z.A.; Angilletta, M.J. Claw size predicts dominance within and between invasive species of crayfish. Anim. Behav. 2020, 166, 153–161. [Google Scholar] [CrossRef]
- Lele, S.-F.; Pârvulescu, L. Experimental evidence of the successful invader Orconectes limosus outcompeting the native Astacus leptodactylus in acquiring shelter and food. Biologia 2017, 72, 877–885. [Google Scholar] [CrossRef]
- Chucholl, C. The bad and the super-bad: Prioritising the threat of six invasive alien to three imperilled native crayfishes. Biol. Invasions 2016, 18, 1967–1988. [Google Scholar] [CrossRef]
- Jussila, J.; Edsman, L.; Maguire, I.; Diéguez-Uribeondo, J.; Theissinger, K. Money Kills Native Ecosystems: European Crayfish as an Example. Front. Ecol. Evol. 2021, 9, 648495. [Google Scholar] [CrossRef]
- Strauss, A.; White, A.; Boots, M. Invading with biological weapons: The importance of disease-mediated invasions. Funct. Ecol. 2012, 26, 1249–1261. [Google Scholar] [CrossRef]
- Manenti, R.; Barzaghi, B.; Nessi, A.; Cioccarelli, S.; Villa, M.; Ficetola, G.F. Not Only Environmental Conditions but Also Human Awareness Matters: A Successful Post-Crayfish Plague Reintroduction of the White-Clawed Crayfish (Austropotamobius pallipes) in Northern Italy. Front. Ecol. Evol. 2021, 9, 621613. [Google Scholar] [CrossRef]
- Wood, T.C.; Smiley, P.C.; Gillespie, R.B.; Gonzalez, J.M.; King, K.W. Injury frequency and severity in crayfish communities as indicators of physical habitat quality and water quality within agricultural headwater streams. Environ. Monit. Assess. 2020, 192, 227. [Google Scholar] [CrossRef]
- Magoulick, D.D.; DiStefano, R.J.; Imhoff, E.M.; Nolen, M.S.; Wagner, B.K. Landscape- and local-scale habitat influences on occupancy and detection probability of stream-dwelling crayfish: Implications for conservation. Hydrobiologia 2017, 799, 217–231. [Google Scholar] [CrossRef]
- Liu, C.; Wolter, C.; Xian, W.; Jeschke, J.M. Species distribution models have limited spatial transferability for invasive species. Ecol. Lett. 2020, 23, 1682–1692. [Google Scholar] [CrossRef]
- Petitpierre, B.; Broennimann, O.; Kueffer, C.; Daehler, C.; Guisan, A. Selecting predictors to maximize the transferability of species distribution models: Lessons from cross-continental plant invasions. Glob. Ecol. Biogeogr. 2017, 26, 275–287. [Google Scholar] [CrossRef]
- Stoeckel, J.A.; Helms, B.S.; Cash, E. Evaluation of a Crayfish Burrowing Chamber Design with Simulated Groundwater Flow. J. Crustac. Biol. 2011, 31, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Ion, M.C.; Puha, A.E.; Suciu, T.; Pârvulescu, L. Get a grip: Unusual disturbances drive crayfish to improvise. Behaviour 2020, 157, 101–120. [Google Scholar] [CrossRef]
- Pârvulescu, L. Introducing a new Austropotamobius crayfish species (Crustacea, Decapoda, Astacidae): A Miocene endemism of the Apuseni Mountains, Romania. Zool. Anz. 2019, 279, 94–102. [Google Scholar] [CrossRef]
- Hengl, T.; De Jesus, J.M.; Heuvelink, G.B.M.; Gonzalez, M.R.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panagos, P.; Borrelli, P.; Meusburger, K. A new European slope length and steepness factor (LS-factor) for modeling soil erosion by water. Geosciences 2015, 5, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Panagos, P.; Borrelli, P.; Meusburger, K.; Alewell, C.; Lugato, E.; Montanarella, L. Estimating the soil erosion cover-management factor at the European scale. Land Use Policy 2015, 48, 38–50. [Google Scholar] [CrossRef]
- Panagos, P.; Borrelli, P.; Poesen, J.; Ballabio, C.; Lugato, E.; Meusburger, K.; Montanarella, L.; Alewell, C. The new assessment of soil loss by water erosion in Europe. Environ. Sci. Policy 2015, 54, 438–447. [Google Scholar] [CrossRef]
- Panagos, P.; Meusburger, K.; Ballabio, C.; Borrelli, P.; Alewell, C. Soil erodibility in Europe: A high-resolution dataset based on LUCAS. Sci. Total Environ. 2014, 479–480, 189–200. [Google Scholar] [CrossRef]
- Genuer, R.; Poggi, J.M.; Tuleau-Malot, C. Variable selection using random forests. Pattern Recognit. Lett. 2010, 31, 2225–2236. [Google Scholar] [CrossRef] [Green Version]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Archer, K.J.; Kimes, R.V. Empirical characterization of random forest variable importance measures. Comput. Stat. Data Anal. 2008, 52, 2249–2260. [Google Scholar] [CrossRef]
- Díaz-Uriarte, R.; Alvarez de Andrés, S. Gene selection and classification of microarray data using random forest. BMC Bioinformatics 2006, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography (Cop.) 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Sepúlveda, M.A.; Bartheld, J.L.; Meynard, C.; Benavides, M.; Astorga, C.; Parra, D.; Medina-Vogel, G. Landscape features and crustacean prey as predictors of the Southern river otter distribution in Chile. Anim. Conserv. 2009, 12, 522–530. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2017. [Google Scholar]
- Liaw, A.; Wiener, M. Classification and Regression by RandomForest. R News 2002, 2, 18–22. [Google Scholar]
- Congalton, R.G. A review of assessing the accuracy of classifications of remotely sensed data. Remote Sens. Environ. 1991, 37, 35–46. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A. Insights into the area under the receiver operating characteristic curve (AUC) as a discrimination measure in species distribution modelling. Glob. Ecol. Biogeogr. 2012, 21, 498–507. [Google Scholar] [CrossRef]
- Piyapong, C.; Tattoni, C.; Ciolli, M.; Dembski, S.; Paradis, E. Modelling the geographical distributions of one native and two introduced species of crayfish in the French Alps. Ecol. Inform. 2020, 60, 101172. [Google Scholar] [CrossRef]
- Holdich, D.M. Distribution of crayfish in Europe and some adjoining countries. Bull. Français Pêche Piscic. 2002, 367, 611–650. [Google Scholar] [CrossRef] [Green Version]
- Vlach, P.; Fischer, D.; Hulec, L. Microhabitat preferences of the stone crayfish Austropotamobius torrentium (Schrank, 1803). Knowl. Manag. Aquat. Ecosyst. 2009, 394–395, 15. [Google Scholar] [CrossRef] [Green Version]
- Grow, L.; Merchant, H. The burrow habitat of the crayfish, Cambarus diogenes diogenes (Girard) (Girard). Am. Midl. Nat. 1980, 103, 231. [Google Scholar] [CrossRef]
- Wiśniewski, K.; Szarmach, D.; Poznańska-Kakareko, M. The role of abiotic and biotic factors in interspecific competition of Polish crayfish-Comprehensive literature review. Oceanol. Hydrobiol. Stud. 2020, 49, 428–441. [Google Scholar] [CrossRef]
- Valido, C.A.R.; Johnson, M.F.; Dugdale, S.J.; Cutts, V.; Fell, H.G.; Higgins, E.A.; Tarr, S.; Templey, C.M.; Algar, A.C. Thermal sensitivity of feeding and burrowing activity of an invasive crayfish in UK waters. Ecohydrology 2021, 14, e2258. [Google Scholar] [CrossRef]
- Garvey, J.E.; Stein, R.A.; Thomas, H.M. Assessing How Fish Predation and Interspecific Prey Competition Influence a Crayfish Assemblage. Ecology 1994, 75, 532–547. [Google Scholar] [CrossRef]
- England, L.E.; Rosemond, A.D. Small reductions in forest cover weaken terrestrial-aquatic linkages in headwater streams. Freshw. Biol. 2004, 49, 721–734. [Google Scholar] [CrossRef]
- Quinn, J.M.; Williamson, R.B.; Smith, R.K.; Vickers, M.L. Effects of riparian grazing and channelisation on streams in Southland, New Zealand. 2. Benthic invertebrates. N. Z. J. Mar. Freshw. Res. 2010, 26, 259–273. [Google Scholar] [CrossRef] [Green Version]
- Pârvulescu, L.; Hamchevici, C. The relation between water quality and the distribution of Gammarus balcanicus schäferna 1922 (Amphipoda: Gammaridae) in the Anina mountains. Carpathian J. Earth Environ. Sci. 2010, 5, 161–168. [Google Scholar]
- Ungureanu, E.; Mojžišová, M.; Tangerman, M.; Ion, M.C.; Parvulescu, L.; Petrusek, A. The spatial distribution of Aphanomyces astaci genotypes across Europe: Introducing the first data from Ukraine. Freshw. Crayfish 2020, 25, 77–87. [Google Scholar] [CrossRef]
- Di Domenico, M.; Curini, V.; Caprioli, R.; Giansante, C.; Mrugała, A.; Mojžišová, M.; Cammà, C.; Petrusek, A. Real-Time PCR Assays for Rapid Identification of Common Aphanomyces astaci Genotypes. Front. Ecol. Evol. 2021, 9, 597585. [Google Scholar] [CrossRef]
- Barnett, Z.C.; Adams, S.B. Review of Dam Effects on Native and Invasive Crayfishes Illustrates Complex Choices for Conservation Planning. Front. Ecol. Evol. 2021, 8, 621723. [Google Scholar] [CrossRef]
- Krieg, R.; King, A.; Zenker, A. Measures to Control Invasive Crayfish Species in Switzerland: A Success Story? Front. Environ. Sci. 2020, 8, 609129. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crayfish Status | Absolute Depth to Bedrock (cm) | Bulk Density (kg/m3) | Clay Content (%) | Silt Content (%) | Sand Content (%) | Coarse Fragments (%) | Soil Erosion by Water (t/ha.yr) | Soil Erodibility | |
|---|---|---|---|---|---|---|---|---|---|
| A. bihariensis | Absence | 19.03 | 1400.5 | - | - | - | 9.0 | - | 0.0 |
| Presence | 15.6 | 1376.3 | - | - | - | 12.3 | - | 0.0 | |
| A. torrentium | Absence | - | 1383.3 | 25.8 | 38.6 | 35.7 | 9.9 | 6.6 | - |
| Presence | - | 1383.1 | 25.6 | 37.7 | 36.7 | 11.4 | 5.4 | - | |
| A. astacus | Absence | 20.6 | 1375.2 | 25.2 | 39.8 | 35.0 | 11.2 | 7.3 | 0.028 |
| Presence | 20.2 | 1397.0 | 26.4 | 39.7 | 34.0 | 9.4 | 8.0 | 0.029 | |
| P. leptodactylus | Absence | 19.6 | 1381.1 | 25.4 | 39.2 | 35.4 | 11.1 | 7.2 | 0.03 |
| Presence | 27.4 | 1416.7 | 29.2 | 40.6 | 30.1 | 3.7 | 4.1 | 0.03 |
| Absolute Depth to Bedrock (cm) | Bulk Density (kg/m3) | Clay Content (%) | Silt Content (%) | Sand Content (%) | Coarse Fragments (%) | Soil Erosion by Water (t/ha.yr) | Soil Erodibility | |
|---|---|---|---|---|---|---|---|---|
| A. bihariensis | −0.60 ** | −0.17 | −0.02 | 0.12 | −0.11 | 0.27 | −0.10 | −0.17 |
| A. torrentium | −0.07 | −0.08 | 0.02 | −0.18 * | 0.06 | 0.08 | −0.12 | 0.06 |
| A. astacus | 0.008 | 0.16 ** | 0.18 ** | 0.02 | −0.13 * | −0.14 ** | 0.04 | 0.18 ** |
| P. leptodactylus | 0.44 ** | 0.22 ** | 0.31 ** | 0.15 ** | −0.30 ** | −0.46 ** | −0.09 | 0.08 |
| OA | AUC | |
|---|---|---|
| A. bihariensis | 0.69 | 0.94 |
| A. torrentium | 0.67 | 0.72 |
| A. astacus | 0.66 | 0.74 |
| P. leptodactylus | 0.94 | 0.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dornik, A.; Ion, M.C.; Chețan, M.A.; Pârvulescu, L. Soil-Related Predictors for Distribution Modelling of Four European Crayfish Species. Water 2021, 13, 2280. https://doi.org/10.3390/w13162280
Dornik A, Ion MC, Chețan MA, Pârvulescu L. Soil-Related Predictors for Distribution Modelling of Four European Crayfish Species. Water. 2021; 13(16):2280. https://doi.org/10.3390/w13162280
Chicago/Turabian StyleDornik, Andrei, Mihaela Constanța Ion, Marinela Adriana Chețan, and Lucian Pârvulescu. 2021. "Soil-Related Predictors for Distribution Modelling of Four European Crayfish Species" Water 13, no. 16: 2280. https://doi.org/10.3390/w13162280
APA StyleDornik, A., Ion, M. C., Chețan, M. A., & Pârvulescu, L. (2021). Soil-Related Predictors for Distribution Modelling of Four European Crayfish Species. Water, 13(16), 2280. https://doi.org/10.3390/w13162280