
Effects of Combined Nitrogen Deficient and Mixotrophic (+Glucose) Culture on the Lipid Accumulation of Parachlorella Kessleri TY



Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Strain and Cultivation
2.2. Determination of Biomass and Biomass Productivity
2.3. Determination of Pigment Contents
2.4. Determination of Chlorophyll Fluorescence
2.5. Determination of Neutral Lipid Content
2.6. Determination of Total Lipid Content and Fatty Acid Composition
2.6.1. Determination of Total Lipid Content
2.6.2. Determination of Fatty Acid Composition
2.7. Determination of Total Carbohydrate Content
2.8. Determination of Total Protein Content
2.9. Statistical Analysis
3. Results
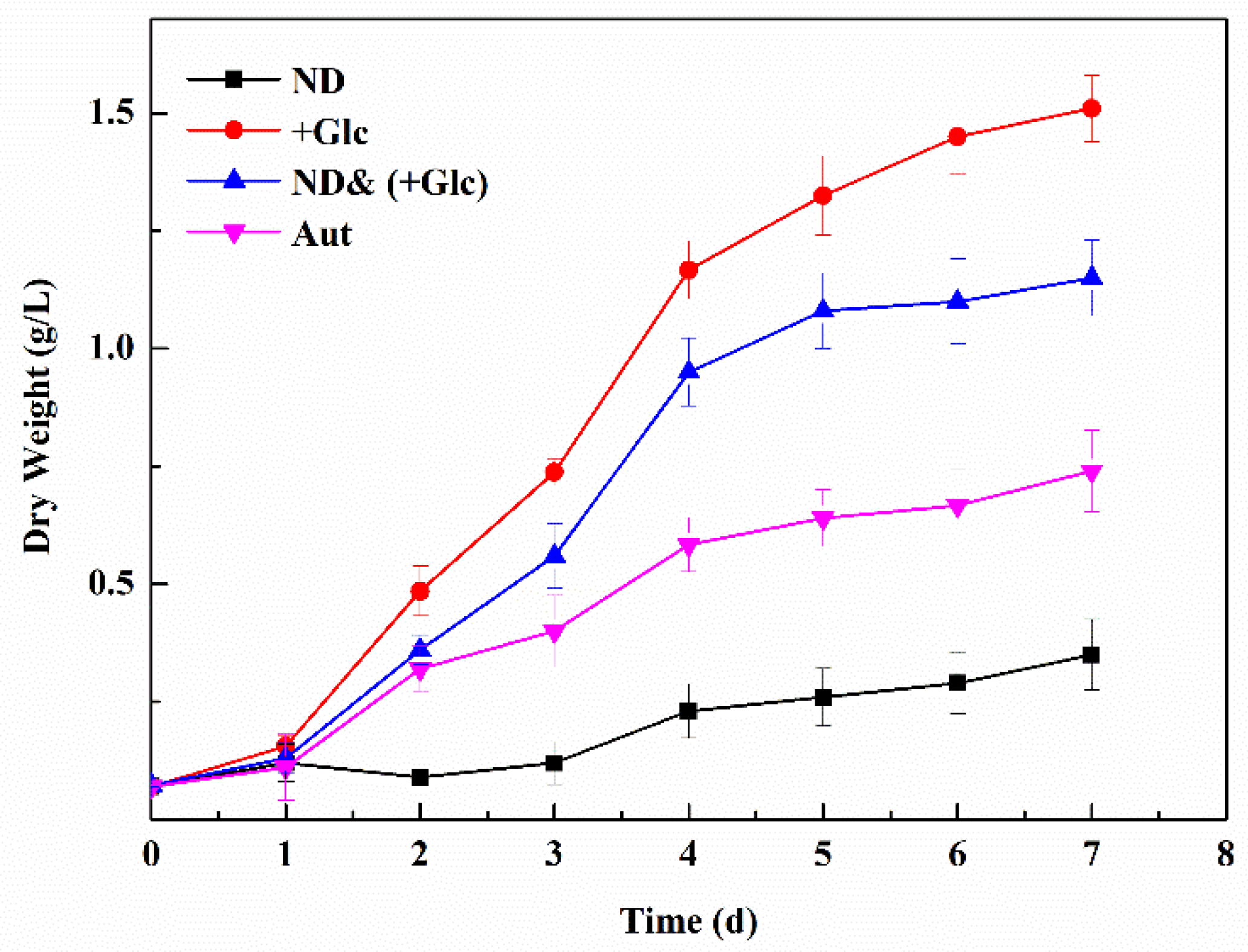
3.1. Analysis of Changes in Growth
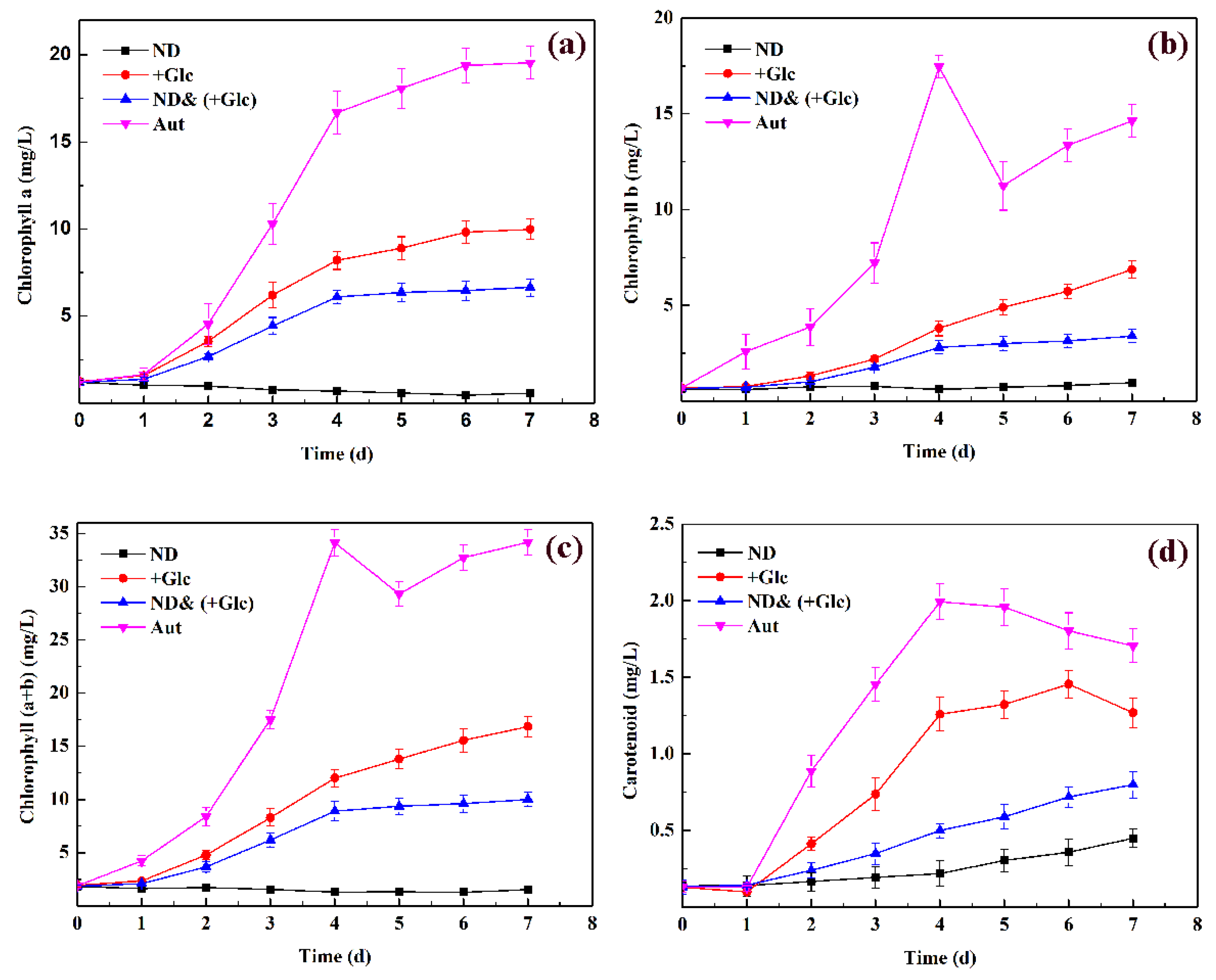
3.2. Analysis of Pigment Content
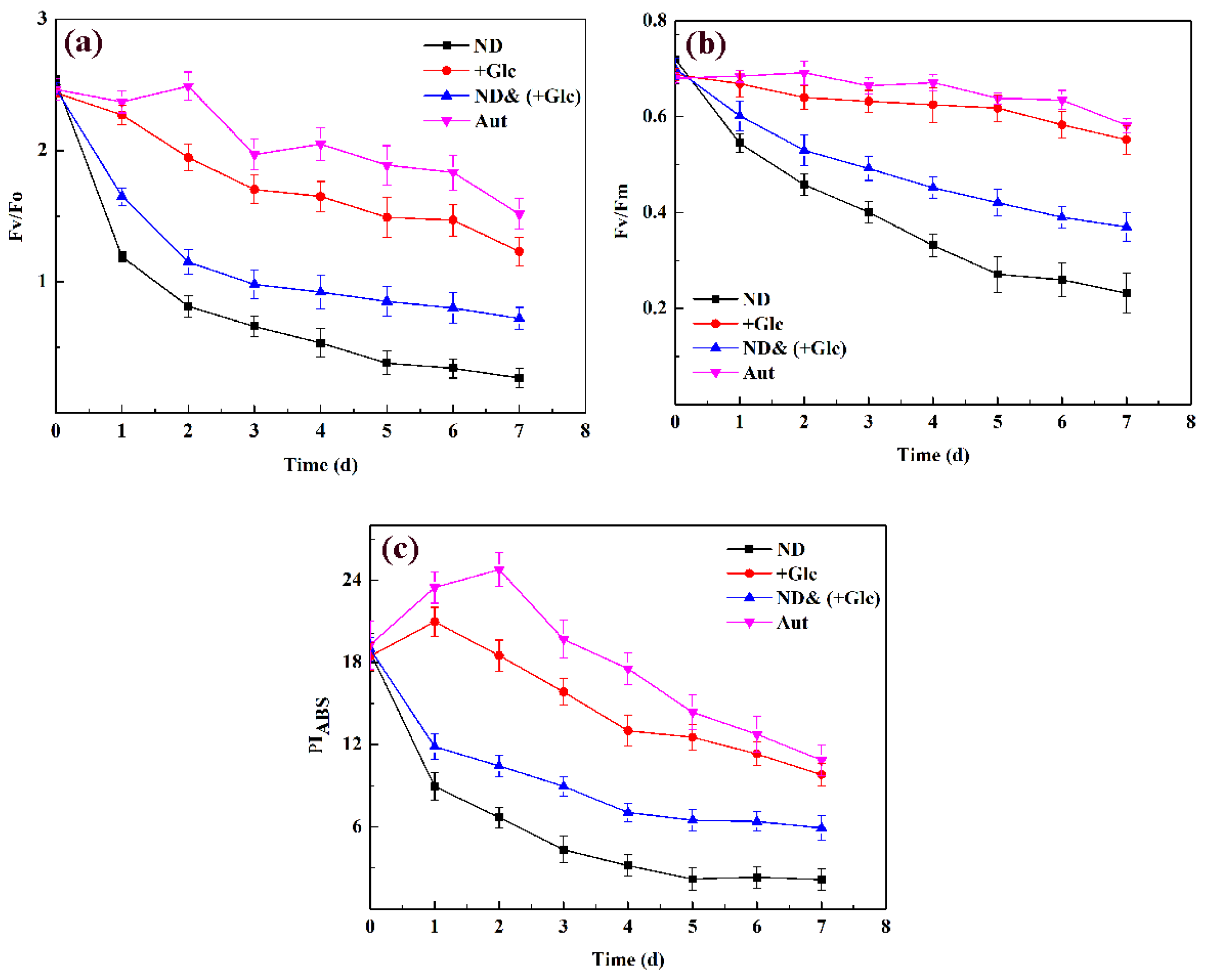
3.3. Analysis of Chlorophyll Fluorescence Characteristics
3.4. Analysis of Neutral Lipid Content
3.5. Analysis of Total Lipid Content and Fatty Acid Composition
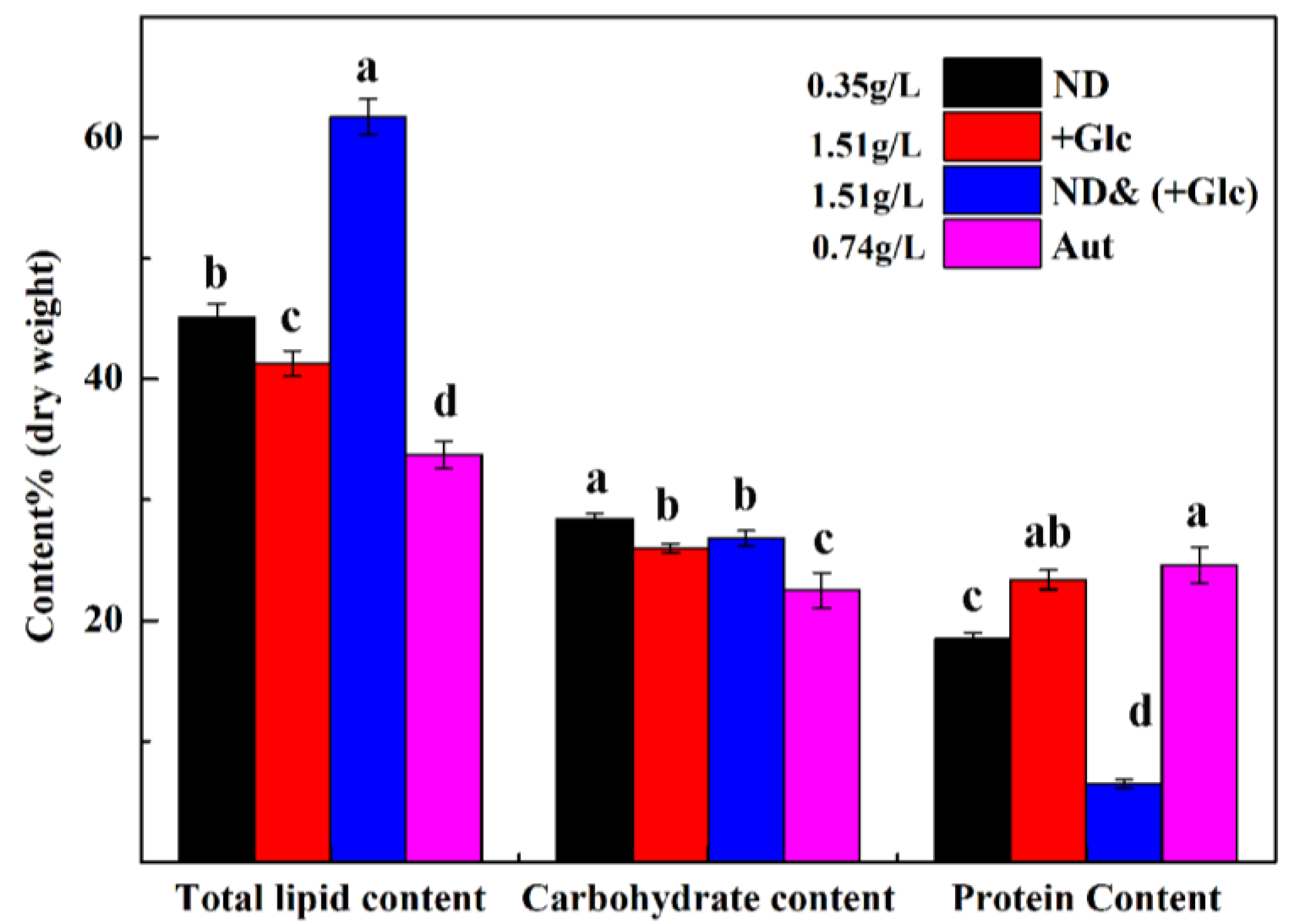
3.5.1. Analysis of Total Lipid Content
3.5.2. Analysis of Fatty Acid Composition
3.6. Analysis of Total Carbohydrate Content
3.7. Analysis of Total Protein Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Swanson, K.J.; Madden, M.C.; Ghio, A.J. Biodiesel exhaust: The need for health effects research. Environ. Health Perspect. 2007, 115, 496–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapuerta, M.; Armas, O.; Rodríguez-Fernández, J. Effect of biodiesel fuels on diesel engine emissions. Prog. Energy Combus. Sci. 2008, 34, 198–223. [Google Scholar] [CrossRef]
- Song, P.P.; Jia, Y.; Li, Q. Effects of nitrogen deficiency on growth and lipid and protein content of Isochrysis galbana. Technol. Mark. 2015, 22, 113–116. [Google Scholar] [CrossRef]
- Pancha, I.; Chokshi, K.; George, B.; Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Nitrogen stress triggered biochemical and morphological changes in the microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2014, 156, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Rios, L.F.; Klein, B.C.; Luz, L.F.J.; Filho, R.M.; Maciel, M.R.F. Nitrogen starvation for lipid accumulation in the microalga species Desmodesmus sp. Appl. Biochem. Biotechnol. 2015, 175, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Siaut, M.; Cuine, S.; Cagnon, C.; Fessler, B.; Nguyen, M.; Carrier, P.; Beyly, A.; Beisson, F.; Triantaphylidès, C.; Li-Beisson, Y.; et al. Oil accumulation in the model green alga Chlamydomonas reinhardtii: Characterization, variability between common laboratory strains and relationship with starch reserves. BMC Biotechnol. 2011, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.N.; Huang, W.; Xu, J.; Wang, Z.M.; Xu, J.L.; Yuan, Z.H. Metabolic changes of starch and lipid triggered by nitrogen starvation in the microalga Chlorella zofingiensis. Bioresour. Technol. 2014, 152, 292–298. [Google Scholar] [CrossRef]
- Lin, T.S.; Wu, J.Y. Effect of carbon sources on growth and lipid accumulation of newly isolated microalgae cultured under mixotrophic condition. Bioresour. Technol. 2015, 184, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Wan, M.X.; Liu, P.; Xia, J.L.; Rosenberg, J.N.; Oyler, G.A.; Betenbaugh, M.J.; Nie, Z.Y.; Qiu, G.Z. The effect of mixotrophy on microalgal growth, lipid content, and expression levels of three pathway genes in Chlorella sorokiniana. Appl. Microbiol. Biotechnol. 2011, 91, 835–844. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Bedaiwy, M.Y.; Osman, M.E.; Ismail, M.M. Mixotrophic and heterotrophic growth of some microalgae using extract of fungal-treated wheat bran. Int. J. Recycl. Org. Waste Agric. 2012, 1, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.F.; Feng, J.; Lv, J.P.; Liu, Q.; Nan, F.R.; Liu, X.D.; Xie, S.L. Physiological changes of Parachlorella Kessleri TY02 in lipid accumulation under nitrogen stress. Int. J. Environ. Res. Public Health 2019, 16, 1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Z.Z.; Wang, J.; Yu, X.B. Effects of fed-batch cultivation and nitrogen deficiency on lipid yield of Chlorella vulgaris. J. Anhui Agric. Sci. 2012, 40, 2453–2456. [Google Scholar] [CrossRef]
- Chen, Y.M. The Invention Relates to a Culture Method for Increasing the Yield of Microalgae Biomass and Lipid. A Patent for Invention. Chinese Patent No. CN103484372A, 2014. [Google Scholar]
- Cho, D.H.; Ramanan, R.; Kim, B.H.; Lee, J.; Kim, S.; Yoo, C.; Choi, G.G.; Oh, H.M.; Kim, H.S. Novel approach for the development of axenic microalgal cultures from environmental samples. J. Phycol. 2013, 49, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.P.; Guo, J.Y.; Feng, J.; Liu, Q.; Xie, S.L. Effect of sulfate ions on growth and pollutants removal of self-flocculating microalga Chlorococcum sp. GD in synthetic municipal wastewater. Bioresour. Technol. 2017, 234, 289–296. [Google Scholar] [CrossRef]
- Mera, R.; Torres, E.; Abalde, J. Effects of sodium sulfate on the freshwater microalga Chlamydomonas moewusii: Implications for the optimization of algal culture media. J. Phycol. 2016, 52, 75–88. [Google Scholar] [CrossRef]
- Markou, G.; Muylaert, K. Effect of light intensity on the degree of ammonia toxicity on PSII activity of Arthrospira platensis and Chlorella vulgaris. Bioresour. Technol. 2016, 216, 453–461. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, C.; Song, L.; Sommerfeld, M.; Hua, Q. A high throughput Nile red method for quantitative measurement of neutral lipids in microalgae. J. Microbiol. Methods 2015, 77, 41–47. [Google Scholar] [CrossRef]
- Bohnenberger, J.E.; Crossetti, L.O. Influence of temperature and nutrient content on lipid production in freshwater microalgae cultures. An. Acad. Bras. Ciênc. 2014, 86, 1239–1248. [Google Scholar] [CrossRef] [Green Version]
- Thang, D.V.; Faruq, A.; Thomas-Hall, S.R.; Simon, Q.; Ekaterina, N.; Schenk, P.M. High protein and high lipid-producing microalgae from northern Australia as potential feedstock for animal feed and biodiesel. Front. Bioeng. Biotechnol. 2015, 3, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Huang, J.; Sun, Z.; Zhong, Y.; Jiang, Y.; Chen, F. Differential lipid and fatty acid profiles of photoautotrophic and heterotrophic Chlorella zofingiensis: Assessment of algal oils for biodiesel production. Bioresour. Technol. 2011, 102, 106–110. [Google Scholar] [CrossRef]
- Li, H.S. Determination of Total Protein and Carbohydrate Content. In Principles and Techniques of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000; pp. 184–199. [Google Scholar]
- Prajapati, S.K.; Kaushik, P.; Malik, A.; Vijay, V.K. Phycoremediation and biogas potential of native algal isolates from soil and wastewater. Bioresour. Technol. 2013, 135, 232–238. [Google Scholar] [CrossRef]
- He, Q.N.; Yang, H.J.; Hu, C.X. Effects of temperature and its combination with high light intensity on lipid production of Monoraphidium dybowskii Y2 from semi-arid desert areas. Bioresour. Technol. 2018, 265, 407–414. [Google Scholar] [CrossRef]
- Guo, Y.N. Optimizing Cultivation and Mechanism of Lipid Accumulation Research in Chlorococcum sphacosum GD. Master’s Dissertation, Shanxi University, Taiyuan, China, 2017. [Google Scholar]
- Deng, D.X.; Deng, D.Y.; Liu, J.P.; Cheng, K.; Xu, Y.J. Field trial multiple comparison results alphabetic difference counting method and its letter abbreviation. Seeds 2018, 37, 131–132. [Google Scholar] [CrossRef]
- Girard, J.M.; Roy, M.L.; Hafsa, M.B.; Gagnon, J.; Faucheux, N.; Heitz, M.; Tremblay, L.; Deschênes, J.S. Mixotrophic cultivation of green microalgae Scenedesmus obliquus on cheese whey permeate for biodiesel production. Algal Res. 2014, 5, 241–248. [Google Scholar] [CrossRef]
- Begum, H.; Yusoff, F.M.; Banerjee, S.; Khatoon, H.; Shariff, M. Availability and utilization of pigments from microalgae. Crit. Rev. Food Sci. 2016, 56, 2209–2222. [Google Scholar] [CrossRef] [PubMed]
- Ordog, V.; Stirk, W.A.; Bálint, P.; Staden, J.; Lovász, C. Changes in lipid, protein and pigment concentrations in nitrogen-stressed Chlorella minutissima cultures. J. Appl. Phycol. 2012, 24, 907–914. [Google Scholar] [CrossRef]
- Møller, A.P.; Biard, C.; Blount, J.D.; Houston, D.C.; Ninni, P.; Saino, N.; Surai, P.F. Carotenoid-dependent signals: Indicators of foraging efficiency, immune competence or detoxification ability? Avian Poult. Biol. Rev. 2000, 11, 137–159. [Google Scholar] [CrossRef]
- Tevatia, R.; Allen, J.; Blum, P.; Demirel, Y.; Black, P. Modeling of rhythmic behavior in neutral lipid production due to continuous supply of limited nitrogen: Mutual growth and lipid accumulation in microalgae. Bioresour. Technol. 2014, 170, 152–159. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.K.; Niu, Y.F.; Ma, Y.H.; Xue, J.; Zhang, M.H.; Yang, W.D.; Liu, J.S.; Lu, S.H.; Guan, Y.; Li, H.Y. Molecular and cellular mechanisms of neutral lipid accumulation in diatom following nitrogen deprivation. Biotechnol. Biofuels 2013, 6, 67. [Google Scholar] [CrossRef] [Green Version]
- Andreotti, V.; Solimeno, A.; Rossi, S.; Ficara, E.; Marazzi, F.; Mezzanotte, V.; García, J. Bioremediation of aquaculture wastewater with the microalgae Tetraselmis suecica: Semi-continuous experiments, simulation and photo-respirometric tests. Sci. Total Environ. 2020, 738, 139859. [Google Scholar] [CrossRef] [PubMed]
- Paes, C.; Faria, G.R.; Tinoco, N.; Castro, D.; Lourenco, S.O. Growth, nutrient uptake and chemical composition of Chlorella sp. and Nannochloropsis oculata under nitrogen starvation. Lat. Am. J. Aquat. Res. 2017, 44, 275–292. [Google Scholar] [CrossRef]
- Tan, X.B.; Yang, L.B.; Zhang, W.W.; Zhang, X.C. Lipids production and nutrients recycling by microalgae mixotrophic culture in anaerobic digestate of sludge using wasted organics as carbon source. Bioresour. Technol. 2020, 297, 122379. [Google Scholar] [CrossRef]
- Li, T.T.; Zheng, Y.; Yu, L.; Chen, S.L. Mixotrophic cultivation of a Chlorella sorokiniana strain for enhanced biomass and lipid production. Biomass Bioenergy 2014, 66, 204–213. [Google Scholar] [CrossRef]
- Juntila, D.J.; Bautista, M.A.; Monotilla, W. Biomass and lipid production of a local isolate Chlorella sorokiniana under mixotrophic growth conditions. Bioresour. Technol. 2015, 191, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.F.; Liu, J.J.; Chu, F.F.; Lam, P.K.S.; Zeng, R.J. Enhancement of FAME productivity of Scenedesmus obliquus by combining nitrogen deprivation with sufficient phosphorus supply in heterotrophic cultivation. Appl. Energy 2015, 158, 348–354. [Google Scholar] [CrossRef]
- Shen, X.F.; Chu, F.F.; Lam, P.K.S.; Zheng, R.J. Biosynthesis of high yield fatty acids from Chlorella vulgaris NIES-227 under nitrogen starvation stress during heterotrophic cultivation. Water Res. 2015, 81, 294–300. [Google Scholar] [CrossRef]
- Knothe, G. Improving biodiesel fuel properties by modifying fatty ester composition. Energy Environ. Sci. 2009, 2, 759–766. [Google Scholar] [CrossRef]
- Hoekman, S.K.; Broch, A.; Robbins, C.; Ceniceros, E.; Natarajan, M. Review of biodiesel composition, properties and specifications. Renew. Sustain. Energy Rev. 2012, 16, 143–169. [Google Scholar] [CrossRef]
- Chen, X.; Yuan, Y.N.; Sun, P.; Mei, D.Q.; Cui, Y. Effects of structural features of the fatty acid methyl esters upon the cetane number of biodiesel. Chem. Eng. Oil Gas 2007, 36, 481–484. [Google Scholar] [CrossRef]
- Luo, W.; Yuan, Z.H.; Liao, C.P. Biodiesel standard and quality assessment. Renew. Energy Res. 2006, 4, 33–37. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, S.S.; Fan, J.F.; Fu, B.; Zhang, L.J. Methods to Improve Biomass and Lipid Accumulation of Lipid-Producing Microalgae by Using the Two-Stage Culture Strategy of Mixotrophic Culture and Nitrogen-Enriched & Nitrogen-Deficient Culture. A Patent for Invention. Chinese Patent No. CN102268377A, 7 December 2011. [Google Scholar]
- Shen, X.F. Heterotrophic and Mixotrophic Cultivation of Microalgae under Nitrogen Starvation for Biodiesel Production. Ph.D. Thesis, University of Science and Technology of China, Wuhan, China, 2017. [Google Scholar]
- Venkata Mohan, S.; Devi, M.P. Salinity stress induced lipid synthesis to harness biodiesel during dual mode cultivation of mixotrophic microalgae. Bioresour. Technol. 2014, 165, 288–294. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Conditions | Biomass Productivity (mg/(L·d)) |
|---|---|
| Autotrophic | 105.27 ± 2.33 c |
| Nitrogen deficient | 50.00 ± 10.33 d |
| Mixotrophic (+Glucose) | 215.71 ± 10.00 a |
| Nitrogen deficient & mixotrophic (+Glucose) | 164.29 ± 10.73 b |
| Culture Conditions Culture Conditions | Lipid Productivity (mg/(L∙d)) |
|---|---|
| Autotrophic | 35.61 ± 0.32 c |
| Nitrogen deficient | 22.48 ± 4.18 d |
| Mixotrophic (+Glucose) | 91.25 ± 1.86 b |
| Nitrogen deficient & mixotrophic (+Glucose) | 101.26 ± 4.18 a |
| Fatty Acids Composition | Autotrophic (% DW) | Nitrogen Deficient (% DW) | Mixotrophic (+Glucose) (% DW) | Nitrogen Deficient & Mixotrophic (+Glucose) (% DW) |
|---|---|---|---|---|
| C16:0 | 22 ± 0.26 d | 26.79 ± 0.55 b | 23.95 ± 0.09 c | 29.02 ± 0.21 a |
| C16:1 | 2.3 ± 0.05 c | 4.17 ± 0.02 a | 3.88 ± 0.06 b | 4.08 ± 0.06 a |
| C16:2 | 6.81 ± 0.06 a | 2.30 ± 0.03 b | — | 0.80 ± 0.02 c |
| C16:3 | — | 3.70 ± 0.04 a | — | 1.10 ± 0.03 b |
| C18:0 | 7.71 ± 0.05 c | 6.29 ± 0.08 d | 8.35 ± 0.03 b | 8.93 ± 0.07 a |
| C18:1 | 5.31 ± 0.14 d | 17.08 ± 0.20 b | 15.87 ± 0.04 c | 17.84 ± 0.04 a |
| C18:2 | 14.64 ± 0.11 d | 22.00 ± 0.10 a | 21.12 ± 0.02 b | 20.38 ± 0.20 c |
| C18:3 | 28.79 ± 0.25 a | 10.30 ± 0.12 d | 17.94 ± 0.12 b | 12.35 ± 0.05 c |
| C16 + C18 | 87.56 ± 0.5 d | 92.64 ± 0.12 b | 91.11 ± 0.09 c | 94.48 ± 0.32 a |
| Other | 12.44 ± 0.5 a | 7.36 ± 0.12 c | 8.89 ± 0.09 b | 5.52 ± 0.32 d |
| SFA | 33.93 ± 0.06 c | 35.71 ± 0.19 b | 35.46 ± 0.16 b | 40.15 ± 0.15 a |
| MUFA | 8.69 ± 0.06 c | 22.90 ± 0.20 a | 21.65 ± 0.09 b | 23.20 ± 0.18 a |
| PUFA | 57.38 ± 0.11 a | 41.39 ± 0.01 b | 42.89 ± 0.08 b | 36.65 ± 0.03 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Ji, L.; Feng, J.; Lv, J.; Xie, S. Effects of Combined Nitrogen Deficient and Mixotrophic (+Glucose) Culture on the Lipid Accumulation of Parachlorella Kessleri TY. Water 2021, 13, 3066. https://doi.org/10.3390/w13213066
Gao Y, Ji L, Feng J, Lv J, Xie S. Effects of Combined Nitrogen Deficient and Mixotrophic (+Glucose) Culture on the Lipid Accumulation of Parachlorella Kessleri TY. Water. 2021; 13(21):3066. https://doi.org/10.3390/w13213066
Chicago/Turabian StyleGao, Yifan, Li Ji, Jia Feng, Junping Lv, and Shulian Xie. 2021. "Effects of Combined Nitrogen Deficient and Mixotrophic (+Glucose) Culture on the Lipid Accumulation of Parachlorella Kessleri TY" Water 13, no. 21: 3066. https://doi.org/10.3390/w13213066
APA StyleGao, Y., Ji, L., Feng, J., Lv, J., & Xie, S. (2021). Effects of Combined Nitrogen Deficient and Mixotrophic (+Glucose) Culture on the Lipid Accumulation of Parachlorella Kessleri TY. Water, 13(21), 3066. https://doi.org/10.3390/w13213066