
Conservation Genetics of Mediterranean Brown Trout in Central Italy (Latium): A Multi-Marker Approach

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
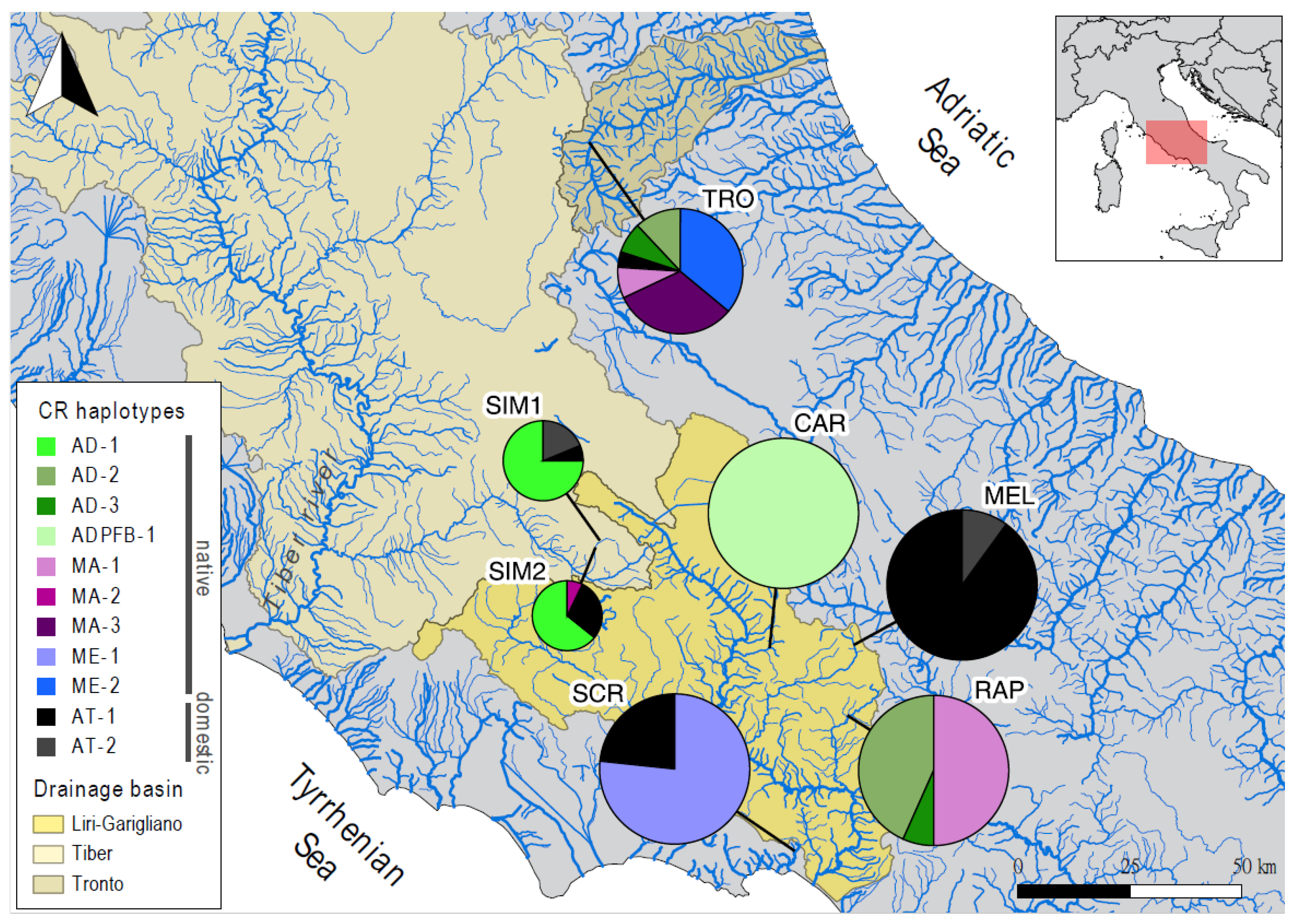
2.1. Study Area, Sample Collection, and DNA Extraction
2.2. Amplification and Genotyping of Mitochondrial and Nuclear Loci
2.3. Population Genetic Diversity and Relationship with Introgression from Domestic Trout
2.4. Population Structure and Demography
3. Results
3.1. Mitochondrial and Nuclear Polymorphism and Population Diversity
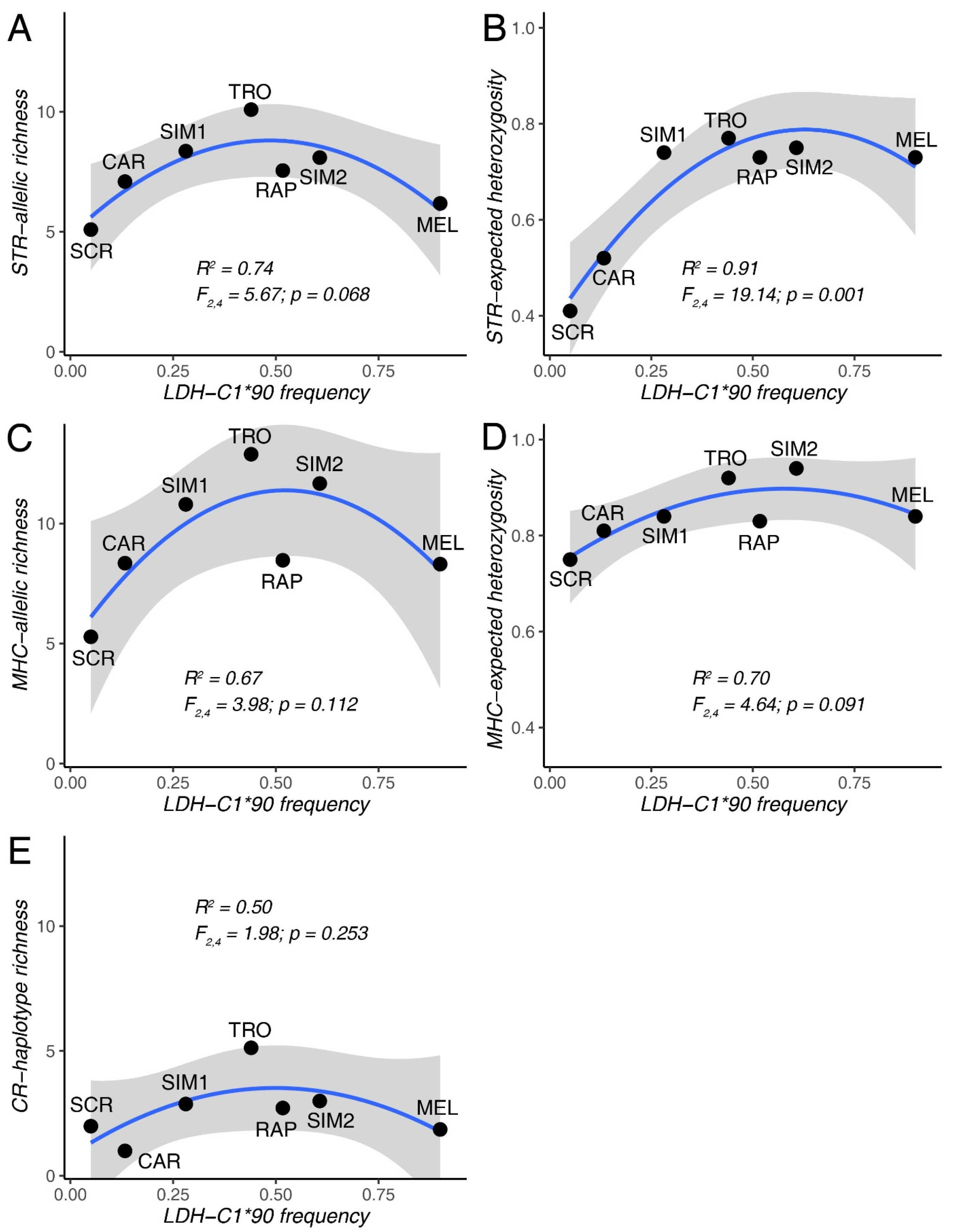
3.2. Extent of Introgression and Its Relationship with Population Diversity
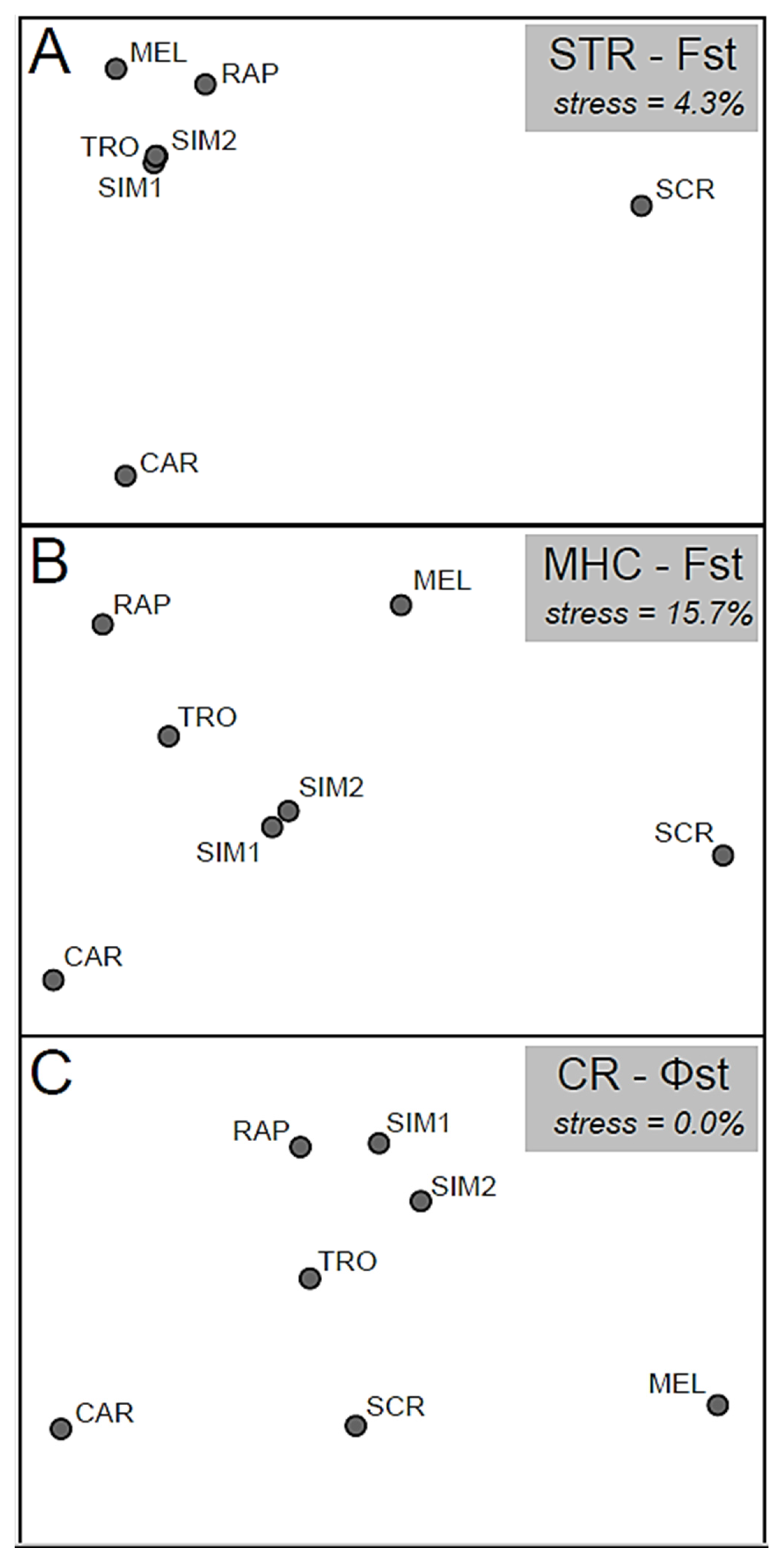
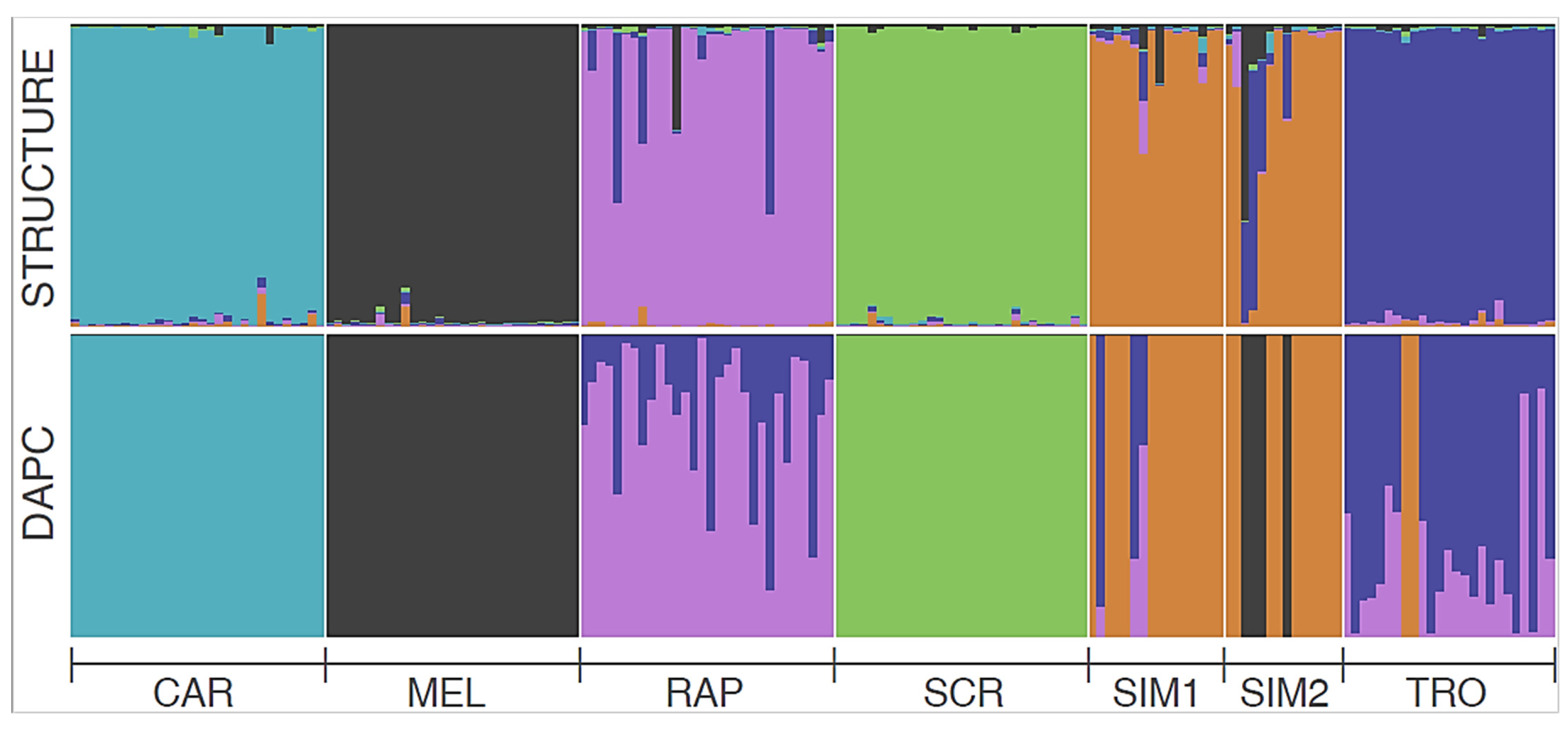
3.3. Genetic Population Structure and Demography
4. Discussion
4.1. Genetic Diversity
4.2. Population Structure and Demography
4.3. Management Implications
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meraner, A.; Gandolfi, A. Genetics of the genus Salmo in Italy: Evolutionary history, population structure, molecular ecology and conservation. In Brown Trout: Biology, Ecology and Management; Lobón-Cerviá, J., Sanz, N., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2018; pp. 65–102. [Google Scholar]
- Sanz, N. Phylogeographic history of brown trout: A review. In Brown Trout: Biology, Ecology and Management; Lobón-Cerviá, J., Sanz, N., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2018; pp. 17–64. [Google Scholar]
- Segherloo, I.H.; Freyhof, J.; Berrebi, P.; Ferchaud, A.-L.; Geiger, M.; Laroche, J.; Levin, B.A.; Normandeau, E.; Bernatchez, L. A genomic perspective on an old question: Salmo trouts or Salmo trutta (Teleostei: Salmonidae)? Mol. Phylogenet. Evol. 2021, 162, 107204. [Google Scholar] [CrossRef] [PubMed]
- Tougard, C.; Justy, F.; Guinand, B.; Douzery, E.J.P.; Berrebi, P. Salmo macrostigma (Teleostei, Salmonidae): Nothing more than a brown trout (S. trutta) lineage? J. Fish Biol. 2018, 93, 302–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernatchez, L. The evolutionary history of brown trout (Salmo trutta L.) inferred from phylogeographic, nested clade, and mismatch analyses of mitochondrial DNA variation. Evolution 2001, 55, 351–379. [Google Scholar] [CrossRef] [PubMed]
- Splendiani, A.; Giovannotti, M.; Righi, T.; Fioravanti, T.; Cerioni, P.N.; Lorenzoni, M.; Carosi, A.; La Porta, G.; Barucchi, V.C. Introgression despite protection: The case of native brown trout in Natura 2000 network in Italy. Conserv. Genet. 2019, 20, 343–356. [Google Scholar] [CrossRef]
- Madeira, M.J.; Gómez-Moliner, B.J.; Machordom Barbé, A. Genetic introgression on freshwater fish populations caused by restocking programmes. Biol. Invasions 2005, 7, 117–125. [Google Scholar] [CrossRef]
- Ferguson, A. Genetic Impacts of Stocking on Indigenous Brown Trout Populations (Environment Agency Science Report No. SC040071/SR). 2007; ISBN 978-1-84432-798-0. Available online: https://www.gov.uk/government/publications/genetic-impacts-of-stocking-on-indigenous-brown-trout-populations (accessed on 3 July 2021).
- Global Invasive Species Database. Available online: http://193.206.192.138/gisd/100_worst.php (accessed on 1 July 2021).
- CEE. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Com. 1992, 206, 7–50. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A31992L0043 (accessed on 1 July 2021).
- Almodóvar, A.; Nicola, G.G.; Ayllón, D.; Elvira, B. Global warming threatens the persistence of Mediterranean brown trout. Glob. Chang. Biol. 2012, 18, 1549–1560. [Google Scholar] [CrossRef] [Green Version]
- Ayllón, D.; Railsback, S.F.; Harvey, B.C.; García Quirós, I.; Nicola, G.G.; Elvira, B.; Almodóvar, A. Mechanistic simulations predict that thermal and hydrological effects of climate change on Mediterranean trout cannot be offset by adaptive behaviour, evolution, and increased food production. Sci. Total Environ. 2019, 693, 133648. [Google Scholar] [CrossRef] [PubMed]
- Leroy, G.; Carroll, E.L.; Bruford, M.W.; DeWoody, J.A.; Strand, A.; Waits, L.; Wang, J. Next-generation metrics for monitoring genetic erosion within populations of conservation concern. Evol. Appl. 2018, 11, 1066–1083. [Google Scholar] [CrossRef] [Green Version]
- Moritz, C. Defining ‘Evolutionarily Significant Units’. Trends Ecol. Evol. 1994, 9, 373–375. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Luikart, G.; Aitken, S.N. Conservation and the Genetics of Populations; Wiley-Blackwell: Oxford, UK, 2016. [Google Scholar]
- Palsbøll, P.J.; Bérubé, M.; Allendorf, F.W. Identification of management units using population genetic data. Trends Ecol. Evol. 2007, 22, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Funk, W.C.; McKay, J.K.; Hohenlohe, P.A.; Allendorf, F.W. Harnessing genomics for delineating conservation units. Trends Ecol. Evol. 2012, 27, 489–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willi, Y.; Kristensen, T.N.; Sgrò, C.M.; Weeksd, A.R.; Ørstedb, M.; Hoffmannd, A.A. Conservation genetics as a management tool: The five best-supported paradigms to assist the management of threatened species. Proc. Natl. Acad. Sci. USA 2022, 119, e2105076119. [Google Scholar] [CrossRef]
- Zachos, F.E.; Apollonio, M.; Bärmann, E.V.; Festa-bianchet, M.; Göhlich, U.; Christian, J.; Haring, E.; Kruckenhauser, L.; Lovari, S.; Mcdevitt, A.D.; et al. Species inflation and taxonomic artefacts—A critical comment on recent trends in mammalian classificatio. Mamm. Biol. 2013, 78, 1–6. [Google Scholar] [CrossRef]
- Splendiani, A.; Palmas, F.; Sabatini, A.; Caputo Barucchi, V. The name of the trout: Considerations on the taxonomic status of the Salmo trutta L., 1758 complex (Osteichthyes: Salmonidae) in Italy. Eur. Zool. J. 2019, 86, 432–442. [Google Scholar] [CrossRef] [Green Version]
- Serrote, C.M.L.; Reiniger, L.R.S.; Silva, K.B.; Rabaiolli, S.M.S.; Stefanel, C.M. Determining the Polymorphism Information Content of a molecular marker. Gene 2020, 726, 144175. [Google Scholar] [CrossRef] [PubMed]
- Livi, S.; De Innocentiis, S.; Longobardi, A.; Cataudella, S.; Tancioni, L.; Rampacci, M.; Marino, G. Genetic structure of Barbus spp. populations in the Marches Region of central Italy and its relevance to conservation actions. J. Fish Biol. 2013, 82, 806–826. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.R.; Milana, V.; Pulcini, D.; Cataudella, S.; Martinoli, M.; Tancioni, L. An integrated genetic and morphological approach to clarify the conservation status of the threatened Italian endemic species Alburnus albidus (Cypriniformes: Cyprinidae). Hydrobiologia 2016, 770, 73–87. [Google Scholar] [CrossRef]
- Rossi, A.R.; Petrosino, G.; Crescenzo, S.; Milana, V.; Talarico, L.; Martinoli, M.; Rakaj, A.; Lorenzoni, M.; Carosi, A.; Ciuffardi, L.; et al. Phylogeography and population structure of Squalius lucumonis: A baseline for conservation of an Italian endangered freshwater fish. J. Nat. Cons. 2021, 64, 126085. [Google Scholar] [CrossRef]
- Gratton, P.; Allegrucci, G.; Sbordoni, V.; Gandolfi, A. The evolutionary jigsaw puzzle of the surviving trout (Salmo trutta L. complex) diversity in the italian region. A multilocus Bayesian approach. Mol. Phylogenet. Evol. 2014, 79, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Fabiani, A.; Gratton, P.; Zappes, I.A.; Seminara, M.; D’Orsi, A.; Sbordoni, V.; Allegrucci, G. Investigating the genetic structure of trout from the Garden of Ninfa (central Italy): Suggestions for conservation and management. Fish. Manag. Ecol. 2018, 25, 1–11. [Google Scholar] [CrossRef]
- Sanz, N.; Araguas, R.M.; Fernández-Cebrián, R.; Lobón-Cerviá, J. Factors modelling population structure in brown trout Salmo trutta L.: Genetic monitoring of populations in Esva River (northwestern Spain). Hydrobiologia 2019, 837, 117–131. [Google Scholar] [CrossRef]
- Splendiani, A.; Fioravanti, T.; Ruggeri, P.; Giovannotti, M.; Carosi, A.; Marconi, M.; Lorenzoni, M.; Righi, T.; Nisi, P.; Vincenzo, C.; et al. Life history and genetic characterisation of sea trout Salmo trutta in the Adriatic Sea. Freshw. Biol. 2019, 65, 460–473. [Google Scholar] [CrossRef]
- Berrebi, P.; Caputo Barucchi, V.; Splendiani, A.; Muracciole, S.; Sabatini, A.; Palmas, F.; Tougard, C.; Arculeo, M.; Maric, S. Brown trout (Salmo trutta L.) high genetic diversity around the Tyrrhenian Sea as revealed by nuclear and mitochondrial markers. Hydrobiologia 2019, 826, 209–231. [Google Scholar] [CrossRef]
- McMeel, O.M.; Hoey, E.M.; Ferguson, A. Partial nucleotide sequences, and routine typing by polymerase chain reaction-restriction fragment length polymorphism, of the brown trout (Salmo trutta) lactate dehydrogenase, LDH-C1*90 and *100 alleles. Mol. Ecol. 2001, 10, 29–34. [Google Scholar] [CrossRef]
- Kaufman, J. Unfinished Business: Evolution of the MHC and the Adaptive Immune System of Jawed Vertebrates. Annu. Rev. Immunol. 2018, 36, 383–409. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.M.; Skaala, Ø.; Jensen, L.F.; Bekkevold, D.; Mensberg, K.-L.D. Gene flow, effective population size and selection at major histocompatibility complex genes: Brown trout in the Hardanger Fjord, Norway. Mol. Ecol. 2007, 16, 1413–1425. [Google Scholar] [CrossRef] [Green Version]
- Schenekar, T.; Weiss, S. Selection and genetic drift in captive versus wild populations: An assessment of neutral and adaptive (MHC-linked) genetic variation in wild and hatchery brown trout (Salmo trutta) populations. Conserv. Genet. 2017, 18, 1011–1022. [Google Scholar] [CrossRef] [Green Version]
- Talarico, L.; Marta, S.; Rossi, A.R.; Crescenzo, S.; Petrosino, G.; Martinoli, M.; Tancioni, L. Balancing selection, genetic drift, and human mediated-introgression interplay to shape MHC (functional) diversity in Mediterranean brown trout. Ecol. Evol. 2021, 11, 10026–10041. [Google Scholar] [CrossRef] [PubMed]
- Caputo, V.; Giovannotti, M.; Nisi Cerioni, P.; Caniglia, M.L.; Splendiani, A. Genetic diversity of brown trout in central Italy. J. Fish Biol. 2004, 65, 403–418. [Google Scholar] [CrossRef]
- Palombo, V.; De Zio, E.; Salvatore, G.; Esposito, S.; Iaffaldano, N.; D’Andrea, M. Genotyping of two Mediterranean trout populations in Central-Southern Italy for conservation purposes using a rainbow-trout-derived SNP array. Animals 2021, 11, 1803. [Google Scholar] [CrossRef] [PubMed]
- Splendiani, A.; Ruggeri, P.; Giovannotti, M.; Pesaresi, S.; Occhipinti, G.; Fioravanti, T.; Lorenzoni, M.; Nisi Cerioni, P.; Caputo Barucchi, V. Alien brown trout invasion of the Italian peninsula: The role of geological, climate and anthropogenic factors. Biol. Invasions 2016, 18, 2029–2044. [Google Scholar] [CrossRef]
- Gratton, P.; Allegrucci, G.; Gandolfi, A.; Sbordoni, V. Genetic differentiation and hybridization in two naturally occurring sympatric trout Salmo spp. forms from a small karstic lake. J. Fish Biol. 2013, 82, 637–657. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.R.; Petrosino, G.; Milana, V.; Martinoli, M.; Rakaj, A.; Tancioni, L.; Petrosino, G.; Milana, V.; Martinoli, M.; Rakaj, A.; et al. Genetic identification of native populations of Mediterranean brown trout Salmo trutta L. complex (Osteichthyes: Salmonidae) in central Italy. Eur. Zool. J. 2019, 86, 424–431. [Google Scholar] [CrossRef] [Green Version]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef] [PubMed]
- Cortey, M.; García-Marín, J.L. Evidence for phylogeographically informative sequence variation in the mitochondrial control region of Atlantic brown trout. J. Fish Biol. 2002, 60, 1058–1063. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Estoup, A.; Presa, P.; Krieg, F.; Vaiman, D.; Guyomard, R. (CT)n and (GT)n microsatellites: A new class of genetic markers for Salmo trutta L. (brown trout). Heredity 1993, 71, 488–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, P.T.; Hamilton, L.C.; McConnell, S.K.; Wright, J.M. Rapid analysis of genetic variation in Atlantic salmon (Salmo salar) by PCR multiplexing of dinucleotide and tetranucleotide microsatellites. Can. J. Fish. Aquat. Sci. 1996, 53, 2292–2298. [Google Scholar] [CrossRef]
- Slettan, A.; Olsaker, I.; Lie, Ø. Atlantic salmon, Salmo salar, microsatellites at the SSOSL25, SSOSL85, SSOSL311, SSOSL417 loci. Anim. Genet. 1995, 26, 277–285. [Google Scholar] [CrossRef]
- Cairney, M.; Taggart, J.B.; Høyheim, B. Characterization of microsatellite and minisatellite loci in Atlantic salmon (Salmo salar L.) and cross-species amplification in other salmonids. Mol. Ecol. 2000, 9, 2175–2178. [Google Scholar] [CrossRef] [PubMed]
- Paterson, S.; Piertney, S.B.; Knox, D.; Gilbey, J.; Verspoor, E. Characterization and PCR multiplexing of novel highly variable tetranucleotide Atlantic salmon (Salmo salar L.) microsatellites. Mol. Ecol. Notes 2004, 4, 160–162. [Google Scholar] [CrossRef]
- Rexroad, C.E.; Coleman, R.L.; Hershberger, W.K.; Killefer, J. Rapid communication: Thirty-eight polymorphic microsatellite markers for mapping in rainbow trout. J. Anim. Sci. 2002, 80, 541–542. [Google Scholar] [CrossRef] [PubMed]
- King, T.L.; Eackles, M.S.; Letcher, B.H. Microsatellite DNA markers for the study of Atlantic salmon (Salmo salar) kinship, population structure, and mixed-fishery analyses. Mol. Ecol. Notes 2005, 5, 130–132. [Google Scholar] [CrossRef]
- Scribner, K.T.; Gust, J.R.; Fields, R.L. Isolation and characterization of novel salmon microsatellite loci: Cross-species amplification and population genetic applications. Can. J. Fish. Aquat. Sci. 1996, 53, 833–841. [Google Scholar] [CrossRef]
- Thorsen, J.; Zhu, B.; Frengen, E.; Osoegawa, K.; de Jong, P.J.; Koop, B.F.; Davidson, W.S.; Høyheim, B. A highly redundant BAC library of Atlantic salmon (Salmo salar): An important tool for salmon projects. BMC Genom. 2005, 6, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousset, F. GENEPOP’007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Chapuis, M.P.; Estoup, A. Microsatellite null alleles and estimation of population differentiation. Mol. Biol. Evol. 2007, 24, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Res. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Adamack, A.T.; Gruber, B. PopGenReport: Simplifying basic population genetic analyses in R. Methods Ecol. Evol. 2014, 5, 384–387. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4. Available online: https://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 20 September 2021).
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. CLUMPAK: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.L.; Liu, J.X. StructureSelector: A web-based software to select and visualize the optimal number of clusters using multiple methods. Mol. Ecol. Resour. 2018, 18, 176–177. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Piry, S.; Alapetite, A.; Cornuet, J.M.; Paetkau, D.; Baudouin, L.; Estoup, A. GENECLASS2: A Software for genetic assignment and first-generation migrant detection. J. Hered. 2004, 95, 536–539. [Google Scholar] [CrossRef]
- Rannala, B.; Mountain, J.L. Detecting immigration by using multilocus genotypes. Proc. Natl. Acad. Sci. USA 1997, 94, 9197–9201. [Google Scholar] [CrossRef] [Green Version]
- Wang, J. A new method for estimating effective population sizes from a single sample of multilocus genotypes. Mol. Ecol. 2009, 18, 2148–2164. [Google Scholar] [CrossRef]
- Serbezov, D.; Bernatchez, L.; Olsen, E.M.; VØllestad, L.A. Mating patterns and determinants of individual reproductive success in brown trout (Salmo trutta) revealed by parentage analysis of an entire stream living population. Mol. Ecol. 2010, 19, 3193–3205. [Google Scholar] [CrossRef]
- Piry, S.; Luikart, G.; Cornuet, J.-M. BOTTLENECK: A computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Splendiani, A.; Giovannotti, M.; Nisi Cerioni, P.; Caniglia, M.L.; Caputo, V. Phylogeographic inferences on the native brown trout mtDNA variation in central Italy. Ital. J. Zool. 2006, 73, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Delling, B.; Sabatini, A.; Muracciole, S.; Tougard, C.; Berrebi, P. Morphologic and genetic characterization of Corsican and Sardinian trout with comments on Salmo taxonomy. Knowl. Manag. Aquat. Ecosyst. 2020, 421, 21. [Google Scholar] [CrossRef]
- Thaulow, J.; Borgstrøm, R.; Heun, M. Brown trout population structure highly affected by multiple stocking and river diversion in a high mountain national park. Conserv. Genet. 2013, 14, 145–158. [Google Scholar] [CrossRef]
- Torterotot, J.B.; Perrier, C.; Bergeron, N.E.; Bernatchez, L. Influence of forest road culverts and waterfalls on the fine-scale distribution of brook trout genetic diversity in a boreal watershed. Trans. Am. Fish. Soc. 2014, 143, 1577–1591. [Google Scholar] [CrossRef]
- Almodóvar, A.; Nicola, G.G.; Elvira, B.; García-Marín, J.L. Introgression variability among Iberian brown trout Evolutionary Significant Units: The influence of local management and environmental features. Freshw. Biol. 2006, 51, 1175–1187. [Google Scholar] [CrossRef]
- Consuegra, S.; Phillips, N.; Gajardo, G.; Garcia de Leaniz, C. Winning the invasion roulette: Escapes from fish farms increase admixture and facilitate establishment of non-native rainbow trout. Evol. Appl. 2011, 4, 660–671. [Google Scholar] [CrossRef] [PubMed]
- Rius, M.; Darling, J.A. How important is intraspecific genetic admixture to the success of colonising populations? Trends Ecol. Evol. 2014, 29, 233–242. [Google Scholar] [CrossRef]
- Berrebi, P.; Horvath, A.; Splendiani, A.; Palm, S.; Bernaś, R. Genetic diversity of domestic brown trout stocks in Europe. Aquaculture 2021, 544, 737043. [Google Scholar] [CrossRef]
- Hänfling, B. Understanding the establishment success of non-indigenous fishes: Lessons from population genetics. J. Fish Biol. 2007, 71 (Suppl. D), 115–135. [Google Scholar] [CrossRef]
- Miller, L.M.; Close, T.; Kapuscinski, R. Lower fitness of hatchery and hybrid rainbow trout compared to naturalized populations in Lake Superior tributaries. Mol. Ecol. 2004, 13, 3379–3388. [Google Scholar] [CrossRef]
- Hansen, M.M.; Fraser, D.J.; Meier, K.; Mensberg, K.L.D. Sixty years of anthropogenic pressure: A spatio-temporal genetic analysis of brown trout populations subject to stocking and population declines. Mol. Ecol. 2009, 18, 2549–2562. [Google Scholar] [CrossRef]
- Wollebaek, J.; Knut, H.R.; Brabrand, A.; Heggenes, J. Interbreeding of genetically distinct native brown trout (Salmo trutta) populations designates offspring fitness. Aquaculture 2012, 356, 158–168. [Google Scholar] [CrossRef]
- Garcia de Leaniz, C.; Fleming, I.A.; Einum, S.; Verspoor, E.; Jordan, W.C.; Consuegra, S.; Aubin-Horth, N.; Laius, D.; Letcher, B.H.; Youngson, A.F.; et al. A critical review of adaptive genetic variation in Atlantic salmon: Implications for conservation. Biol. Rev. 2007, 82, 173–211. [Google Scholar] [CrossRef]
- Garcia-Marín, J.L.; Araguas, R.M.; Vera, M.; Sanz, N. Understanding the Brown Trout Population Genetic Structure in the Iberian Peninsula. In Brown Trout: Biology, Ecology and Management; Lobón Cerviá, J., Sanz, N., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2018; pp. 103–126. [Google Scholar]
- White, S.L.; Miller, W.L.; Dowell, S.A.; Bartron, M.L.; Wagner, T. Limited hatchery introgression into wild brook trout (Salvelinus fontinalis) populations despite reoccurring stocking. Evol. Appl. 2018, 11, 1567–1581. [Google Scholar] [CrossRef]
- Hansen, M.M.; Ruzzante, D.E.; Nielsen, E.E.; Mensberg, K.L.D. Microsatellite and mitochondrial DNA polymorphism reveals life-history dependent interbreeding between hatchery and wild brown trout (Salmo trutta L.). Mol. Ecol. 2000, 9, 583–594. [Google Scholar] [CrossRef]
- Querci, G.; Pecchioli, E.; Leonzio, C.; Frati, F.; Nardi, F. Molecular characterization and hybridization in Salmo (trutta) macrostigma morphotypes from Central Italy. Hydrobiologia 2013, 702, 191–200. [Google Scholar] [CrossRef]
- Mezzera, M.; Largiadèr, C.R. Evidence for selective angling of introduced trout and their hybrids in a stocked brown trout population. J. Fish Biol. 2001, 59, 287–301. [Google Scholar] [CrossRef]
- Hargrove, J.S.; Kazyak, D.C.; Lubinski, B.A.; Rogers, K.M.; Bowers, O.K.; Fesenmyer, K.A.; Habera, J.W.; Henegar, J. Landscape and stocking effects on population genetics of Tennessee Brook Trout. Conserv. Genet. 2021; in press. [Google Scholar] [CrossRef]
- Saha, A.; Andersson, A.; Kurland, S.; Keehnen, N.L.P.; Kutschera, V.E.; Hössjer, O.; Ekman, D.; Karlsson, S.; Kardos, M.; Ståhl, G.; et al. Whole-genome resequencing confirms reproductive isolation between sympatric demes of brown trout (Salmo trutta) detected with allozymes. Mol. Ecol. 2022, 31, 498–511. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pop | River Catchment | Drainage Basin (Outlet) | Lat° (N) | Lon° (E) | Elev. (m) | Water T° (± SE) | Fish Assemblage | River Substrate | Protected Area |
|---|---|---|---|---|---|---|---|---|---|
| CAR | Carpello | Liri-Garigliano (Tyrrhenian Sea) | 41.70 | 13.68 | 293 | 11.4 (±0.2) | Gasterosteus aculeatus | Gravel and aquatic macrophytes | SAC, Regional Natural Reserve |
| MEL | Melfa | Liri-Garigliano (Tyrrhenian Sea) | 41.70 | 13.91 | 1159 | 8.8 (±0.1) | - | Boulder, pebbles, gravel | SAC, National Park |
| RAP | Rapido | Liri-Garigliano (Tyrrhenian Sea) | 41.56 | 13.88 | 154 | 13.6 (±0.0) | - | Pebbles, gravel | - |
| SCR | Santa Croce | Santa Croce (Tyrrhenian Sea) | 41.29 | 13.71 | 37 | 13.7 (±0.2) | Anguilla anguilla, Lampetra planeri, Sarmarutilus rubilio, Squalius squalus | Gravel and aquatic macrophytes | SAC, close to Regional Park |
| SIM1 | Simbrivio | Tiber (Tyrrhenian Sea) | 41.94 | 13.24 | 794 | 11.3 (±0.1) | - | Boulder, pebbles, gravel | SAC, Regional Park |
| SIM2 | Simbrivio | Tiber (Tyrrhenian Sea) | 41.92 | 13.22 | 701 | 12.0 (±0.2) | - | Pebbles, gravel | SAC, Regional Park |
| TRO | Tronto | Tronto (Adriatic Sea) | 42.73 | 13.26 | 693 | 10.7 (±0.1) | - | Pebbles, gravel | - |
| CR | LDH-C1 | 11 STR loci | MHC-DAB | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pop | N (NMHC) | H (Hp) | Hr | Hd (±SE) | π% (±SE) | *90 | Ho | He | A (Ap) | Ar | Ho (±SE) | He (±SE) | Ne (95% CI) | A (Ap) | Ar | Ho | He | |||
| CAR | 30 (28) | 1 (1) | 1.00 | - - | - - | 0.13 | 0.27 | 0.24 | 7.09 (1.45) | 4.92 | 0.47 (0.09) | 0.52 (0.10) | 40 (24–71) | 13 (6) | 8.35 | 0.86 | 0.81 | |||
| MEL | 30 (27) | 2 (0) | 1.86 | 0.19 (0.09) | 0.034 (0.05) | 0.90 | 0.20 | 0.18 | 6.18 (0.45) | 5.21 | 0.69 (0.06) | 0.73 (0.05) | 34 (21–62) | 12 (2) | 8.31 | 0.81 | 0.84 | |||
| RAP | 30 (25) | 3 (0) | 2.72 | 0.58 (0.04) | 0.471 (0.289) | 0.52 | 0.43 | 0.51 | 7.55 (0.73) | 5.95 | 0.78 (0.08) | 0.73 (0.07) | 17 (9–35) | 12 (5) | 8.47 | 0.88 | 0.83 | |||
| SCR | 30 (26) | 2 (1) | 1.99 | 0.37 (0.08) | 0.408 (0.258) | 0.05 | 0.10 | 0.10 | 5.09 (1.00) | 3.83 | 0.41 (0.08) | 0.41 (0.07) | 34 (22–60) | 7 (3) | 5.28 | 0.65 | 0.75 | |||
| SIM1 | 16 (15) | 3 (0) | 2.88 | 0.43 (0.13) | 0.501 (0.314) | 0.28 | 0.44 | 0.42 | 8.36 (0.73) | 6.93 | 0.74 (0.06) | 0.74 (0.05) | 45 (24–95) | 15 (3) | 10.80 | 0.60 | 0.84 | |||
| SIM2 | 14 (12) | 3 (1) | 3.00 | 0.54 (0.11) | 0.591 (0.364) | 0.61 | 0.36 | 0.49 | 8.09 (0.73) | 6.82 | 0.68 (0.05) | 0.75 (0.06) | 88 (35–∞) | 14 (5) | 11.67 | 0.75 | 0.94 | |||
| TRO | 25 (23) | 6 (2) | 5.12 | 0.77 (0.05) | 0.677 (0.394) | 0.44 | 0.40 | 0.50 | 10.09 (1.73) | 7.44 | 0.75 (0.05) | 0.77 (0.05) | 54 (32–104) | 20 (11) | 12.89 | 0.91 | 0.92 | |||
| Grouping | Hierarchical Level | nDNA | mtDNA | ||||
|---|---|---|---|---|---|---|---|
| 11 STR | MHC-DAB | CR (K2P) | |||||
| Variation | p | Variation | p | Variation | p | ||
| One group | Among populations | 21.96 | 0.000 | 15.08 | 0.000 | 62.77 | 0.000 |
| Within populations | 78.04 | 0.000 | 84.92 | 0.000 | 37.23 | 0.000 | |
| Four groups: drainage basins CAR/MEL/RAP vs. SCR vs. SIM1/SIM2 vs. TRO | Among groups | 4.61 | 0.273 | 1.05 | 0.393 | −28.78 | 0.899 |
| Among populations within groups | 18.16 | 0.000 | 14.23 | 0.000 | 89.13 | 0.000 | |
| Within populations | 77.23 | 0.000 | 84.72 | 0.000 | 39.64 | 0.000 | |
| Two groups: mountain vs. lowland CAR/RAP/SCR vs. MEL/SIM1/SIM2/TRO | Among groups | 2.06 | 0.231 | −0.13 | 0.574 | 3.53 | 0.463 |
| Among populations within groups | 20.56 | 0.000 | 15.16 | 0.000 | 59.78 | 0.000 | |
| Within populations | 77.38 | 0.000 | 84.96 | 0.000 | 36.69 | 0.000 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, A.R.; Talarico, L.; Petrosino, G.; Crescenzo, S.; Tancioni, L. Conservation Genetics of Mediterranean Brown Trout in Central Italy (Latium): A Multi-Marker Approach. Water 2022, 14, 937. https://doi.org/10.3390/w14060937
Rossi AR, Talarico L, Petrosino G, Crescenzo S, Tancioni L. Conservation Genetics of Mediterranean Brown Trout in Central Italy (Latium): A Multi-Marker Approach. Water. 2022; 14(6):937. https://doi.org/10.3390/w14060937
Chicago/Turabian StyleRossi, Anna Rita, Lorenzo Talarico, Gerardo Petrosino, Simone Crescenzo, and Lorenzo Tancioni. 2022. "Conservation Genetics of Mediterranean Brown Trout in Central Italy (Latium): A Multi-Marker Approach" Water 14, no. 6: 937. https://doi.org/10.3390/w14060937
APA StyleRossi, A. R., Talarico, L., Petrosino, G., Crescenzo, S., & Tancioni, L. (2022). Conservation Genetics of Mediterranean Brown Trout in Central Italy (Latium): A Multi-Marker Approach. Water, 14(6), 937. https://doi.org/10.3390/w14060937