
Bacterial Microbiomes in the Sediments of Lotic Systems Ecologic Drivers and Role: A Case Study from the Mureş River, Transylvania, Romania

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Background
3. Materials and Methods
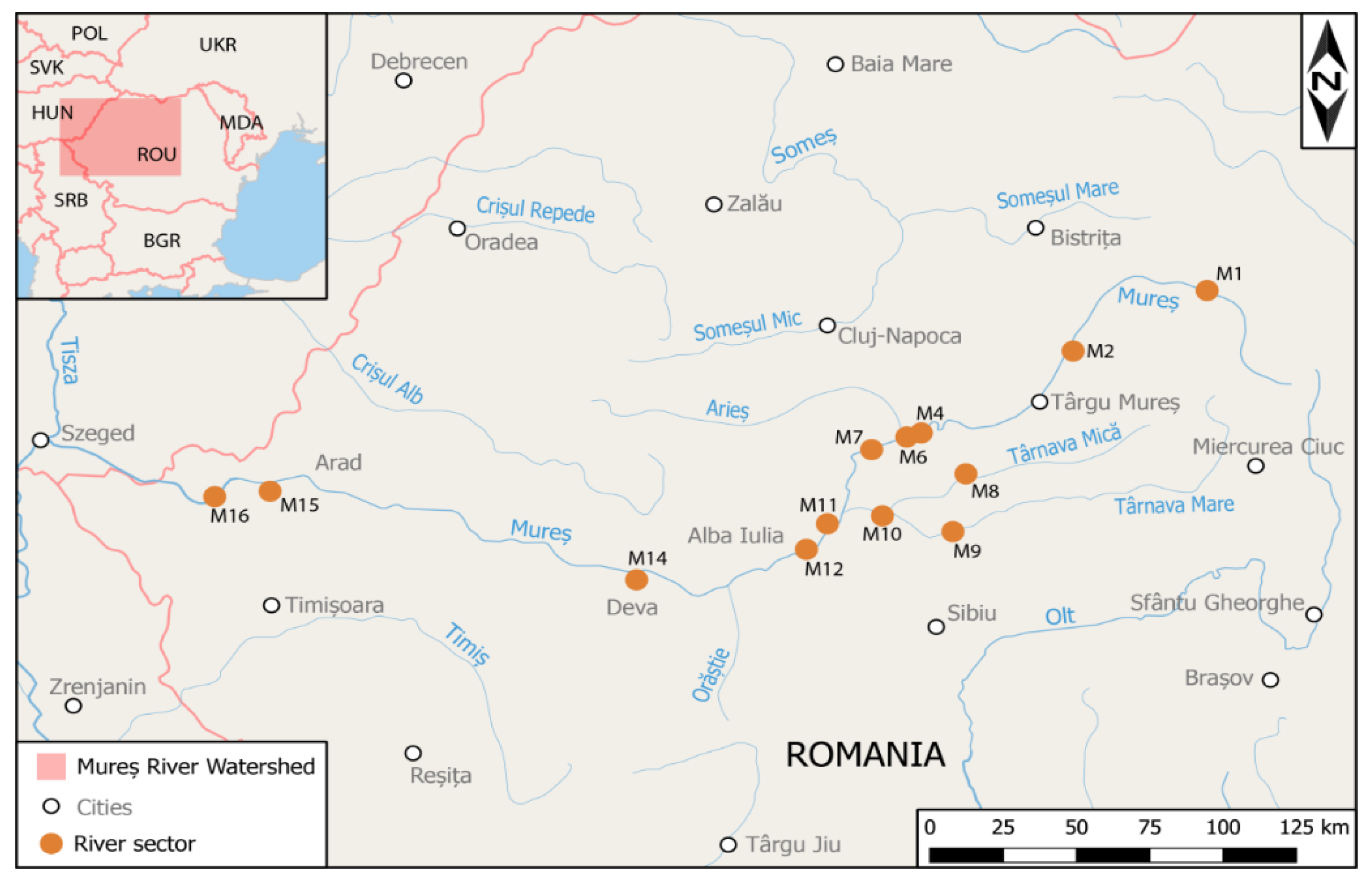
3.1. River Sector Sampling and Chemical Analysis
3.2. DNA Extraction and Quantification
3.3. 16S rRNA Gene Metabarcoding Analysis
3.4. Statistical Analysis
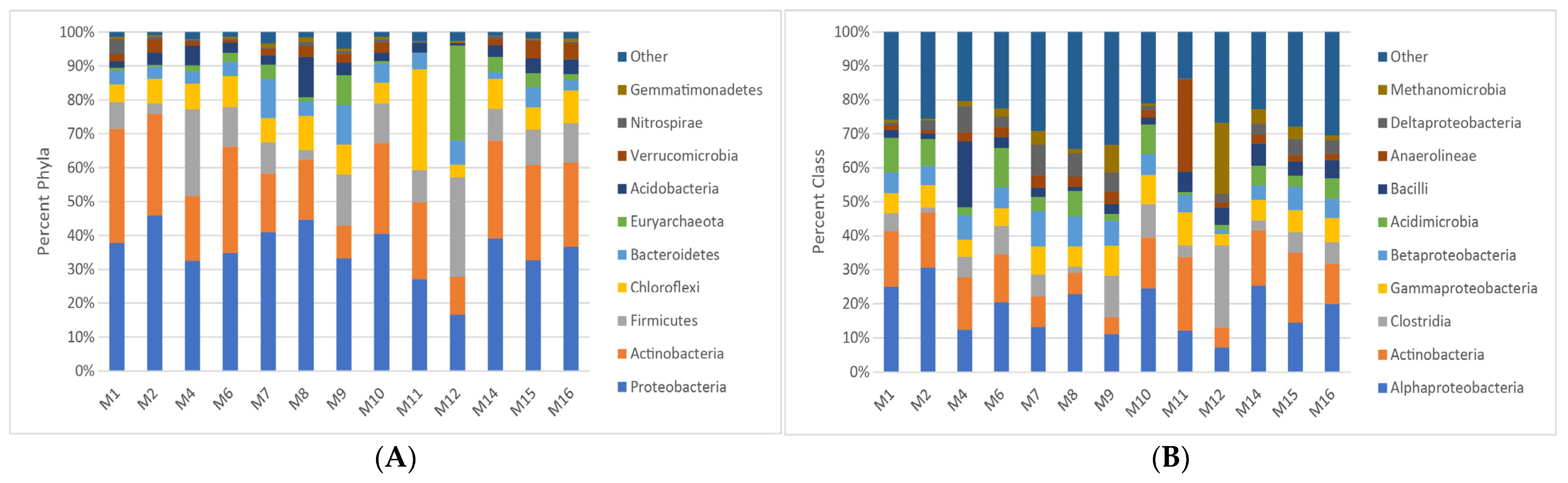
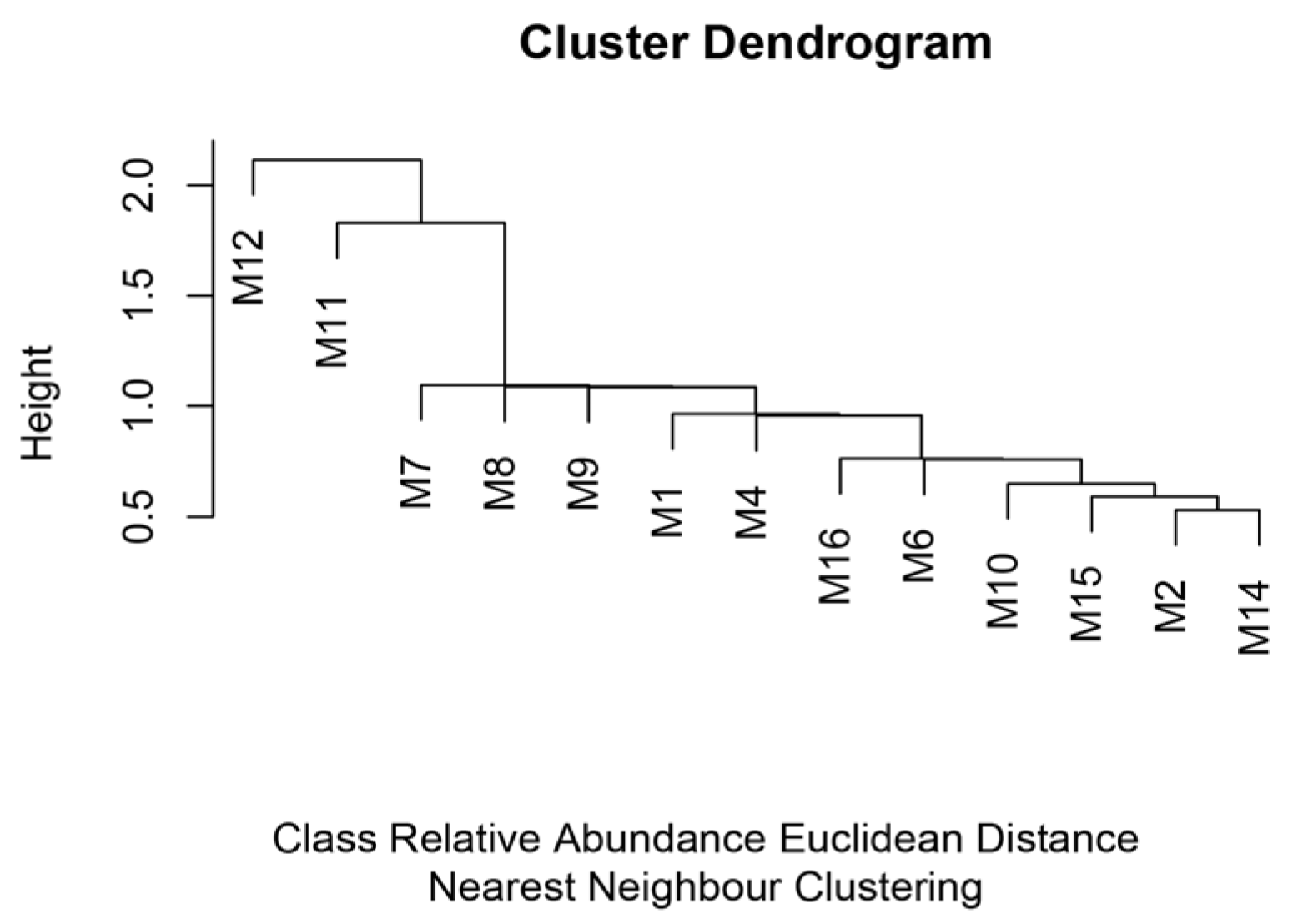
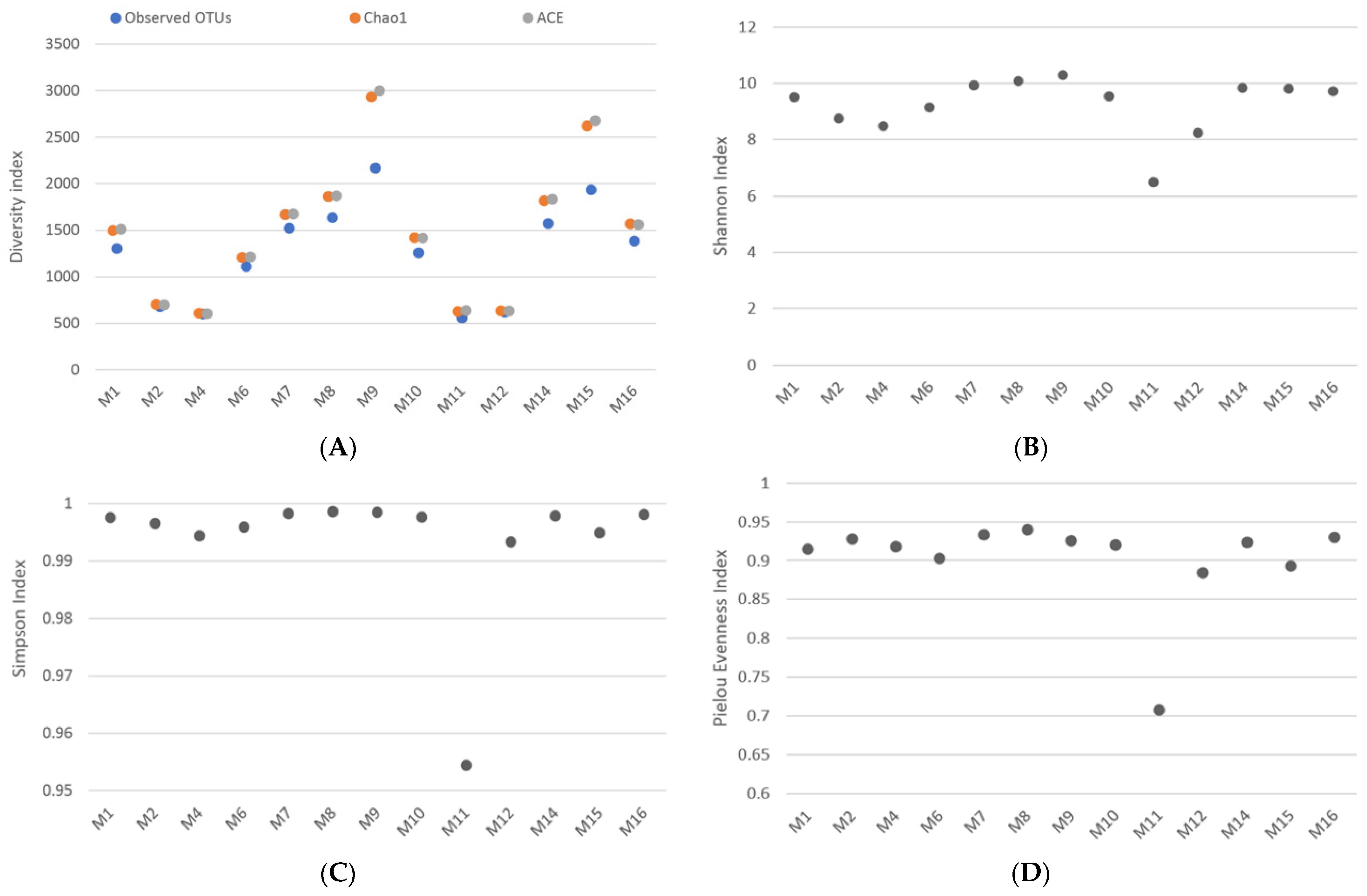
4. Results
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Global Water Partnership (GWP). Toward Water Security: A Framework for Action to Achieve the Vision for Water in the 21st Century; Global Water Partnership (GWP): Stockholm, Sweden, 2000. [Google Scholar]
- International Union for the Conservation of Nature and Natural Resources (IUCN). Vision for Water and Nature. A World Strategy for Conservation and Sustainable Management of Water Resources in the 21st Century—Compilation of All Project Documents; International Union for the Conservation of Nature and Natural Resources (IUCN): Cambridge, UK, 2000. [Google Scholar]
- De Villiers, M. Water: The Fate of Our Most Precious Resource; A Mariner Book; Houghton Mifflin Company: Boston, MA, USA, 2001; pp. 1–368. [Google Scholar]
- Fernández, N.; Gershenson, C. Measuring Complexity in an Aquatic Ecosystem. In Advances in Computational Biology; Springer: Cham, Switzerland, 2013. [Google Scholar]
- Gough, P. From Sea to Source, International Guidance for the Restoration of Fish Migration Highways; Regional Water Authority Hunze en Aa’s: Veendam, The Netherlands, 2012; Chapter 1; pp. 21–27. [Google Scholar]
- Middleton, N. Rivers: A Very Short Introduction; Oxford University Press: Oxford, UK, 2012; pp. 1–127. [Google Scholar]
- Shiklomanov, I.A. Forthcoming World Water Resources at the Beginning of the 21st Century; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Oprean, L. Apa Resursă Fundamentală a Dezvoltării Durabile. Metode şi Tehnici Neconvenţionale de Epurare şi Tratare a Apei; Coordinator Oprean, L.; Editura Academiei Române: București, Romania, 2012; Volume 1, Available online: https://appliedecologysibiu.files.wordpress.com/2016/02/lp-horea-olosutean-ro.pdf (accessed on 2 March 2021).
- Oprean, L. Apa Resursă Fundamentală a Dezvoltării Durabile. Metode şi Tehnici Neconvenţionale de Epurare şi Tratare a Apei; Coordinator Oprean, L.; Editura Academiei Române: București, Romania, 2012; Volume 2, Available online: https://doctorat.unibuc.ro/wp-content/uploads/sites/13/2019/01/l.-CV-SARBU.pdf (accessed on 2 March 2021).
- Brookes, A.; Gregory, K.; Dawson, F. An assessment of river channelization in England and Wales. Sci. Total Environ. 1983, 27, 97–111. [Google Scholar] [CrossRef]
- Kirchman, D.L. The uptake of inorganic nutrients by heterotrophic bacteria. Microb. Ecol. 1994, 28, 255–271. [Google Scholar] [CrossRef] [PubMed]
- Gregory, K. The human role in changing river channels. Geomorphology 2006, 79, 172–191. [Google Scholar] [CrossRef]
- Bai, Y.; Qi, W.; Liang, J.; Qu, J. Using high-throughput sequencing to assess the impacts of treated and untreated wastewater discharge on prokaryotic communities in an urban river. Appl. Microbiol. Biotechnol. 2013, 98, 1841–1851. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-González, C.; Niño-García, J.P.; Del Giorgio, P.A. Terrestrial origin of bacterial communities in complex boreal freshwater networks. Ecol. Lett. 2015, 18, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Curtean-Bănăduc, A.; Olosutean, H.; Bănăduc, D. Influence of Environmental Variables on the Structure and Diversity of Ephemeropteran Communities: A Case Study of the Timiş River, Romania. Acta Zool. Bulg. 2016, 68, 215–224. [Google Scholar]
- Curtean-Bănăduc, A.; Maric, S.; Gabor, G.; Didenko, A.; Rey Planellas, S.; Bănăduc, D. Hucho hucho (Linnaeus, 1758): Last natural viable population in the Eastern Carpathians—Conservation elements. Turk. J. Zool. 2019, 43, 215–223. [Google Scholar] [CrossRef]
- Curtean-Bănăduc, A.; Didenko, A.; Guti, G.; Bănăduc, D. Telestes souffia (Risso, 1827) species conservation at the Eastern limit of range. Appl. Ecol. Environ. Res. 2018, 16, 291–303. [Google Scholar] [CrossRef]
- IUCN Environmental Flows, Managing Water Allocation and Trade-Offs. Available online: https://www.iucn.org/downloads/water_briefing_eflows.pdf (accessed on 2 March 2021).
- Kuriqi, A.; Pinheiro, A.N.; Sordo-Ward, A.; Garrote, L. Water-energy-ecosystem nexus: Balancing competing interests at a run-of-river hydropower plant coupling a hydrologic–ecohydraulic approach. Energy Convers. Manag. 2020, 223, 113267. [Google Scholar] [CrossRef]
- Kuriqi, A.; Pinheiro, A.N.; Sordo-Ward, A.; Bejarano, M.D.; Garrote, L. Ecological impacts of run-of-river hydropower plants—Current status and future prospects on the brink of energy transition. Renew. Sustain. Energy Rev. 2021, 142, 110833. [Google Scholar] [CrossRef]
- Costea, G.; Push, M.T.; Bănăduc, D.; Cosmoiu, D.; Curtean-Bănăduc, A. A Review of Hydropower Plants in Romania: Distribution, Current Knowledge, and Their Effects on Fish in Headwater Streams. Renew. Sustain. Energy Rev. 2021, 54, 111003. [Google Scholar] [CrossRef]
- Bănăduc, D.; Joy, M.; Olosutean, H.; Afanasyev, S.; Curtean-Bănăduc, A. Natural and anthropogenic driving forces as key elements in the Lower Danube Basin—South-Eastern Carpathians—North-Western Black Sea coast area lakes, a broken stepping stones for fish in a climatic change scenario. Environ. Sci. Eur. 2020, 32, 73. [Google Scholar] [CrossRef]
- Curtean-Bănăduc, A.; Burcea, A.; Mihuţ, C.-M.; Bănăduc, D. The benthic trophic corner stone compartment in POPs transfer from abiotic environment to higher trophic levels—Trichoptera and Ephemeroptera pre-alert indicator role. Water 2021, 13, 1778. [Google Scholar] [CrossRef]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Champomier Vergès, M.-C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Herrero Corral, G.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Meybeck, M. Global analysis of river systems: From Earth system controls to Anthropocene syndromes. Philos. Trans. R. Soc. B Biol. Sci. 2003, 358, 1935–1955. [Google Scholar] [CrossRef] [PubMed]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Leveque, C. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2005, 81, 163–182. [Google Scholar] [CrossRef]
- Dudgeon, D. Anthropocene Extinctions: Global Threats to Riverine Biodiversity and the Tragedy of the Freshwater Commons. In River Conservation: Challenges and Opportunities; Sabater, S., Elosegi, A., Eds.; Fundación BBVA: Bilbao, Spain, 2013; pp. 129–167. [Google Scholar]
- Steffen, W.; Crutzen, P.J.; McNeill, J.R. The Anthropocene: Are Humans Now Overwhelming the Great Forces of Nature? University of California Press: Los Angeles, CA, USA, 2007; Volume 36, pp. 614–621. [Google Scholar]
- Stevenson, R.J.; Sabater, S. Understanding global change in river ecosystems: Science to support policy in a changing world. Hydrobiologia 2010, 657, 3–18. [Google Scholar] [CrossRef]
- Schneider-Binder, E. The Habitats along the Upper Danube in Germany and Changes to Them Induced by Human Impacts. In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 27–48. [Google Scholar]
- Cianfaglione, K.; Pedrotti, F. Italy in the Danube Geography: Territory, Landscape, Environment, Vegetation, Fauna, Culture, Human Management and Outlooks for the Future. In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 87–118. [Google Scholar]
- Adámek, Z.; Jurajdová, Z.; Janáč, M.; Zahrádková, S.; Němejcová, D.; Jurajda, P. The Response of Fish Assemblages to Human Impacts along the Lower Stretch of the Rivers Morava and Dyje (Danube River Basin, Czech Republic). In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 135–150. [Google Scholar]
- Ćaleta, M.; Mustafić, P.; Zanella, D.; Buj, I.; Marčić, Z.; Mrakovčić, M. Human Impact on the Dobra River (Croatia). In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 151–168. [Google Scholar]
- Dekić, R.; Ivanc, A.; Ćetković, D.; Lolić, A. Anthropogenic Impact and Environmental Quality of Different Tributaries of the River Vrbas (Bosnia and Hertzegovina). In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 169–214. [Google Scholar]
- Guti, G. Assessment of Long-Term Changes in the Szigetköz Floodplain of the Danube River. In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 215–240. [Google Scholar]
- Ðikanović, V.; Nikčević, M.; Mićković, B.; Hegediš, A.; Mrdak, D.; Pešić, V. Anthropogenic Pressures on Watercourses of the Danube River Basin in Montenegro. In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 241–256. [Google Scholar]
- Lenhardt, M.; Snederevac-Lalić, M.; Hegediš, A.; Skorić, S.; Cvijanović, G.; Višnjić-Jeftić, Ž.; Djikanović, V.; Jovičić, K.; Jaćimović, M.; Jarić, I. Human Impacts on Fish Fauna in the Danube River in Serbia: Current Status and Ecological Implications. In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 257–280. [Google Scholar]
- Mišiková Elexová, E.; Makovinská, J. Assessment of the Aquatic Ecosystem in the Slovak Stretch of the Danube River. In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 281–300. [Google Scholar]
- Maślanko, W.; Ferencz, B.; Dawidek, J. State and Changes of Natural Environment in Polish Part of the Danube River Basin Poland. In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 301–326. [Google Scholar]
- Afanasyev, S.; Lyashenko, A.; Iarochevitch, A.; Lietytska, O.; Zorina-Sakharova, K.; Marushevska, O. Pressures and Impacts on Ecological Status of Surface Water Bodies in Ukrainian part of the Danube River Basin. In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 327–359. [Google Scholar]
- Bakiu, R. Drina River (Sava’s Tributary of Danube River) and Human Impact in Albania. In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 359–380. [Google Scholar]
- Kostov, V.; Slavevska-Stamenkovic, V.; Ristovska, M.; Stojov, V.; Marić, S. Characteristics of the Danube Drainage Area in the Republic of Macedonia. In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 381–392. [Google Scholar]
- Kenderov, L.; Trichkova, T. Long-Term Changes in the Ecological Conditions of the Iskar River (Danube River Basin, Bulgaria). In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 393–424. [Google Scholar]
- Curtean-Bănăduc, A.; Bănăduc, D. Human impact effects on Târnava Mare Basin Aquatic Biodiversity (Transylvania, Romania). In Human Impact on Danube Watershed Biodiversity in the XXI Century. Geobotany Studies (Basics, Methods and Case Studies); Bănăduc, D., Curtean-Bănăduc, A., Pedrotti, F., Cianfaglione, K., Akeroyd, J., Eds.; Springer: Cham, Switzerland, 2020; pp. 425–437. [Google Scholar]
- Posea, G. Enciclopedia Geografică a României; Ştiinţifică şi Enciclopedică: Bucureşti, Romania, 1982; p. 262. [Google Scholar]
- Mutihac, V. Structura Geologică a Teritoriului României; Editura Tehnicǎ: Bucharest, Romania, 1990. [Google Scholar]
- Diaconu, C.; Stănculescu, S. Rîurile României, Monografie Hidrologică; Institutul de Meteorologie și Hidrologie: București, Romania, 1971; pp. 1–750. [Google Scholar]
- Badea, L.; Bugă, D.; Cioflica, G.; Cucu, V.; Doniţă, I.; Gâştescu, P.; Iordan, I.; Morariu, T.; Niculescu, G.; Oancea, D.; et al. Geografia României, II, Geografia Umană şi Economic; Editura Academiei Republicii Socialiste România: Bucharest, Romania, 1984; pp. 1–543. [Google Scholar]
- Triebskorn, R.; Telcean, I.; Casper, H.; Farkas, A.; Sandu, C.; Stan, G.; Colărescu, O.; Dori, T.; Köhler, H.-R. Monitoring pollution in River Mureş, Romania, part II: Metal accumulation and histopathology in fish. Environ. Monit. Assess. 2007, 141, 177–188. [Google Scholar] [CrossRef]
- Curtean-Bănăduc, A.; Bănăduc, D. The Transylvanian Water Tower through history. Danub. News 2015, 17, 1–4. [Google Scholar]
- Curtean-Bănăduc, A.; Bănăduc, D.; Burcea, A.; Berg, V.; Lyche, J.L.; Gheorghe, L.M. Persistent organic pollutants in Mureş watershed. In The Impact of Persistent Organic Pollutants on Freshwater Ecosystems and Human Health; Curtean-Bănăduc, A., Ed.; Publisher Lucian Blaga, University of Sibiu: Sibiu, Romania, 2016; pp. 117–152. [Google Scholar]
- Burcea, A.; Boeraş, I.; Mihuţ, C.-M.; Bănăduc, D.; Matei, C.; Curtean-Bănăduc, A. Adding the Mureş River Basin (Transylvania, Romania) to the List of Hotspots with High Contamination with Pharmaceuticals. Sustainability 2020, 12, 10197. [Google Scholar] [CrossRef]
- Curtean-Bănăduc, A.; Burcea, A.; Mihuţ, C.-M.; Berg, V.; Lyche, J.L.; Bănăduc, D. Bioaccumulation of persistent organic pollutants in the gonads of Barbus barbus (Linnaeus, 1758). Ecotoxicol. Environ. Saf. 2020, 201, 110852. [Google Scholar] [CrossRef]
- Bănăduc, D.; Sas, A.; Cianfaglione, K.; Barinova, S.; Curtean-Bănăduc, A. The Role of Aquatic Refuge Habitats for Fish, and Threats in the Context of Climate Change and Human Impact, during Seasonal Hydrological Drought in the Saxon Villages Area (Transylvania, Romania). Atmosphere 2021, 12, 1209. [Google Scholar] [CrossRef]
- Curtean–Bănăduc, A.; Bănăduc, D.; Bucşa, C. Watersheds Management (Transylvania/Romania): Implications, Risks, Solutions. In Strategies to Enhance Environmental Security in Transition Countries; NATO Science for Peace and Security Series C-Environmental Security; Springer: Dordrecht, The Netherlands, 2007; pp. 225–238. ISBN 978-1-4020-5994-0. ISSN 1971-4668. [Google Scholar] [CrossRef]
- Bănăduc, D.; Curtean-Bănăduc, A.; Cianfaglione, K.; Akeroyd, J.R.; Cioca, L.-I. Proposed environmental risk management elements in a Carpathian valley basin, within the Roşia Montană European historical mining area. Int. J. Environ. Res. Public Health 2021, 18, 4565. [Google Scholar] [CrossRef] [PubMed]
- Mureş Water Directorate (MWD): Report for 2004—Annual Synthesis Concerning the Water Quality Protection in Mureş Hydrographic Basin, Vol. I and II. (1) (PDF) The Mureş River Ecosystem—Scientific Background Information as the Basis for a Catchment Approach in the Framework of IAD. Available online: https://www.researchgate.net/publication/242267567_The_Mures_River_ecosystem_-_scientific_background_information_as_the_basis_for_a_catchment_approach_in_the_framework_of_IAD (accessed on 15 August 2021).
- Köhler, H.-R.; Triebskorn, R.; Sandu, C. Monitoring pollution in the river Mureş. Bull. Int. Assoc. Danub. Res.-Danub. News 2005, 11, 1–6. [Google Scholar]
- Köhler, H.-R.; Sandu, C.; Scheil, V.; Nagy-Petrică, E.M.; Segner, H.; Telcean, I.; Stan, G.; Triebskorn, R. Monitoring Pollution in River MureŞ, Romania, Part III: Biochemical effect markers in fish and integrative reflection. Environ. Monit. Assess. 2006, 127, 47–54. [Google Scholar] [CrossRef]
- Sandu, C.; Farkas, A.; Musa-Iacob, R.; Ionică, D.; Parpală, L.; Zinevici, V.; Dobre, D.; Radu, M.; Presing, M.; Casper, H.; et al. Monitoring Pollution in River Mureş, Romania, Part I: How Aquatic Communities Are Affected; Environmental Monitoring and Assessment; Springer: Bucureşti, Romania, 2006. [Google Scholar]
- Bănăduc, D.; Oprean, L.; Bogdan, A. Fish species of community interest management issues in Natura 2000 site Sighişoara-Târnava Mare (Transylvania, Romania). Rev. Econ. 2011, 3, 23–27. [Google Scholar]
- Curtean-Bănăduc, A.; Bănăduc, D. Benthic macro-invertebrate and fish communities of some southern Tarnava Mare River tributaries (Transylvania, Romania). Transylv. Rev. Syst. Ecol. Res. 2007, 4, 135–148. [Google Scholar]
- Bănăduc, D. Fish associations—Habitats quality relation in the Târnave rivers ecological assessment. Transylv. Rev. Syst. Ecol. Res. 2005, 2, 123–136. [Google Scholar]
- Bănăduc, D.; Pânzar, C.; Bogorin, P.; Hoza, O.; Curtean-Bănăduc, A. Human impact on Tȃrnava Mare River and its effects on aquatic biodiversity. Acta Oecol. Carp. 2016, 9, 189–198. [Google Scholar]
- Savio, D.; Sinclair, L.; Ijaz, U.Z.; Parajka, J.; Reischer, G.; Stadler, P.; Blaschke, A.P.; Blöschl, G.; Mach, R.; Kirschner, A.; et al. Bacterial diversity along a 2600 km river continuum. Environ. Microbiol. 2015, 17, 4994–5007. [Google Scholar] [CrossRef]
- Lupan, I.; Carpa, R.; Oltean, A.; Kelemen, B.; Popescu, O. Release of Antibiotic Resistant Bacteria by a Waste Treatment Plant from Romania. Microbes Environ. 2017, 32, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Florea, A.B. Antimicrobial susceptibility of Escherichia coli isolated from Arieş river (Romania). An. Univ. Oradea Fasc. Biol. 2011, 18, 34–38. [Google Scholar]
- Ward, J.V. The four-dimensional nature of lotic ecosystems. J. N. Am. Benthol Soc. 1989, 8, 2–8. [Google Scholar] [CrossRef]
- Del Monte-Luna, P.; Brook, B.W.; Zetina-Rejón, M.J.; Cruz-Escalona, V.H. The carrying capacity of ecosystems. Glob. Ecol. Biogeogr. 2004, 13, 485–495. [Google Scholar] [CrossRef]
- Cherry, J.A. Ecology of wetland ecosystems: Water, substrate, and life. Nat. Educ. Knowl. 2011, 3, 16. [Google Scholar]
- Afanasyev, S.A. Forming of Hydrobiota of the River Systems in the Territory of Ukraine in View of History of Hydrographic Net. Hydrobiol. J. 2015, 51, 3–12. [Google Scholar] [CrossRef]
- Fogarty, M.J.; Gamble, R.; Perretti, C.T. Dynamic Complexity in Exploited Marine Ecosystems. Front. Ecol. Evol. 2016, 4, 68. [Google Scholar] [CrossRef] [Green Version]
- Barinova, S. Empirical Model of the Functioning of Aquatic Ecosystems. Int. J. Oceanogr. Aquac. 2017, 1, 1–9. [Google Scholar] [CrossRef]
- Cianfaglione, K. Plant Landscape and Models of French Atlantic Estuarine Systems. Extended Summary of the Doctoral Thesis. Transylv. Rev. Syst. Ecol. Res. 2021, 23, 15–36. [Google Scholar]
- Zimmerman, B.; Lindberg, C.; Plsek, P. A Complexity Science Primer. In Edgeware: Insights from Complexity Science for Health Care Leaders, 2nd ed.; Zimmerman, B., Lindberg, C., Plsek, P., Eds.; VHA Inc.: Irving, TX, USA, 2001; pp. 3–20. [Google Scholar]
- Potter, P.E.; Hamblin, W.K. Big Rivers Worldwide; Brigham Young University Geology Studies: Provo, UT, USA, 2005. [Google Scholar]
- Best, J. Anthropogenic stresses on the world’s big rivers. Nat. Geosci. 2019, 12, 7–21. [Google Scholar] [CrossRef]
- Bănăduc, D.; Rey, S.; Trichkova, T.; Lenhardt, M.; Curtean-Bănăduc, A. The Lower Danube River—Danube Delta—North West Black Sea: A pivotal area of major interest for the past, present and future of its fish fauna—A short review. Sci. Total Environ. 2016, 545, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Tockner, K.; Zarfl, C.; Robinson, C. Rivers of Europe, 2nd ed.; Elsevier: Bayern, Germany, 2021; p. 800. [Google Scholar]
- Dahm, C.; Bulton, A.; Correa, L.; Kingsford, R.; Jenkins, K.; Sheldon, F. The role of science in planning, policy and conservation of river ecosystems. In Rivers Conservation: Challenges and Opportunities; Sabater, S., Elosegi, A., Eds.; Fundación BBVA: Bilbao, Spain, 2013; p. 399. [Google Scholar]
- Amyes, S. What’s the Secret of Bacteria’s Success? Bacteria: A Very Short Introduction; Oxford University Press’s Academic Insights for the Thinking World: Oxford, UK, 2013. [Google Scholar]
- Maddock, I. The importance of physical habitat assessment for evaluating river health. Freshw. Biol. 1999, 41, 373–391. [Google Scholar] [CrossRef]
- Christoph, H.; Patrik, L.; Günther, U.; Ulrich, P.; Habersack, H. The Role of Sediment and Sediment Dynamics in the Aquatic Environment. In Riverine Ecosystems Management; Springer: Cham, Switzerland, 2018; pp. 151–169. [Google Scholar]
- Bănăduc, D.; Voicu, R.; Curtean-Bănăduc, A. Sediments as factor in the fate of the threatened endemic fish species Romanichthys valsanicola Dumitrescu, Bănărescu and Stoica, 1957 (Vâlsan River basin, Danube Basin). Transylv. Rev. Syst. Ecol. Res. 2020, 22, 15–30. [Google Scholar] [CrossRef]
- Battin, T.J.; Luyssaert, S.; Kaplan, L.A.; Aufdenkampe, A.K.; Richter, A.; Tranvik, L.J. The boundless carbon cycle. Nat. Geosci. 2009, 2, 598–600. [Google Scholar] [CrossRef]
- Findlay, S. Stream microbial ecology. J. N. Am. Benthol. Soc. 2010, 29, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Madsen, E.L. Microorganisms and their roles in fundamental biogeochemical cycles. Curr. Opin. Biotechnol. 2011, 22, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Vaqué, D.; Pace, M.L.; Findlay, S.; Lints, D. Fate of bacterial production in a heterotrophic ecosystem: Grazing by protists and metazoans in the Hudson Estuary. Mar. Ecol. Prog. Ser. 1992, 89, 155–163. [Google Scholar] [CrossRef]
- Naegeli, M.W.; Uehlinger, U. Contribution of the Hyporheic Zone to Ecosystem Metabolism in a Prealpine Gravel-Bed-River. J. N. Am. Benthol. Soc. 1997, 16, 794–804. [Google Scholar] [CrossRef]
- Gibert, J.; Mathieu, J.; Fournier, F. Groundwater/Surface Water Ecotones: Biological and Hydrological Interactions and Management Options, 1st ed.; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Pusch, M.; Fiebig, D.; Brettar, I.; Eisenmann, H.; Ellis, B.K.; Kaplan, L.A.; Lock, M.A.; Naegeli, M.W.; Traunspurger, W. The role of micro-organisms in the ecological connectivity of running waters. Freshw. Biol. 1998, 40, 453–495. [Google Scholar] [CrossRef]
- Craft, J.A.; Stanford, J.A.; Pusch, M. Microbial respiration within a floodplain aquifer of a large gravel-bed river. Freshw. Biol. 2002, 47, 251–261. [Google Scholar] [CrossRef]
- Huber, D.H.; Ugwuanyi, I.R.; Malkaram, S.A.; Montenegro-Garcia, N.A.; Noundou, V.L.; Chavarria-Palma, J.E. Metagenome Sequences of Sediment from a Recovering Industrialized Appalachian River in West Virginia. Genome Announc. 2018, 6, e00350-18. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, S.M.; Jones, E.; Bearquiver, A.; Blackwolf, F.; Roundstone, W.; Scott, N.; Hooker, J.; Madsen, R.; Coleman, M.; Gilbert, J.A. Human and Environmental Impacts on River Sediment Microbial Communities. PLoS ONE 2014, 9, e97435. [Google Scholar]
- Wang, Y.; Sheng, H.-F.; He, Y.; Wu, J.-Y.; Jiang, Y.-X.; Tam, N.F.-Y.; Zhou, H.-W. Comparison of the Levels of Bacterial Diversity in Freshwater, Intertidal Wetland, and Marine Sediments by Using Millions of Illumina Tags. Appl. Environ. Microbiol. 2012, 78, 8264–8271. [Google Scholar] [CrossRef] [Green Version]
- Thompson, L.R.; Sanders, J.G.; McDonald, D.; Amir, A.; Ladau, J.; Locey, K.J.; Prill, R.J.; Tripathi, A.; Gibbons, S.M.; Ackermann, G.; et al. A communal catalogue reveals Earth’s multiscale microbial diversity. Nature 2017, 551, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Engel, A.S.; Meisinger, D.B.; Porter, M.L.; Payn, R.A.; Schmid, M.; Stern, L.A.; Schleifer, K.H.; Lee, N.M. Linking phylogenetic and functional diversity to nutrient spiraling in microbial mats from Lower Kane Cave (USA). ISME J. 2010, 4, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Kristiansson, E.; Fick, J.; Janzon, A.; Grabic, R.; Rutgersson, C.; Weijdegård, B.; Söderström, H.; Larsson, D.J. Pyrosequencing of antibiotic-contaminated river sediments reveals high levels of resistance and gene transfer elements. PLoS ONE 2011, 6, e17038. [Google Scholar] [CrossRef]
- Zeglin, L.H.; Dahm, C.N.; Barrett, J.E.; Gooseff, M.; Fitpatrick, S.K.; Takacs-Vesbach, C.D. Bacterial Community Structure along Moisture Gradients in the Parafluvial Sediments of Two Ephemeral Desert Streams. Microb. Ecol. 2010, 61, 543–556. [Google Scholar] [CrossRef]
- Ramsey, P.W.; Rillig, M.C.; Feris, K.P.; Gordon, N.S.; Moore, J.N.; Holben, W.E.; Gannon, J.E. Relationship between communities and processes; new insights from a field study of a contaminated ecosystem. Ecol. Lett. 2005, 8, 1201–1210. [Google Scholar] [CrossRef]
- Feris, K.P.; Ramsey, P.W.; Frazar, C.; Rillig, M.C.; Gannon, J.E.; Holben, W.E. Structure and seasonal dynamics of hyporheic zone microbial communities in free-stone rivers of the estern United States. Microb. Ecol. 2003, 46, 200–215. [Google Scholar] [CrossRef]
- Feris, K.P.; Ramsey, P.W.; Frazar, C.; Rillig, M.; Moore, J.N.; Gannon, J.E.; Holben, W.E. Seasonal dynamics of shallow-hyporheic-zone microbial community structure along a heavy-metal contamination gradient. Appl. Environ. Microbiol. 2004, 70, 2323–2331. [Google Scholar] [CrossRef] [Green Version]
- Feris, K.P.; Ramsey, P.W.; Gibbons, S.M.; Frazar, C.; Rillig, M.C.; Moore, J.N.; Gannon, J.E.; Holben, W.E. Hyporheic microbial community development is a sensitive indicator of metal contamination. Environ. Sci. Technol. 2009, 43, 6158–6163. [Google Scholar] [CrossRef]
- Crump, B.C.; Armbrust, E.V.; Baross, J.A. Phylogenetic Analysis of Particle-Attached and Free-Living Bacterial Communities in the Columbia River, Its Estuary, and the Adjacent Coastal Ocean. Appl. Environ. Microbiol. 1999, 65, 3192–3204. [Google Scholar] [CrossRef] [Green Version]
- Zwart, G.; Crump, B.C.; van Kamst-Agterveld, M.P.; Hagen, F.; Han, S.K. Typical freshwater bacteria: An analysis of available 16S rRNA gene sequences from plankton of lakes and rivers. Aquat. Microb. Ecol. 2002, 28, 141–155. [Google Scholar] [CrossRef] [Green Version]
- Cottrell, M.T.; Waidner, L.A.; Yu, L.; Kirchman, D.L. Bacterial diversity of metagenomic and PCR libraries from the Delaware River. Environ. Microbiol. 2005, 7, 1883–1895. [Google Scholar] [CrossRef] [PubMed]
- Winter, C.; Hein, T.; Kavka, G.; Mach, R.L.; Farnleitner, A.H. Longitudinal Changes in the Bacterial Community Composition of the Danube River: A Whole-River Approach. Appl. Environ. Microbiol. 2007, 73, 421–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemke, M.J.; Lienau, E.K.; Rothe, J.; Pagioro, T.A.; Rosenfeld, J.; DeSalle, R. Description of Freshwater Bacterial Assemblages from the Upper Paraná River Floodpulse System, Brazil. Microb. Ecol. 2008, 57, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Mueller-Spitz, S.R.; Goetz, G.W.; McLellan, S.L. Temporal and spatial variability in nearshore bacterioplankton communities of Lake Michigan. FEMS Microbiol. Ecol. 2009, 67, 511–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S. A Guide to the Natural History of Freshwater Lake Bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Huang, S.; Sun, G.; Xu, Z.; Xu, M. Phylogenetic diversity, composition and distribution of bacterioplankton community in the Dongjiang River, China. FEMS Microbiol. Ecol. 2012, 80, 30–44. [Google Scholar] [CrossRef] [Green Version]
- Lv, X.; Yu, J.; Fu, Y.; Ma, B.; Qu, F.; Ning, K.; Wu, H. A meta-analysis of the bacterial and archaeal diversity observed in wetland soils. Sci. World J. 2014, 2014, 437684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böcklemann, U.; Manz, W.; Neu, T.R.; Szewzyk, U. Characterization of the microbial community of lotic organic aggregates (‘river snow’) in the Elbe River of Germany by cultivation and molecular methods. FEMS Microbiol. Ecol. 2000, 33, 157–170. [Google Scholar] [CrossRef]
- Liu, M.; Cheng, S.; Ou, D.; Yang, Y.; Liu, H.; Hou, L.; Gao, L.; Xu, S. Organochlorine pesticides in surface sediments and suspended particulate matters from the Yangtze estuary, China. Environ. Pollut. 2008, 156, 168–173. [Google Scholar] [CrossRef]
- Crump, B.C.; Hobbie, J.E. Synchrony and seasonality in bacterioplankton communities of two temperate rivers. Limnol. Oceanogr. 2005, 50, 1718–1729. [Google Scholar] [CrossRef]
- Köchling, T.; Sanz, J.L.; Galdino, L.; Florencio, L.; Kato, M.T. Impact of pollution on the microbial diversity of a tropical river in an urbanized region of northeastern Brazil. Int. Microbiol. 2017, 20, 11–24. [Google Scholar] [PubMed]
- Zhang, C.; Liu, L.; Ma, Y.; Li, F. Using isomeric and metabolic ratios of DDT to identify the sources and fate of DDT in Chinese agricultural topsoil. Environ. Sci. Technol. 2018, 52, 1990–1996. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and Qualitative β Diversity Measures Lead to Different Insights into Factors That Structure Microbial Communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abia, A.L.K.; Alisoltani, A.; Keshri, J.; Ubomba-Jaswa, E. Metagenomic analysis of the bacterial communities and their functional profiles in water and sediments of the Apies River, South Africa, as a function of land use. Sci. Total Environ. 2018, 616, 326–334. [Google Scholar] [CrossRef]
- Su, Z.; Dai, T.; Tang, Y.; Tao, Y.; Huang, B.; Mu, Q.; Wen, D. Sediment bacterial community structures and their predicted functions implied the impacts from natural processes and anthropogenic activities in coastal area. Mar. Pollut. Bull. 2018, 131, 481–495. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, J.; Li, H.; Yang, H.; Peng, C.; Peng, Z.; Lu, L. Shift in the microbial community composition of surface water and sediment along an urban river. Sci. Total Environ. 2018, 627, 600–612. [Google Scholar] [CrossRef] [PubMed]
- Thorp, J.H.; Thoms, M.C.; Delong, M.D. The riverine ecosystem synthesis: Biocomplexity in river networks across space and time. River Res. Appl. 2006, 22, 123–147. [Google Scholar] [CrossRef]
- Sridhar, K.R. Structural and Functional Attributes to Evaluate Lotic Ecosystems Health, in Recent Research Trends in Environmental Management; Prasad, A.G.D., Ed.; Daya Publishing House: Delhi, India, 2016; Chapter 2; pp. 8–24. [Google Scholar]
- Terui, A.; Kim, S.; Dolph, C.L.; Kadoya, T.; Miyazaki, Y. Ecosystem size and complexity dictate riverine biodiversity. bioRxiv 2021. [Google Scholar] [CrossRef]
- Sundberg, C.; Al-Soud, W.A.; Larsson, M.; Alm, E.; Yekta, S.S.; Svensson, B.H.; Sørensen, S.J.; Karlsson, A. 454 pyrosequencing analyses of bacterial and archaeal richness in 21 full-scale biogas digesters. FEMS Microbiol. Ecol. 2013, 85, 612–626. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Clemente, J.C.; Kuczynski, J.; Rideout, J.R.; Stombaugh, J.; Wendel, D.; Wilke, A.; Huse, S.; Hufnagle, J.; Meyer, F.; et al. The Biological Observation Matrix (BIOM) format or: How I learned to stop worrying and love the ome-ome. GigaScience 2012, 1, 2047–217X. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.; Xu, Z.Z.; Peddada, S.; Amir, A.; Bittinger, K.; Gonzalez, A.; Lozupone, C.; Zaneveld, J.R.; Vázquez-Baeza, Y.; Birmingham, A.; et al. Normalization and microbial differential abundance strategies depend upon data characteristics. Microbiome 2017, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S rRNA Gene Database and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Champaign, IL, USA, 1949. [Google Scholar]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Chao, A.; Lee, S.M. Estimating the number of classes via sample coverage. J. Am. Stat. Assoc. 1992, 87, 210–217. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Bittinger, K.; Charlson, E.S.; Hoffmann, C.; Lewis, J.; Wu, G.D.; Collman, R.G.; Bushman, F.D.; Li, H. Associating microbiome composition with environmental covariates using generalized UniFrac distances. Bioinformatics 2012, 28, 2106–2113. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Knight, R. Global Patterns in Bacterial Diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 11436–11440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorensen, T.A. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons. Biol. Skar. 1948, 5, 1–34. [Google Scholar]
- Legendre, P.; De Cáceres, M. Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef]
- Jaccard, P. Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Cover, T.; Hart, P. Nearest neighbor pattern classification. IEEE Trans. Inf. Theory 1967, 13, 21–27. [Google Scholar] [CrossRef]
- Vázquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A tool for visualizing high-throughput microbial community data. GigaScience 2013, 2, 2047–217X. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 22 February 2021).
- Kersters, K.; De Vos, P.; Gillis, M.; Swings, J.; Vandamme, P.; Stackebrandt, E. Introduction to the Proteobacteria. In The Prokaryotes; Dworkin, M., Falkow, S., Rosemberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 3–37. [Google Scholar]
- Michán, C.; Blasco, J.; Alhama, J. High-throughput molecular analyses of microbiomes as a tool to monitor the wellbeing of aquatic environments. Microb. Biotechnol. 2021, 14, 870–885. [Google Scholar] [CrossRef] [PubMed]
- Anandan, R.; Dharumadurai, D.; Manogaran, G.P. An introduction to Actinobacteria. In Actinobacteria—Basics and Biotechlological Applications; Dharumadurai, D., Jiang, Y., Eds.; IntechOpen: Tamil Nadu, India, 2016. [Google Scholar] [CrossRef] [Green Version]
- Hashmi, I.; Bindschedler, S.; Junier, P. Firmicutes Chapter in Beneficial Microbes in Agro-Ecology, Bacteria and Fungi; Academic Press: Cambridge, MA, USA, 2020; pp. 336–396. [Google Scholar]
- Speirs, L.B.M.; Rice, D.T.F.; Petrovski, S.; Seviour, R.J. The Phylogeny, Biodiversity, and Ecology of the Chloroflexi in Activated Sludge. Front. Microbiol. 2019, 10, 2015. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.; Qu, X.; Peng, W.; Yu, Y.; Zhang, M. Nutrients Drive the Structures of Bacterial Communities in Sediments and Surface Waters in the River-Lake System of Poyang Lake. Water 2019, 11, 930. [Google Scholar] [CrossRef] [Green Version]
- Lee, Z.M.-P.; Poret-Peterson, A.; Siefert, J.L.; Kaul, D.; Moustafa, A.; Allen, A.; Dupont, C.L.; Eguiarte, L.E.; Souza, V.; Elser, J.J. Nutrient Stoichiometry Shapes Microbial Community Structure in an Evaporitic Shallow Pond. Front. Microbiol. 2017, 8, 949. [Google Scholar] [CrossRef]
- Zeglin, L.H. Stream microbial diversity in response to environmental changes: Review and synthesis of existing research. Front. Microbiol. 2015, 6, 454. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| River Sector | Nitrates (μg/g) | Ammonium (μg/g) | Orthophosphates (μg/g) |
|---|---|---|---|
| M1 | 1.586402 | 35.63673 | 30.67308 |
| M2 | 3.869318 | 5.922449 | 16.88462 |
| M4 | 1.726912 | 23.18776 | 25.38462 |
| M6 | 5.614232 | 30.82041 | 25.14423 |
| M7 | 0.522472 | 14.53469 | 33.31731 |
| M8 | 3.888952 | 9.432653 | 40.09615 |
| M9 | 3.928839 | 33.47347 | 29.90385 |
| M10 | 6.625468 | 37.3102 | 46.97115 |
| M11 | 1.629462 | 2.249388 | 102.5962 |
| M12 | 4.846442 | 156.3673 | 60 |
| M14 | 3.415297 | 21.71837 | 38.60577 |
| M15 | 0.463483 | 1.449388 | 17.01923 |
| M16 | 7.6875 | 16.12653 | 37.25962 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boeraş, I.; Burcea, A.; Coman, C.; Bănăduc, D.; Curtean-Bănăduc, A. Bacterial Microbiomes in the Sediments of Lotic Systems Ecologic Drivers and Role: A Case Study from the Mureş River, Transylvania, Romania. Water 2021, 13, 3518. https://doi.org/10.3390/w13243518
Boeraş I, Burcea A, Coman C, Bănăduc D, Curtean-Bănăduc A. Bacterial Microbiomes in the Sediments of Lotic Systems Ecologic Drivers and Role: A Case Study from the Mureş River, Transylvania, Romania. Water. 2021; 13(24):3518. https://doi.org/10.3390/w13243518
Chicago/Turabian StyleBoeraş, Ioana, Alexandru Burcea, Cristian Coman, Doru Bănăduc, and Angela Curtean-Bănăduc. 2021. "Bacterial Microbiomes in the Sediments of Lotic Systems Ecologic Drivers and Role: A Case Study from the Mureş River, Transylvania, Romania" Water 13, no. 24: 3518. https://doi.org/10.3390/w13243518
APA StyleBoeraş, I., Burcea, A., Coman, C., Bănăduc, D., & Curtean-Bănăduc, A. (2021). Bacterial Microbiomes in the Sediments of Lotic Systems Ecologic Drivers and Role: A Case Study from the Mureş River, Transylvania, Romania. Water, 13(24), 3518. https://doi.org/10.3390/w13243518