
Size Matters, but Species Do Not: No Evidence for Species-Specific Swimming Performance in Co-Occurring Great Basin Stream Fishes


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
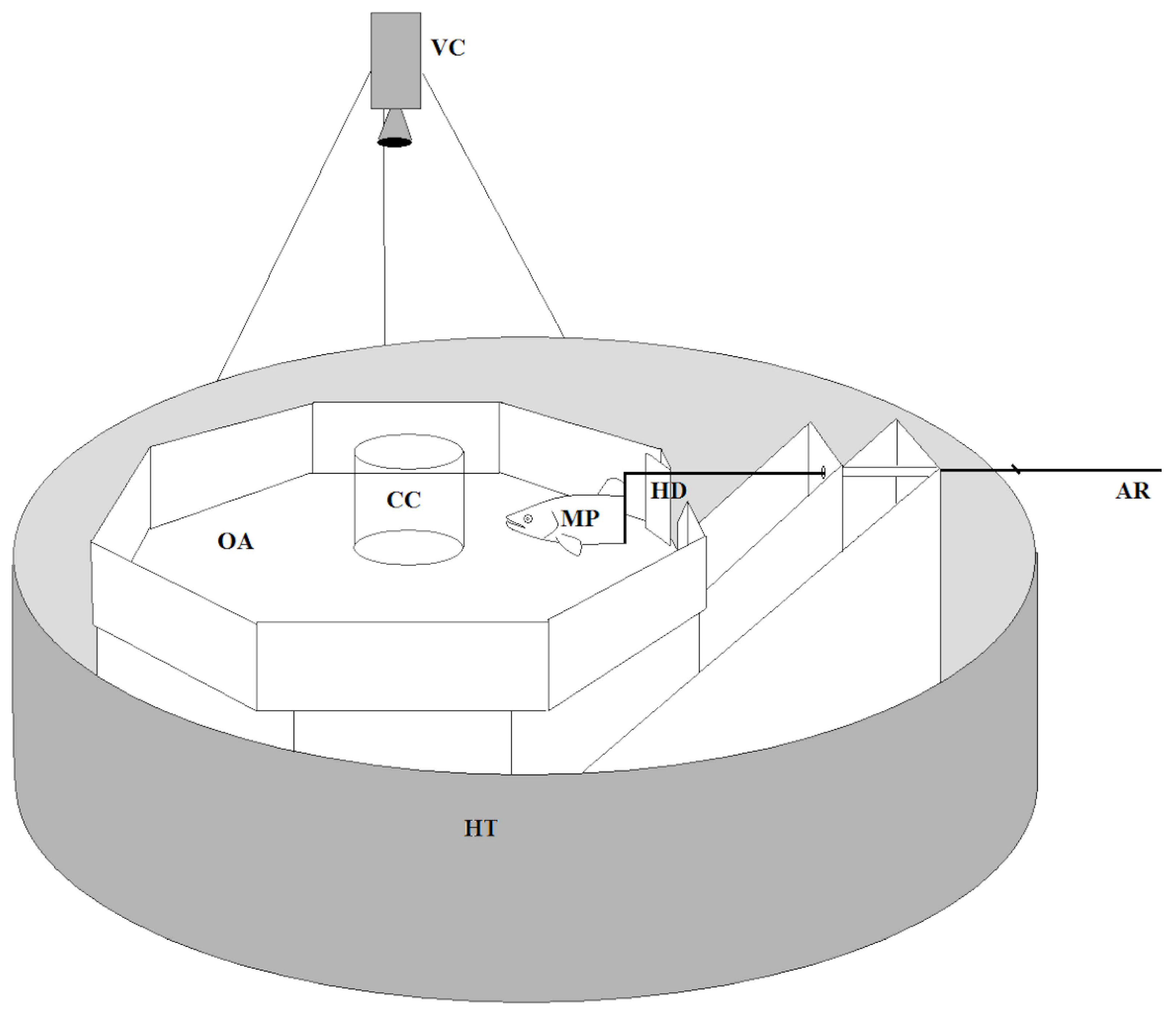
2.1. Collection & Maintenance
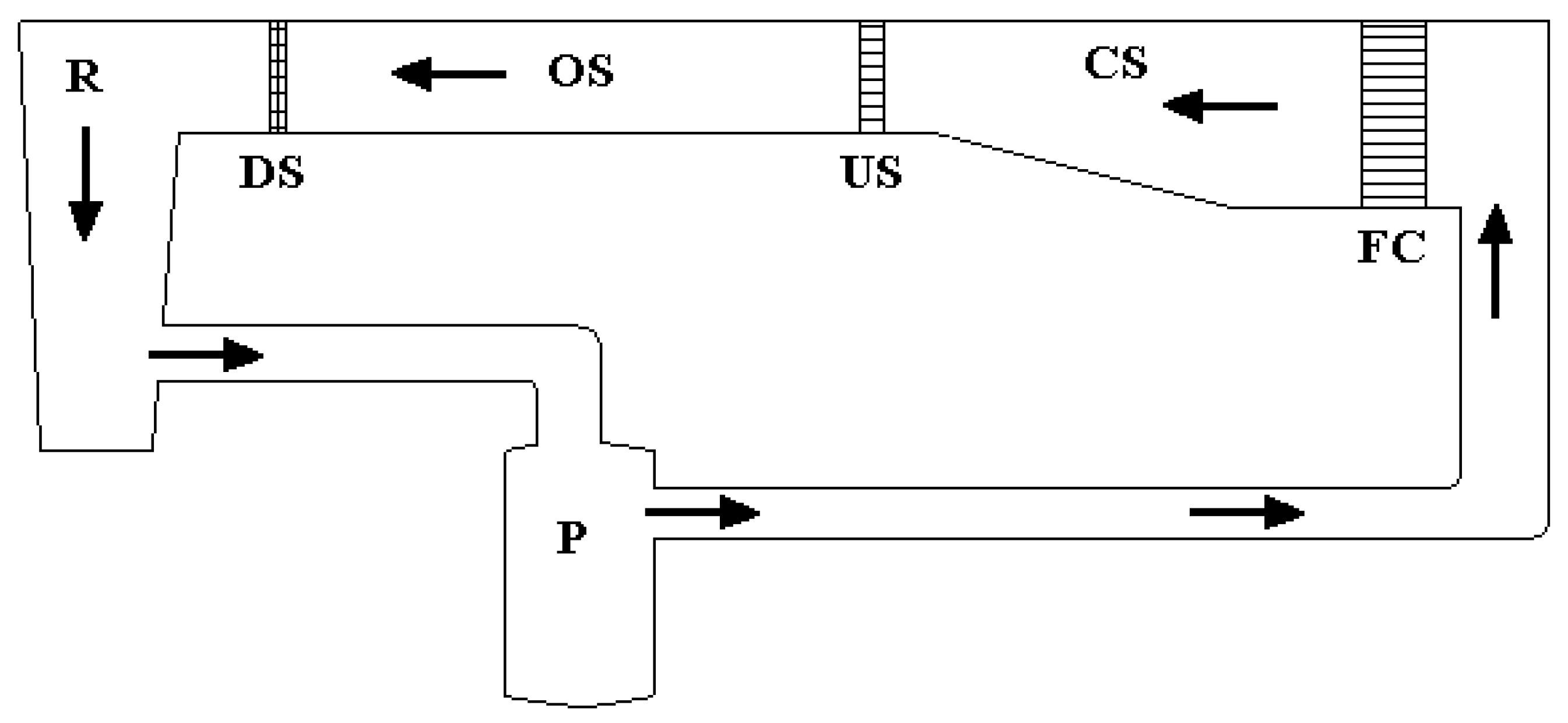
2.2. Burst Speed
2.3. Prolonged Speed
2.4. Statistical Analysis
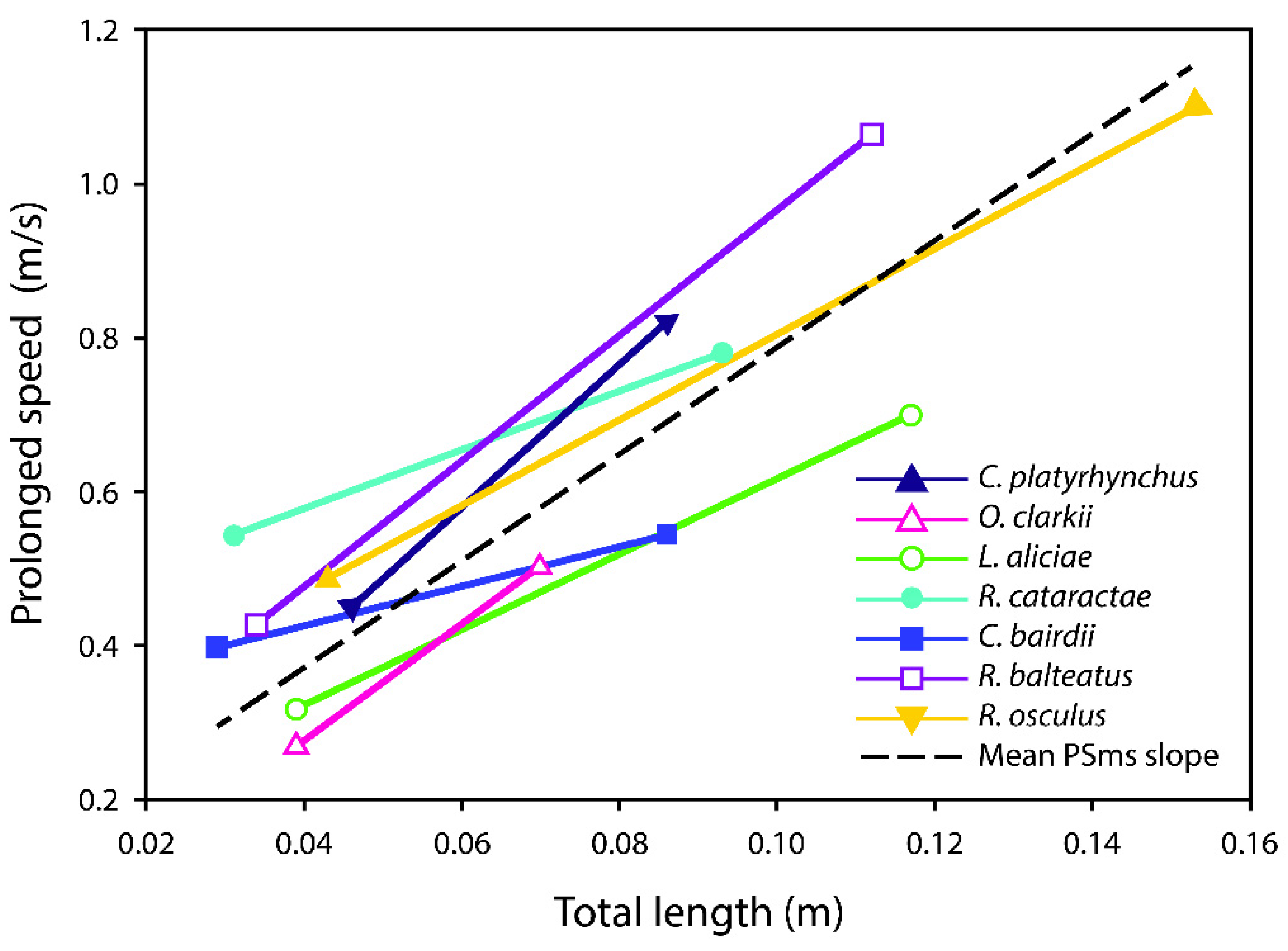
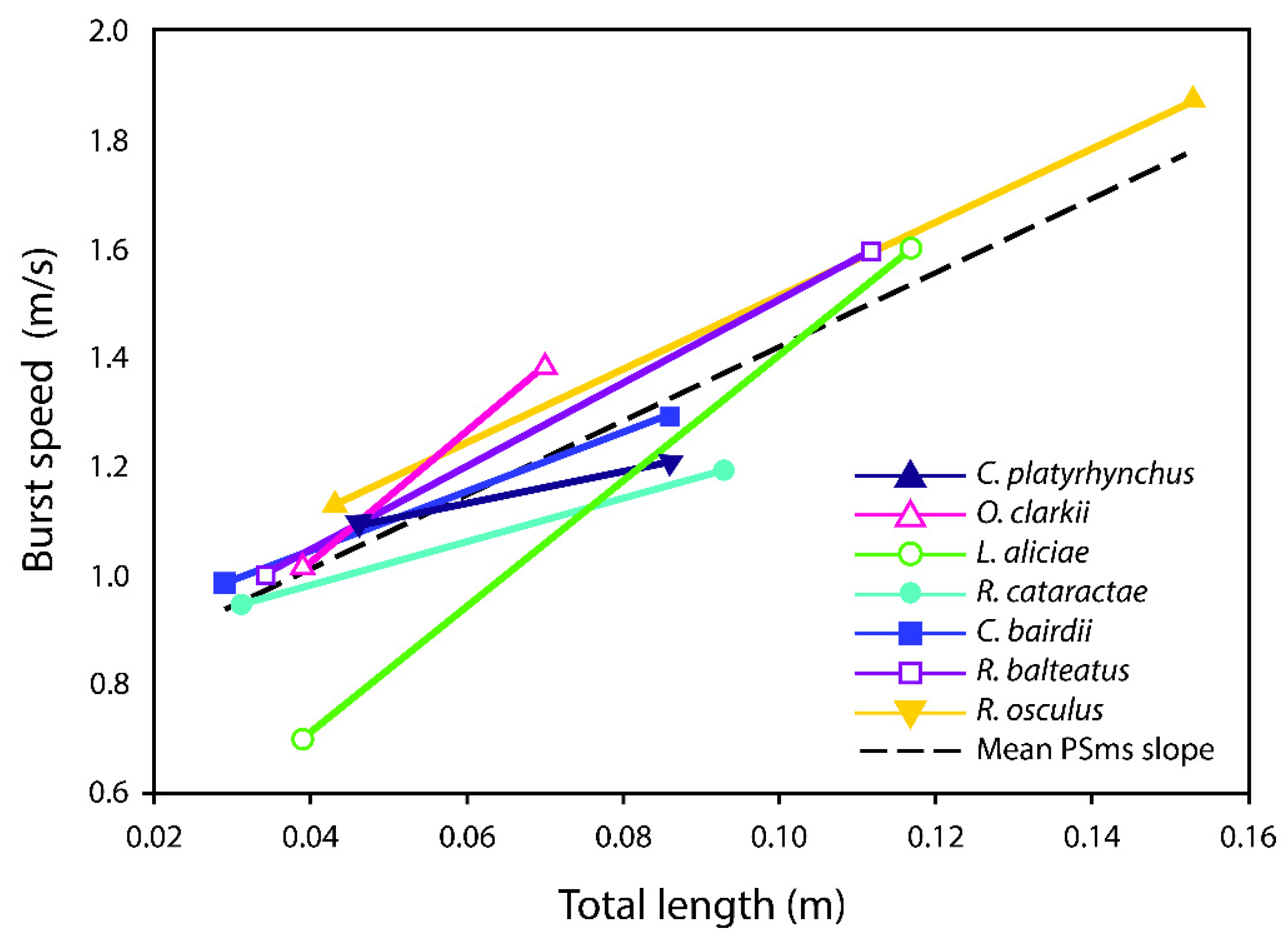
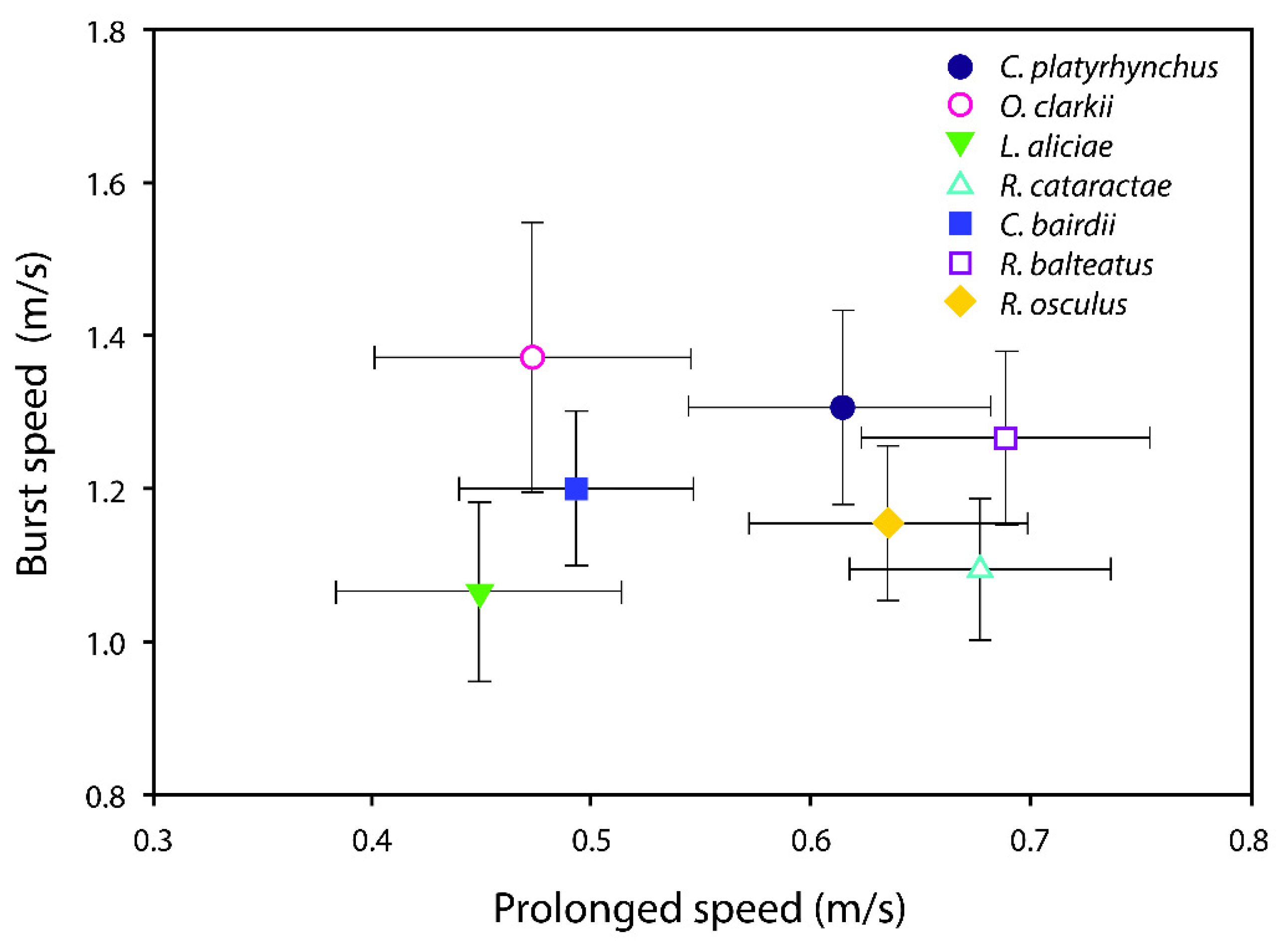
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meyers, P.J.; Belk, M.C. Shape variation in a benthic stream fish across flow regimes. Hydrobiologia 2014, 738, 147–154. [Google Scholar] [CrossRef]
- Barros, T.F.; Louvise, J.; Caramashi, E.P. Flow gradient drives morphological divergence in an Amazon pelagic stream fish. Hydrobiologia 2019, 833, 217–229. [Google Scholar] [CrossRef]
- Beamish, F.H. Swimming capacity. In Fish Physiology; Hoar, W.S., Randall, D.J., Eds.; Academic Press: New York, NY, USA; London, UK, 1978; Volume 7, pp. 101–187. [Google Scholar]
- Videler, J.J.; Wardle, C.S. Fish swimming stride by stride: Speed limits and endurance. Rev. Fish Biol. Fish. 1991, 1, 23–40. [Google Scholar] [CrossRef]
- Hammer, C. Fatigue and exercise tests with fish. Comp. Biochem. Physiol. Part A Phys. 1995, 112, 1–20. [Google Scholar] [CrossRef]
- Pakkasmaa, S.; Piironen, J. Water velocity shapes juvenile salmonids. Ecol. Evol. 2000, 14, 721–730. [Google Scholar] [CrossRef]
- Spoljaric, M.A.; Reimchen, T.E. Habitat-specific trends in ontogeny of body shape in stickleback from coastal archipelago: Potential for rapid shifts in colonizing populations. J. Morphol. 2011, 272, 590–597. [Google Scholar] [CrossRef]
- Rincón, P.; Bastir, M.; Grossman, G. Form and performance: Body shape and prey-capture success in four drift-feeding minnows. Oecologia 2007, 152, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.B.; Mcphail, J.D. Burst swimming and size-related predation of newly emerged coho salmon Oncorhynchus kisutch. Trans. Am. Fish. Soc. 1985, 114, 546–551. [Google Scholar] [CrossRef]
- Walker, J.A.; Ghalambor, C.K.; Griset, O.L.; McKenney, D.; Reznick, D.N. Do faster starts increase the probability of evading predators? Funct. Ecol. 2005, 19, 751–897. [Google Scholar] [CrossRef]
- Videler, J.J. Fish Swimming; Chapman & Hall: London, UK, 1993. [Google Scholar]
- Kolok, A.S. Interindividual variation in the prolonged locomotor performance of ectothermic vertebrates: A comparison of fish and herpetofaunal methodologies and a brief review of the recent fish literature. Can. J. Fish. Aquat. Sci. 1999, 56, 700–710. [Google Scholar] [CrossRef]
- Reidy, S.P.; Kerr, S.R.; Nelson, J.A. Aerobic and anaerobic swimming performance of individual Atlantic cod. J. Exp. Biol. 2000, 203, 347–357. [Google Scholar] [CrossRef]
- Plaut, I. Critical swimming speed: Its ecological relevance. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 131, 41–50. [Google Scholar] [CrossRef]
- Rubio-Gracio, F.; García-Berthou, E.; Guasch, H.; Zamora, L.; Vila-Gispert, A. Size-related effects and the influence of metabolic traits and morphology on swimming performance in fish. Curr. Zool. 2020, 66, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Webb, P.W.; Skadsen, J.M. Strike tactics of Esox. Can. J. Zool. Rev. Can. Zool. 1980, 58, 1462–1469. [Google Scholar] [CrossRef]
- Webb, P.W. Body form, locomotion and foraging in aquatic vertebrates. Am. Zool. 1984, 24, 107–120. [Google Scholar] [CrossRef]
- Ojanguren, A.F.; Brana, F. Effects of size and morphology on swimming performance in juvenile brown trout (Salmo trutta L.). Ecol. Freshw. Fish 2003, 12, 241–246. [Google Scholar] [CrossRef]
- Wakeling, J.M. Fast start mechanics. In Fish Biomechanics: Fish Physiology; Shadwick, R.E., Lauder, G.V., Eds.; Academic Press: London, UK, 2007; pp. 333–368. [Google Scholar]
- Hynes, H.B. The Ecology of Running Water; University of Toronto Press: Toronto, ON, Canada, 1970. [Google Scholar]
- Webb, P.W. Fast-start resistance of trout. J. Exp. Biol. 1982, 96, 93–106. [Google Scholar] [CrossRef]
- Webb, P.W. Locomotor patterns in the evolution of actinopterygian fishes. Am. Zool. 1982, 22, 329–342. [Google Scholar] [CrossRef]
- Weihs, D. A hydrodynamical analysis of fish turning manoeuvres. Proc. R. Soc. Lond. 1972, 182, 59–72. [Google Scholar]
- Weihs, D. The mechanism of rapid starting of a slender fish. Biorheology 1973, 10, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Webb, P.W. Effects of partial caudal-fin amputation on the kinematics and metabolic rate of underyearling sockeye salmon (Oncorhynchus nerka) at steady swimming speeds. J. Exp. Biol. 1973, 59, 565–582. [Google Scholar] [CrossRef]
- Lighthill, M.J. Mathematical Biofluid Dynamics; CBMS-NSF Regional Conference Series in Applied Mathematics; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 1975. [Google Scholar]
- Webb, P.W. Effects of median-fin amputation on fast-start performance of rainbow trout (Salmo gairdneri). J. Exp. Biol. 1977, 68, 123–135. [Google Scholar] [CrossRef]
- Willis, S.C.; Winemiller, K.O.; Lopez-Fernandez, H. Habitat structural complexity and morphological diversity of fish assemblages in a Neotropical floodplain river. Oecologia 2005, 142, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Brinsmead, J.; Fox, M.G. Morphological variation between lake- and stream-dwelling rock bass and pumpkinseed populations. J. Fish Biol. 2002, 61, 1619–1638. [Google Scholar] [CrossRef]
- Naspleda, J.; Vila-Gispert, A.; Fox, M.G.; Zamora, L.; Ruiz-Navarro, A. Morphological variation between non-native lake- and stream-dwelling pumpkinseed Lepomis gibbosus in the Iberian Peninsula. J. Fish Biol. 2012, 81, 1915–1935. [Google Scholar] [CrossRef]
- Prenosil, E.; Koupal, K.; Grauf, J.; Schoenebeck, C.; Hoback, W.W. Swimming and jumping ability of 10 Great Plains fish species. J. Freshw. Ecol. 2016, 31, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Ohlberger, J.; Staaks, G.; Hölker, F. Swimming efficiency and the influence of morphology on swimming costs in fishes. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2006, 176, 17–25. [Google Scholar] [CrossRef]
- Mchenry, M.J.; Lauder, G.V. Ontogeny of form and function: Locomotor morphology and drag in zebrafish (Danio rerio). J. Morphol. 2006, 267, 1099–1109. [Google Scholar] [CrossRef]
- Roscoe, D.W.; Hinch, S.G. Effectiveness monitoring of fish passage facilities: Historical trends, geographic patterns and future directions. Fish Fish. 2010, 11, 12–33. [Google Scholar] [CrossRef]
- Lee, C.G.; Farrell, A.P.; Lotto, A.; MacNutt, M.J.; Hinch, S.G.; Healey, M.C. The effect of temperature on swimming performance and oxygen consumption in adult sockeye (Oncorhynchus nerka) and coho (O. kisutch) salmon stocks. J. Exp. Biol. 2003, 206, 3239–3251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, J.M.; Elliott, J.A. Temperature requirements of Atlantic salmon Salmo salar, brown trout Salmo trutta and Arctic charr Salvenlinus aplinus: Predicting the effects of climate change. J. Fish Biol. 2010, 77, 1793–1817. [Google Scholar] [CrossRef]
- Brett, J.R. The respiratory metabolism and swimming performance of young sockeye salmon. J. Fish. Board Can. 1964, 21, 1183–1226. [Google Scholar] [CrossRef]
- Ward, D.L.; Schultz, A.A.; Matson, P.G. Differences in swimming ability and behaviour in response to high water velocities among native and nonnative fishes. Environ. Biol. Fishes 2003, 68, 87–92. [Google Scholar] [CrossRef]
- Underwood, Z.E.; Myrick, C.A.; Compton, R.I. Comparative swimming performance of five catostomus species and roundtail chub. N. Am. J. Fish. Manag. 2014, 34, 753–763. [Google Scholar] [CrossRef]
- Dockery, D.R.; McMahon, T.E.; Kappenman, K.M.; Blank, M. Evaluation of swimming performance for fish passage of longnose dace Rhinichthys cataractae using an experimental flume. J. Fish Biol. 2017, 90, 980–1000. [Google Scholar] [CrossRef]
- Sigler, W.F.; Sigler, J.W. Fishes of the Great Basin: A Natural History; University of Nevada Press: Reno, NV, USA, 1987. [Google Scholar]
- Peake, S.; Mckinley, R.S.; Scruton, D.A. Swimming performance of various freshwater Newfoundland salmonids relative to habitat selection and fishway design. J. Fish Biol. 1997, 51, 710–723. [Google Scholar] [CrossRef]
- Wolter, C.; Arlinghaus, R. Navigation impacts on freshwater fish assemblages: The ecological relevance of swimming performance. Rev. Fish Biol. Fish. 2003, 13, 63–89. [Google Scholar] [CrossRef]
- Kimball, M.E.; Boswell, K.M.; Rozas, L.P.; Berwaldt, E.K.; Richards, A.R. Swimming abilities of juvenile estuarine fishes: Implications for passage at water control structures. Wetl. Ecol. Manag. 2018, 26, 383–390. [Google Scholar] [CrossRef]
- Ficke, A.D.; Myrick, C.A.; Jud, N. The Swimming and Jumping Ability of Three Small Great Plains Fishes: Implications for Fishway Design. Trans. Am. Fish. 2011, 140, 1521–1531. [Google Scholar] [CrossRef]
- Kane, E.A.; Roeder, M.M.; Derue, M.L.; Ghalambor, C.K. Integration between swim speed and mouth size evolves repeatedly in Trinidadian guppies and aligns with suction-feeding fishes. J. Exp. Biol. 2019, 222, jeb190165. [Google Scholar] [CrossRef] [Green Version]
- Kern, P.; Cramp, R.L.; Gordos, M.A.; Watson, J.R.; Franklin, C.E. Measuring Ucrit and endurance: Equipment choice influences estimates of fish swimming performance. J. Fish Biol. 2018, 92, 237–247. [Google Scholar] [CrossRef]
- Silva, A.T.; Lucas, M.C.; Castro-Santo, T.; Katopodis, C.; Baumgartner, L.J.; Thiem, J.D.; Aarestrup, K.; Pompeu, P.S.; O’Brien, G.C.; Braun, D.C.; et al. The future of fish passage science, engineering, and practice. Fish Fish. 2018, 19, 340–362. [Google Scholar] [CrossRef] [Green Version]
- Langerhans, R.B. Predictability of phenotypic differentiation across flow regimes in fishes. Integr. Comp. Biol. 2008, 48, 699–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimmer, D.M.; Saunders, R.L.; Paim, U. Effects of temperature and season on the position holding performance of juvenile Atlantic salmon (Salmo salar). Can. J. Zool. Rev. Can. Zool. 1985, 63, 92–96. [Google Scholar] [CrossRef]
- Arnold, G.P.; Webb, P.W.; Holford, B.H. The role of the pectoral fins in station-holding of atlantic salmon parr. (Salmo salar). J. Exp. Biol. 1991, 156, 625–629. [Google Scholar] [CrossRef]
- Billman, E.J.; Pyron, M. Evolution of form and function: Morphology and swimming performance in North American minnows. J. Freshw. Ecol. 2005, 20, 221–232. [Google Scholar] [CrossRef]
- Warren, M.L.; Pardew, M.G. Road crossings as barriers to small-stream fish movement. Trans. Am. Fish. Soc. 1998, 127, 637–644. [Google Scholar] [CrossRef]
- Gibson, R.J.; Haedrich, R.L.; Wernerheim, C.M. Loss of fish habitat as a consequence of inappropriately constructed stream crossings. Fisheries 2005, 30, 10–17. [Google Scholar] [CrossRef]
- Johnson, K.; Wait, L.; Monk, S.; Rader, R.; Hotchkiss, R.; Belk, M. Effects of Substrate on Movement Patterns and Behavior of Stream Fish through Culverts: An Experimental Approach. Sustainability 2019, 11, 470. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, E.M.; Heaslip, B.M.; Cramp, R.L.; Riches, M.; Gordos, M.A.; Franklin, C.E. Substrate roughening improves swimming performance in two small-bodied riverine fishes: Implications for culvert remediation and design. Conserv. Physiol. 2017, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Goodrich, H.R.; Watson, J.R.; Cramp, R.L.; Gordos, M.A.; Franklin, C.E. Making culverts great again. Efficacy of a common culvert remediation strategy across sympatric fish species. Ecol. Eng. 2018, 116, 143–153. [Google Scholar] [CrossRef]
- Bunt, C.M.; Castro-Santos, T.; Haro, A. Performance of fish passage structures at upstream barriers to migration. River Res. Appl. 2011, 28, 457–478. [Google Scholar] [CrossRef]
- Yan, G.J.; He, X.K.; Cao, Z.D.; Fu, S.J. An interspecific comparison between morphology and swimming performance in cyprinids. J. Evol. Biol. 2013, 26, 1802–1815. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | N | Collection Location | Standard Length (mm) | Range (mm) | SE |
|---|---|---|---|---|---|
| Oncorhynchus clarkii utah | 53 | Little Dell & Manning Meadows Hatcheries, UT, USA | 54.67 | 39–70 | 1.26 |
| Lepidomeda aliciae | 45 | Soldier Creek, UT, USA | 83.00 | 39–107 | 2.57 |
| Rhinichthys cataractae | 56 | Soldier Creek, UT, USA | 65.18 | 35–91 | 2.33 |
| Cottus bairdii | 52 | Diamond Fork, UT, USA | 63.30 | 29–86 | 1.56 |
| Catostomus platyrhynchus | 44 | Soldier Creek, UT, USA | 93.91 | 43–150 | 4.82 |
| Richardsonius balteatus | 30 | Fish Creek, UT, USA | 79.80 | 34–112 | 3.14 |
| Rhinichthys osculus | 46 | Salina Creek, UT, USA | 69.96 | 46–86 | 1.65 |
| Species | NProlonged | Mean Prolonged Speed (m/s) | Confidence Interval | Prolonged Speed Slope | Confidence Interval |
|---|---|---|---|---|---|
| Catostomus platyrhynchus | 15 | 0.61 | 0.54–0.69 | 5.57 | 3.11–8.04 |
| Oncorhynchus clarkii utah | 42 | 0.47 | 0.40–0.55 | 7.55 | 5.59–9.50 |
| Lepidomeda aliciae | 22 | 0.45 | 0.38–0.51 | 4.92 | 1.55–8.28 |
| Rhinichthys cataractae | 15 | 0.68 | 0.62–0.74 | 3.84 | −2.34–10.01 |
| Cottus bairdii | 25 | 0.49 | 0.44–0.55 | 2.59 | −2.65–7.83 |
| Richardsonius balteatus | 19 | 0.69 | 0.62–0.75 | 8.19 | 5.33–11.06 |
| Rhinichthys osculus | 20 | 0.63 | 0.57–0.70 | 9.32 | 4.15–14.49 |
| Source of Variation | df | F Value | p Value |
|---|---|---|---|
| Species | 6 | 1.31 | 0.2536 |
| Total length | 1 | 61.84 | <0.0001 |
| Sp × TL | 6 | 1.44 | 0.2011 |
| Species | NBurst | Mean Burst Speed (m/s) | Confidence Interval (95%) | Burst Speed Slope | Confidence Interval (95%) |
|---|---|---|---|---|---|
| Catostomus platyrhynchus | 25 | 1.31 | 1.18–1.44 | 6.74 | 2.55–10.94 |
| Oncorhynchus clarki utah | 47 | 1.37 | 1.19–1.55 | 11.75 | 3.87–19.58 |
| Lepidomeda aliciae | 38 | 1.07 | 0.95–1.18 | 12.27 | 5.95–18.59 |
| Rhinichthys cataractae | 27 | 1.10 | 1.01–1.19 | 3.96 | −2.22–10.14 |
| Cottus bairdii | 46 | 1.20 | 1.10–1.30 | 5.38 | 0.43–10.34 |
| Richardsonius balteatus | 29 | 1.27 | 1.16–1.38 | 7.61 | 0.86–14.37 |
| Rhinichthys osculus | 21 | 1.16 | 1.06–1.26 | 2.91 | −9.35–15.17 |
| Source of Variation | df | F Value | p Value |
|---|---|---|---|
| Species | 6 | 0.95 | 0.4627 |
| Total length | 1 | 27.91 | <0.001 |
| Sp × TL | 6 | 0.95 | 0.4601 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aedo, J.R.; Otto, K.R.; Rader, R.B.; Hotchkiss, R.H.; Belk, M.C. Size Matters, but Species Do Not: No Evidence for Species-Specific Swimming Performance in Co-Occurring Great Basin Stream Fishes. Water 2021, 13, 2570. https://doi.org/10.3390/w13182570
Aedo JR, Otto KR, Rader RB, Hotchkiss RH, Belk MC. Size Matters, but Species Do Not: No Evidence for Species-Specific Swimming Performance in Co-Occurring Great Basin Stream Fishes. Water. 2021; 13(18):2570. https://doi.org/10.3390/w13182570
Chicago/Turabian StyleAedo, John R., Keith R. Otto, Russell B. Rader, Rollin H. Hotchkiss, and Mark C. Belk. 2021. "Size Matters, but Species Do Not: No Evidence for Species-Specific Swimming Performance in Co-Occurring Great Basin Stream Fishes" Water 13, no. 18: 2570. https://doi.org/10.3390/w13182570
APA StyleAedo, J. R., Otto, K. R., Rader, R. B., Hotchkiss, R. H., & Belk, M. C. (2021). Size Matters, but Species Do Not: No Evidence for Species-Specific Swimming Performance in Co-Occurring Great Basin Stream Fishes. Water, 13(18), 2570. https://doi.org/10.3390/w13182570