
Changing Water Levels in Lake Superior, MI (USA) Impact Periphytic Diatom Assemblages in the Keweenaw Peninsula



Abstract
:1. Introduction
2. Materials and Methods
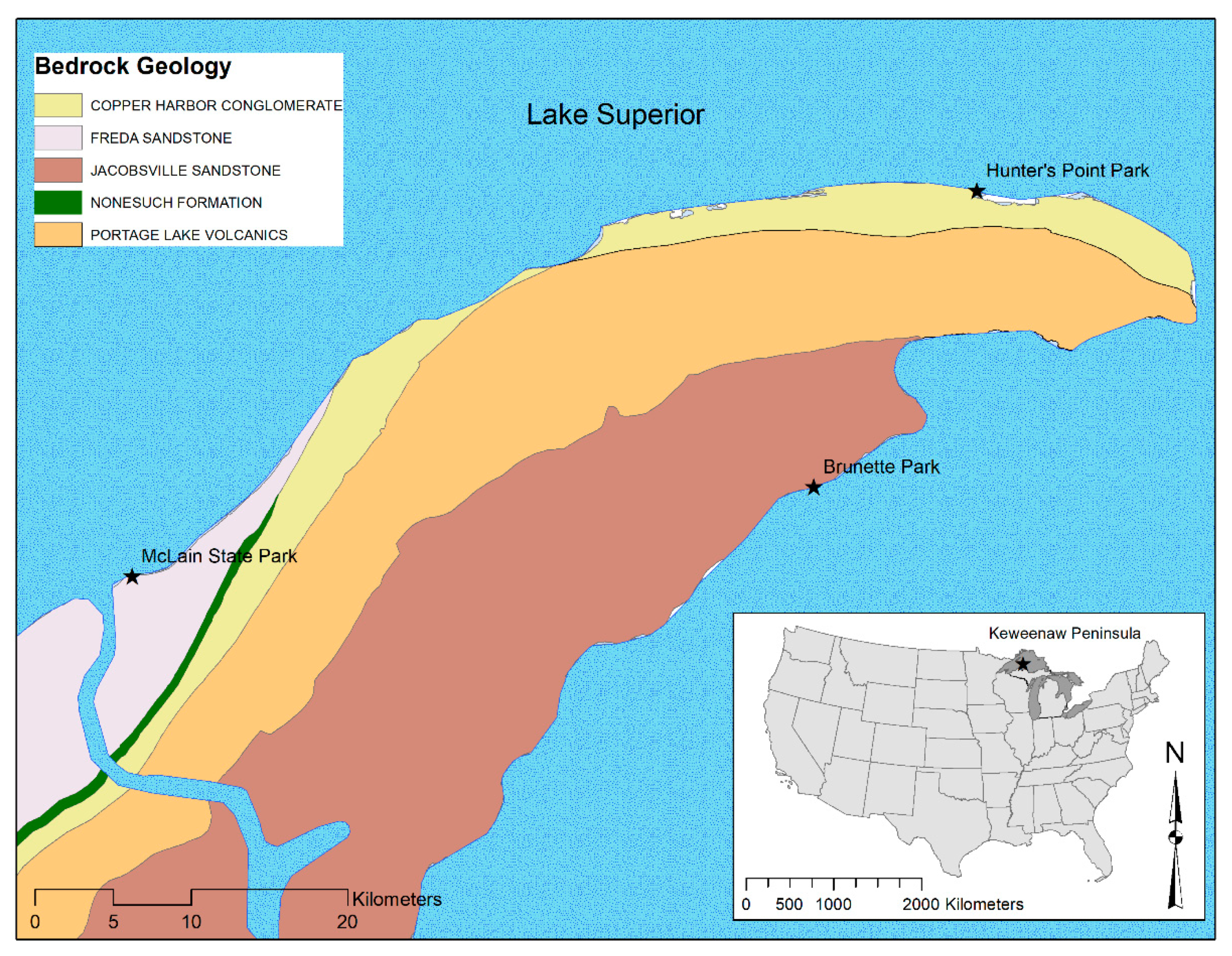
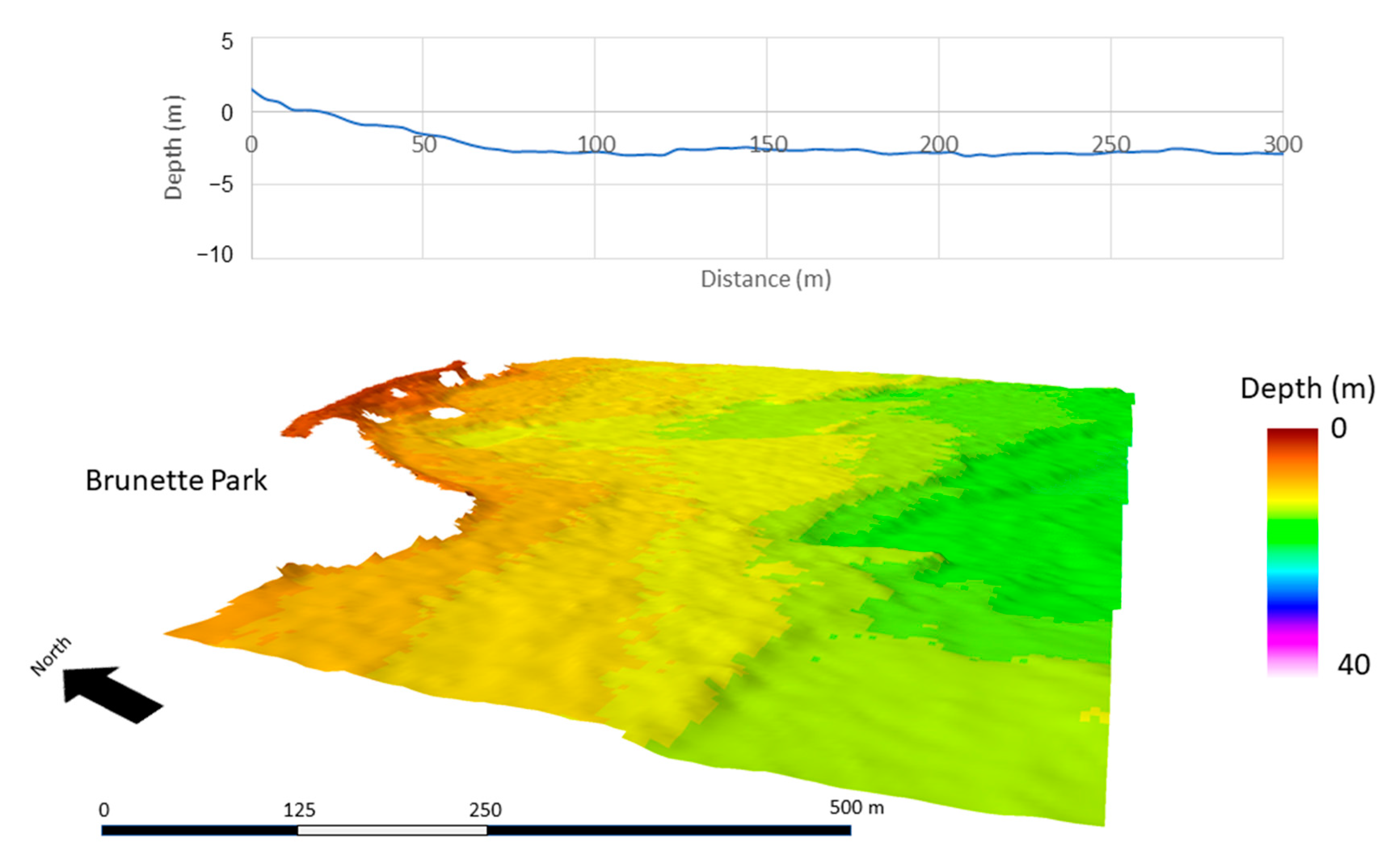
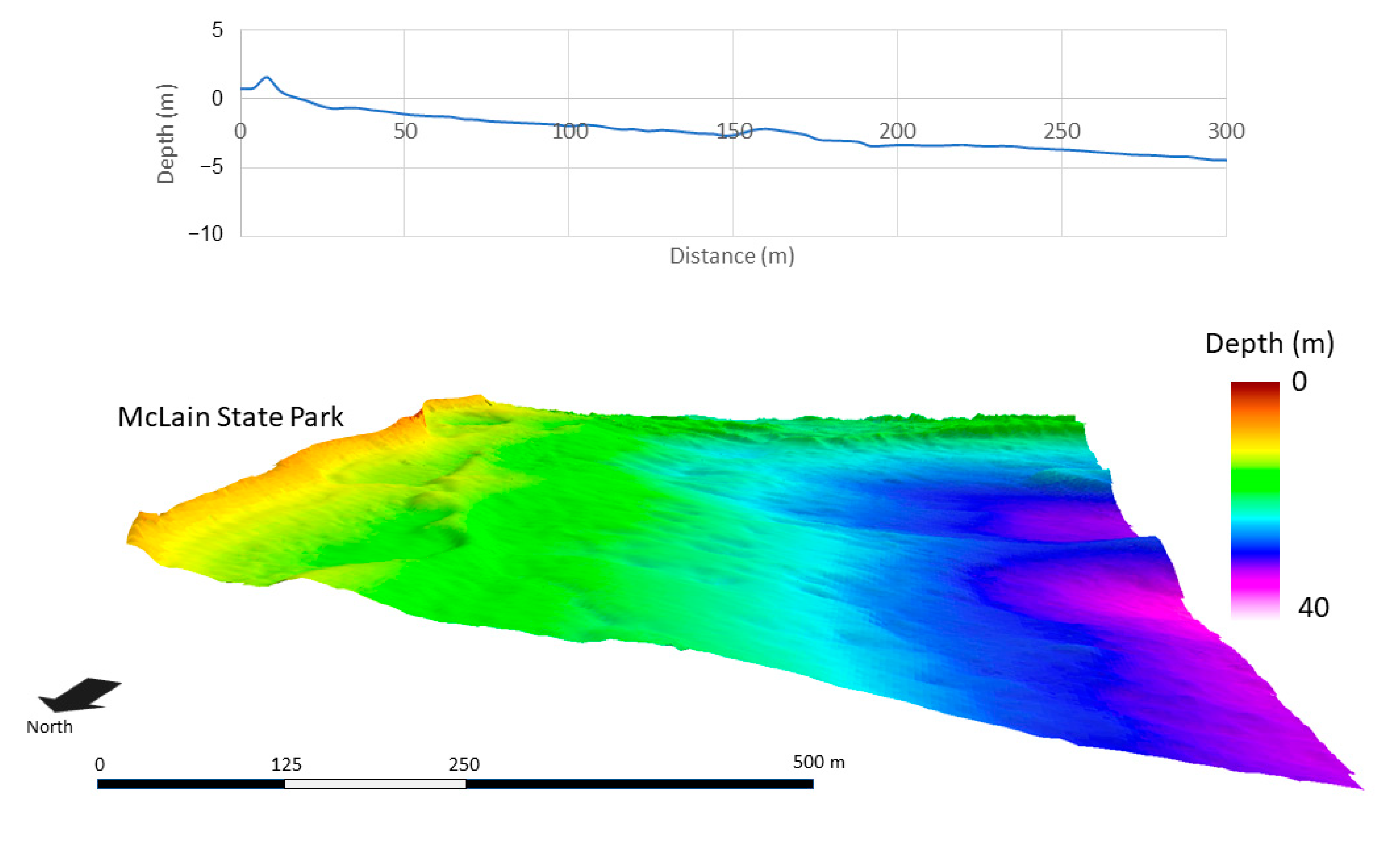
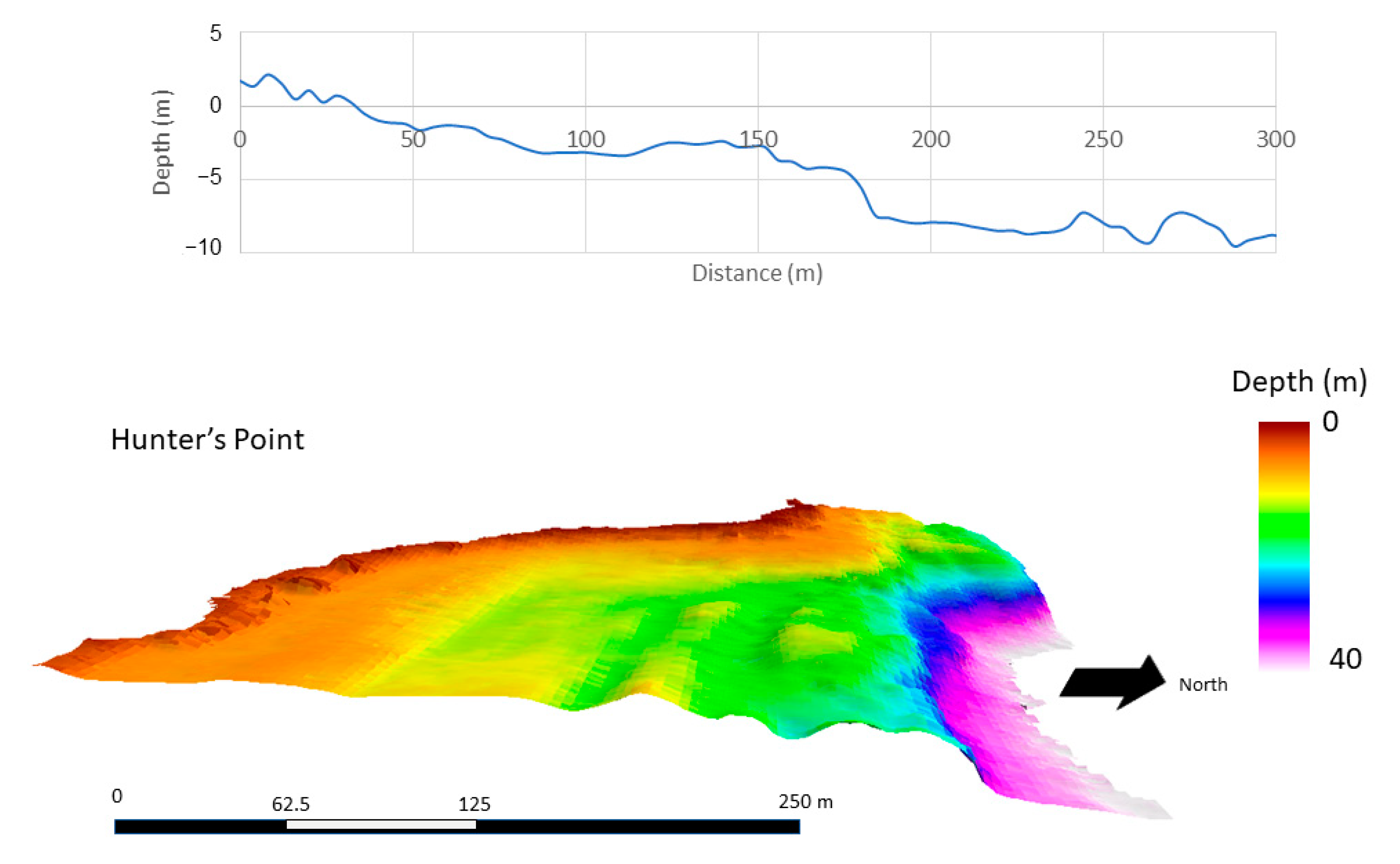
2.1. Site Location
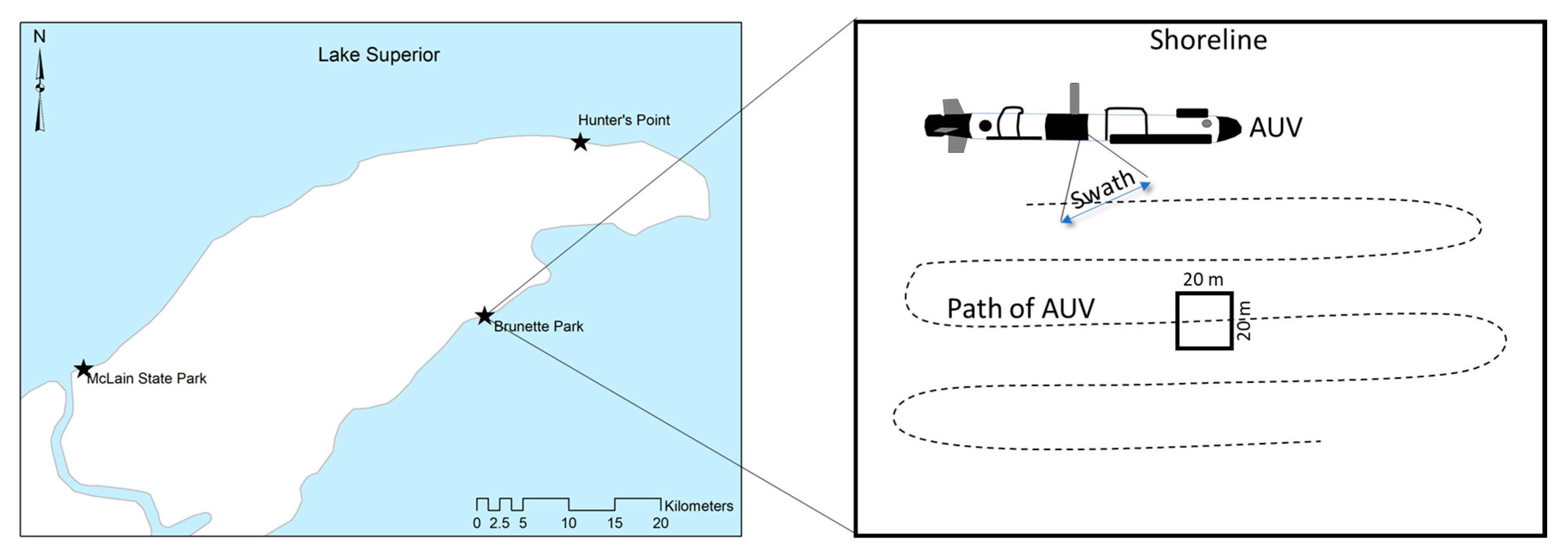
2.2. Sample Collection
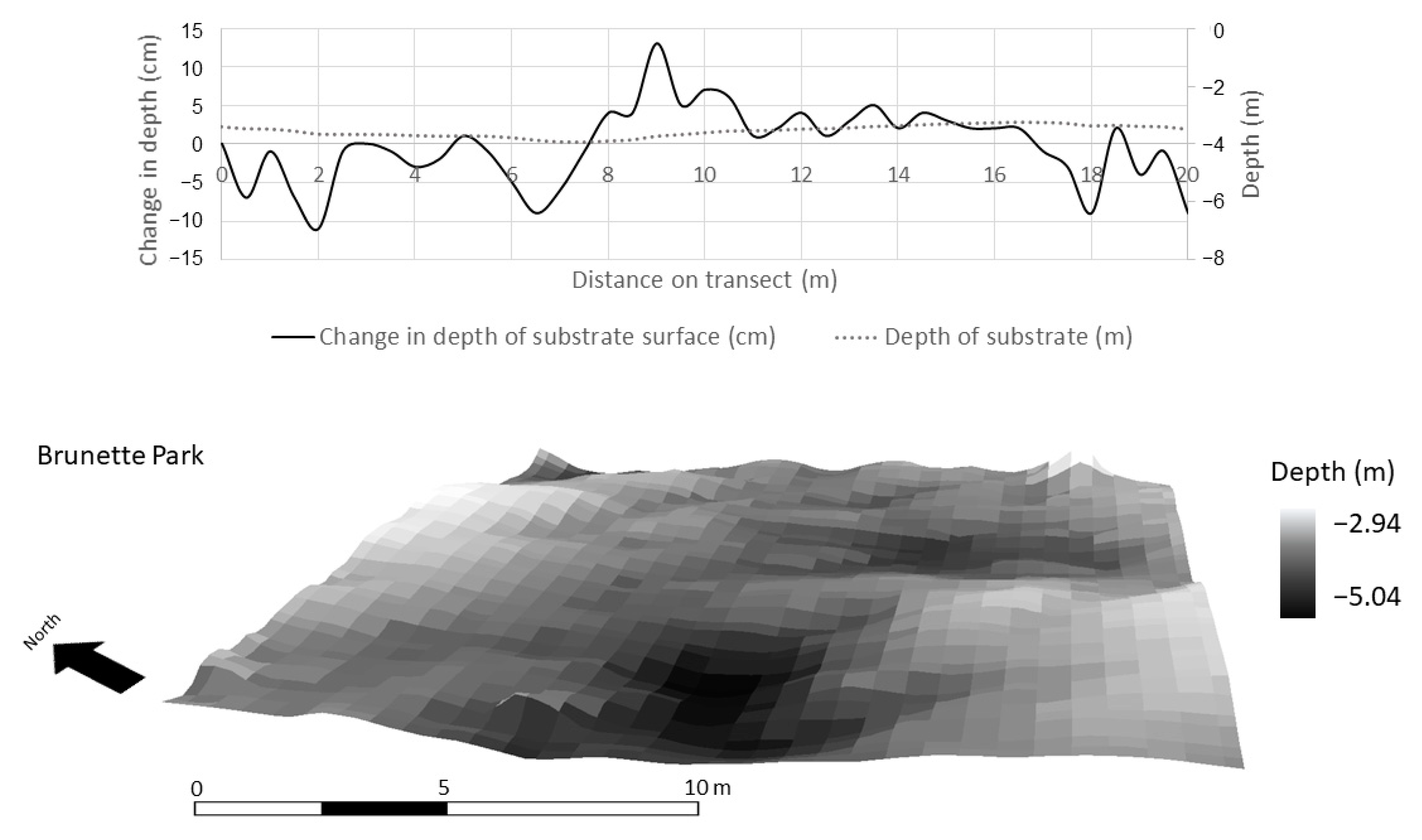
2.3. Quantitative and Spatial Analyses
3. Results
3.1. Physical and Chemical Data
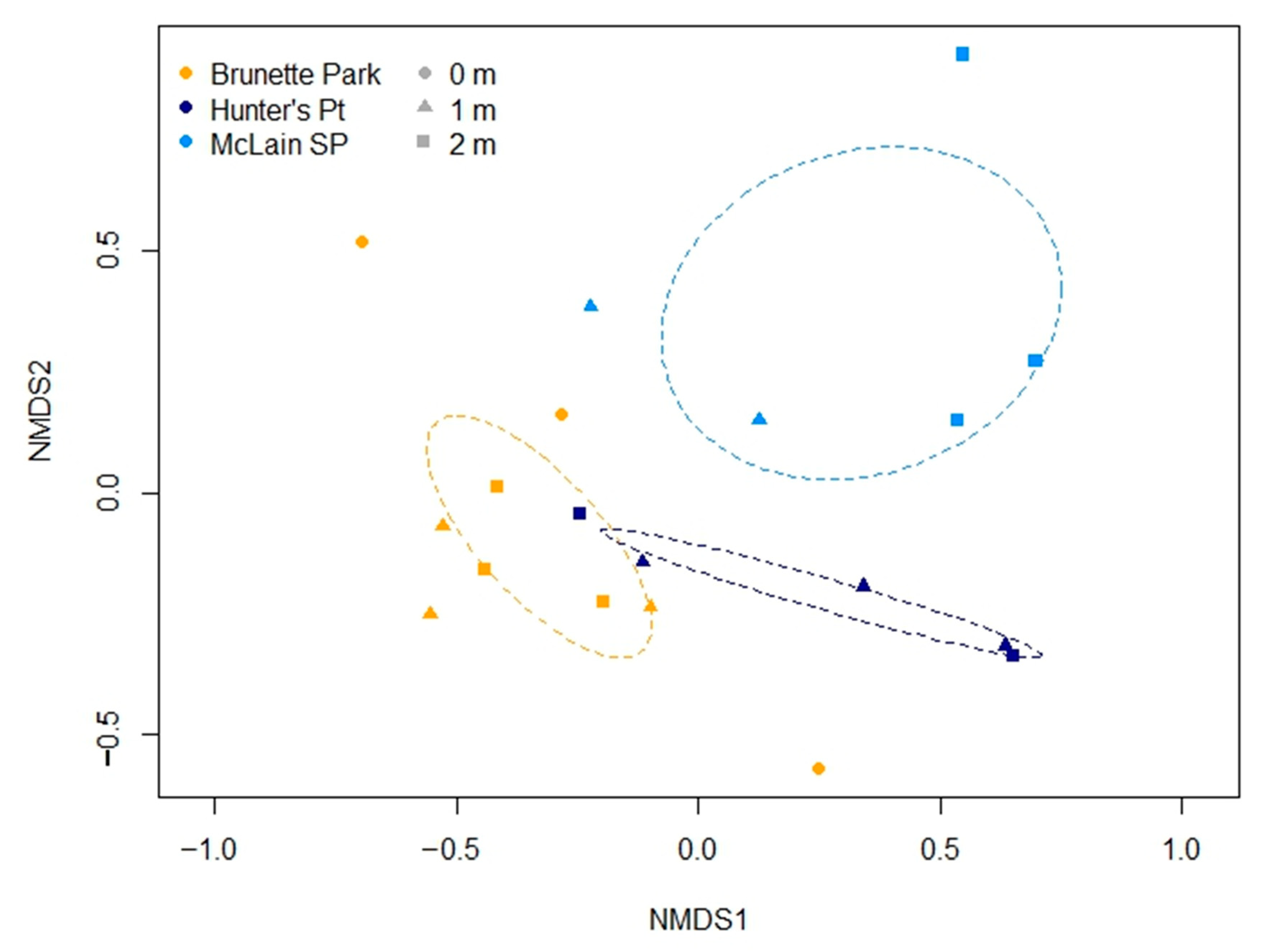
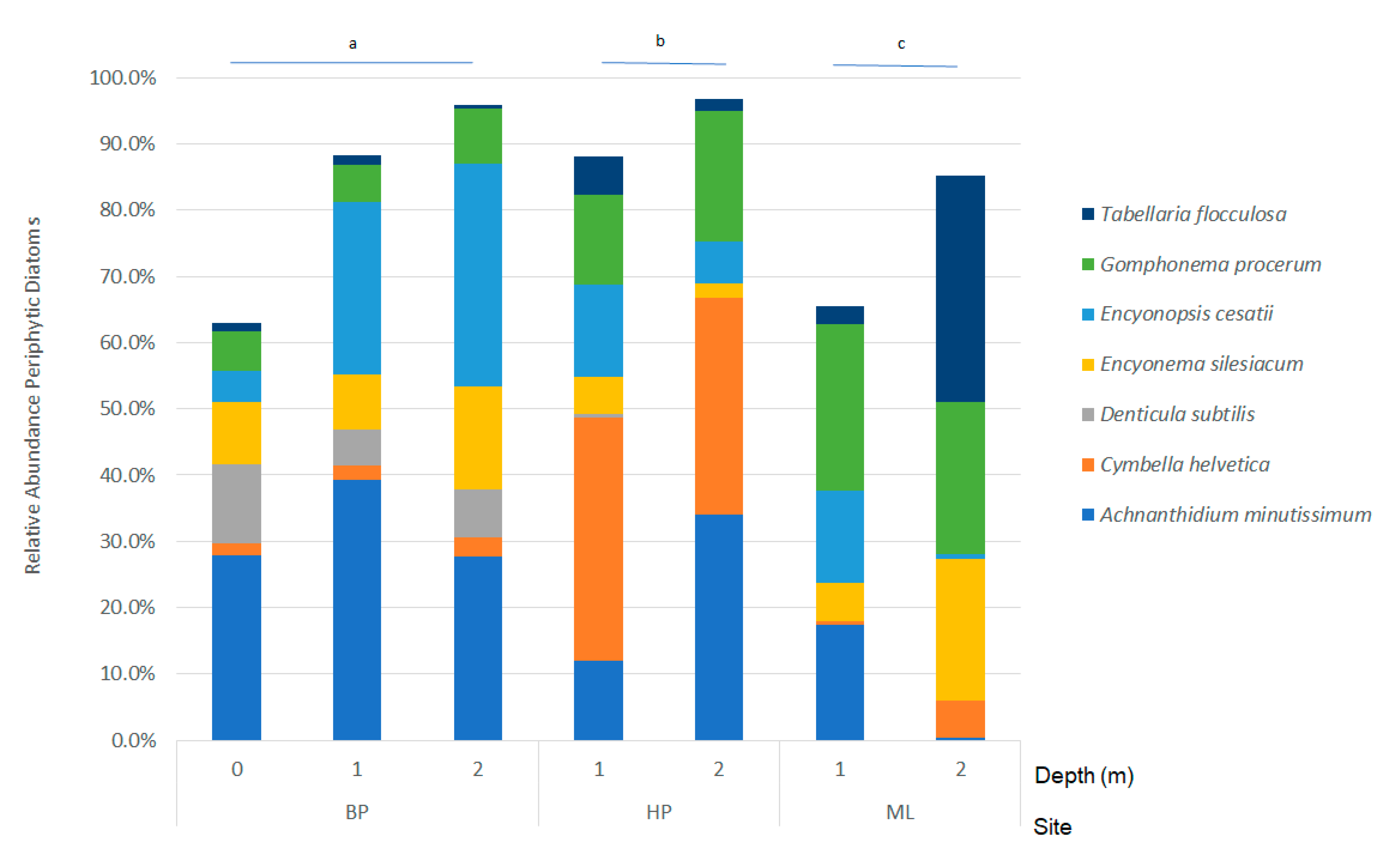
3.2. Periphytic Diatom Assemblages
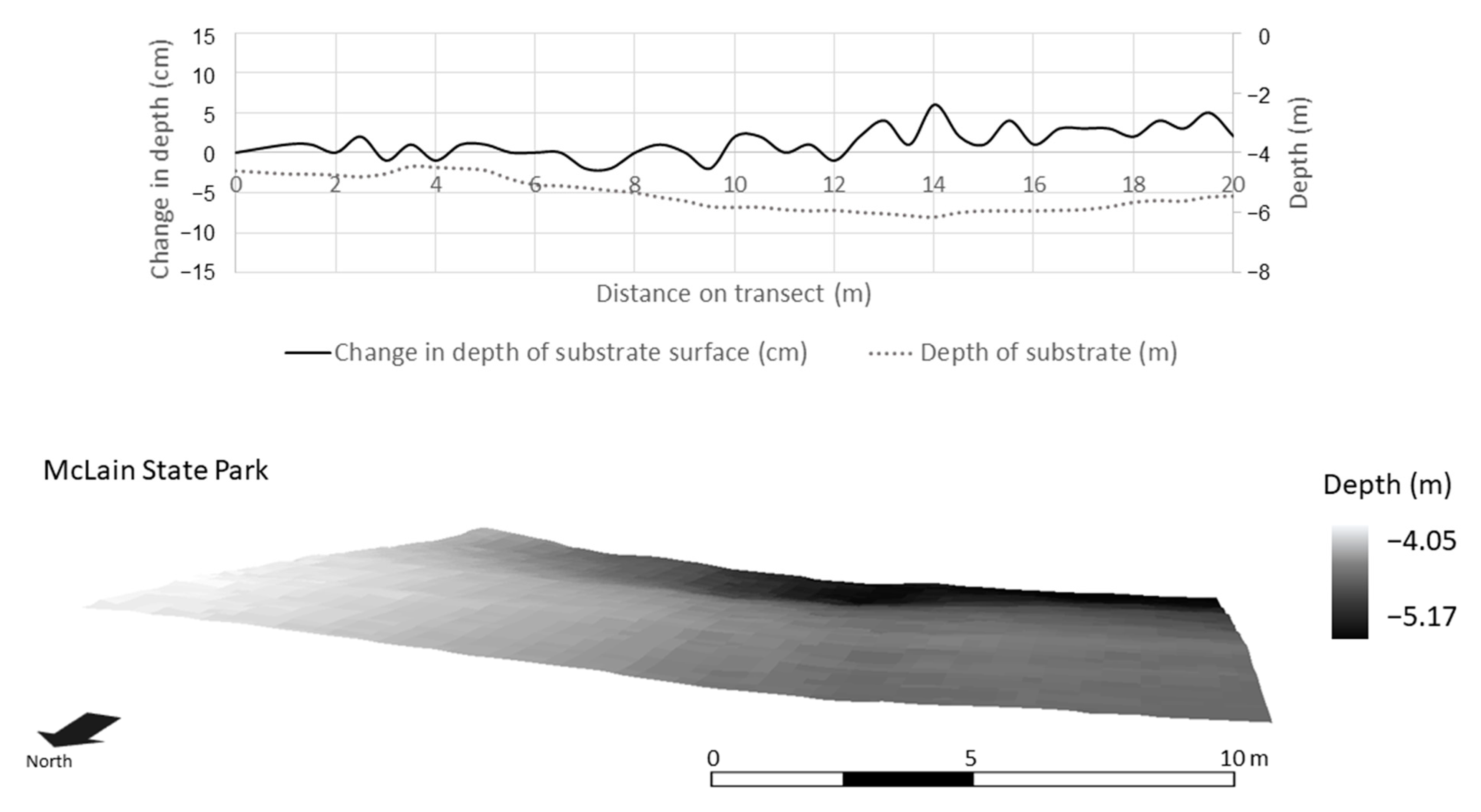
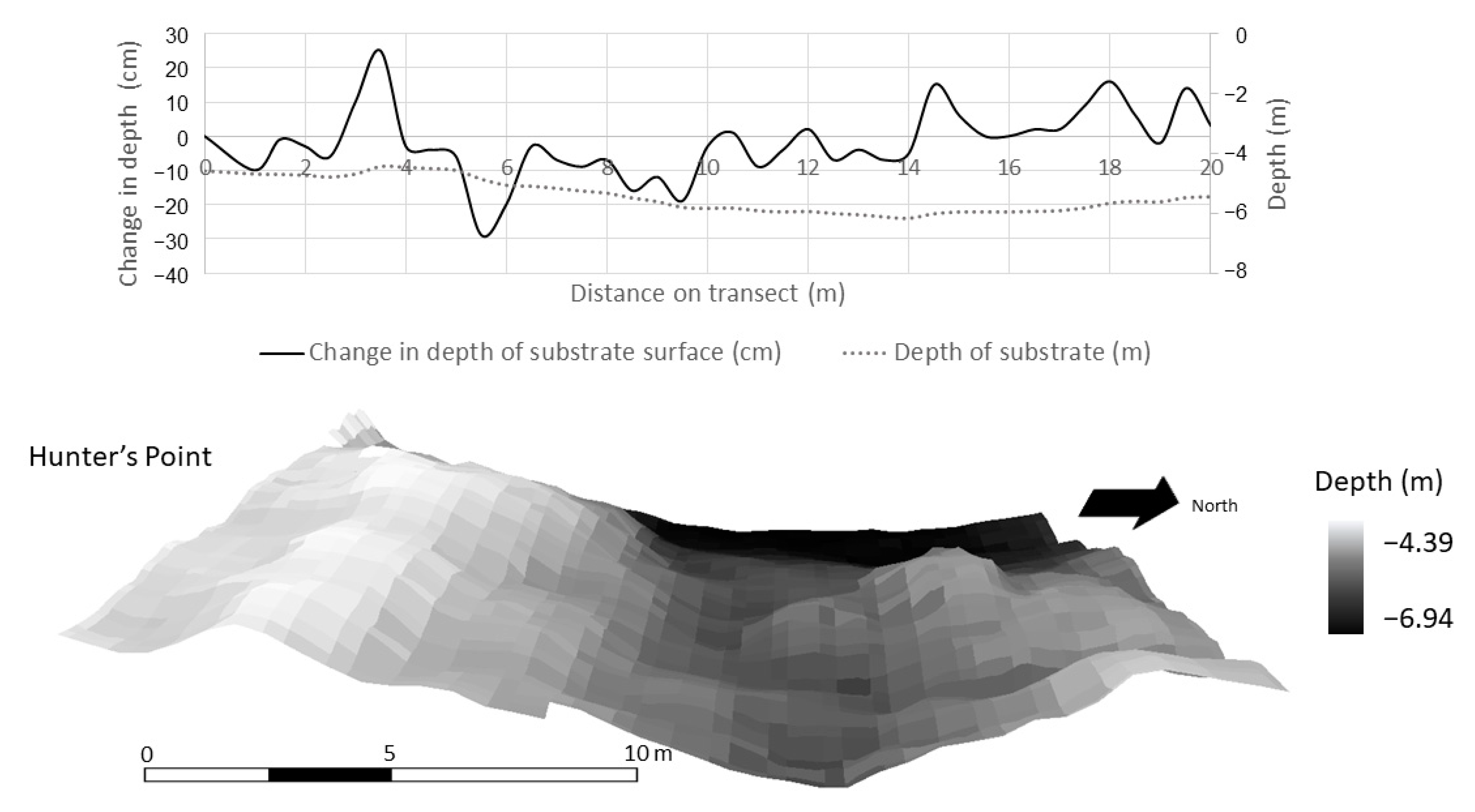
3.3. Spatial Modeling and Habitat Structure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Great Lakes Integrated Sciences and Assessments (GLISA). Annual Climate Trends and Impacts Summary for the Great Lakes Basin. Available online: http://glisa.umich.edu/resources/annual-climate-summary (accessed on 23 November 2020).
- Gronewald, A.D.; Hunter, T.S.; Allison, J.; Fry, L.M.; Kompoltowicz, K.A.; Bolinger, R.A.; Pei, L. Project Documentation Report for Great Lakes Seasonal and Inter-Annual Water Supply Forecasting Improvements Project Phase I: Research and Development; NOAA Great Lakes Environmental Research Laboratory, U.S. Army Corps of Engineers Detroit District and the University Corporation for Atmospheric Research: Ann Arbor, MI, USA, 2017.
- United States Army Corps of Engineers. Great Lakes Water Level Data. Available online: https://www.lre.usace.army.mil/Missions/Great-Lakes-Information/Great-Lakes-Information-2/Water-Level-Data/ (accessed on 21 November 2020).
- Nicholls, K.H. Effects of temperature and other factors on summer phosphorus in the inner Bay of Quinte, Lake Ontario: Implications for climate warming. J. Great Lakes Res. 1999, 25, 250–262. [Google Scholar] [CrossRef]
- Smith, A.L.; Hewitt, N.; Klenk, N.; Bazely, D.R.; Yan, N.; Wood, S.; Henriques, I.; MacLellan, J.I.; Lipsig-Mummé, C. Effects of climate change on the distribution of invasive alien species in Canada: A knowledge synthesis of range change projections in a warming world. Environ. Rev. 2012, 20, 1–16. [Google Scholar] [CrossRef]
- Kraemer, B.M.; Anneville, O.; Chandra, S.; Dix, M.; Kuusisto, E.; Livingstone, D.M.; Rimmer, A.; Schladow, S.G.; Silow, E.; Sitoki, L.M.; et al. Morphometry and average temperature affect lake stratification responses to climate change. Geophys. Res. Lett. 2015, 42, 4981–4988. [Google Scholar] [CrossRef] [Green Version]
- Mason, L.A.; Riseng, C.M.; Gronewold, A.D.; Rutherford, E.S.; Wang, J.; Clites, A.; Smith, S.D.P.; McIntyre, P.B. Fine-scale spatial variation in ice cover and surface temperature trends across the surface of the Laurentian Great Lakes. Clim. Chang. 2016, 138, 71–83. [Google Scholar] [CrossRef]
- Lowe, R.L.; Pan, Y. Benthic Algal Communities and Biological Monitors. In Algal Ecology: Freshwater Benthic Ecosystems; Stevenson, R.J., Bothwell, M.L., Lowe, R.L., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 705–739. [Google Scholar]
- Kingston, J.C.; Lowe, R.L.; Stoermer, E.; Ladewski, T.B. Spatial and temporal distribution of benthic diatoms in northern Lake Michigan. Ecology 1983, 64, 1566. [Google Scholar] [CrossRef]
- Hagy, J.D., III; Houthon, K.A.; Beddick, D.L., Jr.; James, J.B. Quantifying stream periphyton assemblage responses to nutrient amendments with a molecular approach. Freshw. Sci. 2020, 39, 292–308. [Google Scholar] [CrossRef]
- Ledger, M.E.; Hildrew, A.G. Temporal and spatial variation in the epilithic biofilm of an acid stream. Freshw. Biol. 1998, 40, 655–670. [Google Scholar] [CrossRef]
- Sanson, G.; Stolk, R.; Downes, B. A New Method for Characterizing Surface Roughness and Available Space in Biological Systems. Funct. Ecol. 1995, 9, 127–135. [Google Scholar] [CrossRef]
- Bergey, E.A. How protective are refuges? Quantifying algal protection in rock crevices. Freshw. Biol. 2005, 50, 1163–1177. [Google Scholar] [CrossRef]
- Burkholder, J.M. Interactions of benthic algae with their substrata. In Algal Ecology: Freshwater Benthic Ecosystems; Stevenson, R.J., Bothwell, M.L., Lowe, R.L., Eds.; Academic Press: London, UK, 1996; pp. 253–298. [Google Scholar]
- Murdock, J.N.; Dodds, W.K. Linking benthic algal biomass to stream substratum topography 1. J. Phycol. 2007, 43, 449–460. [Google Scholar] [CrossRef]
- Souza, M.L.D.; Ferragut, C. Influence of substratum surface roughness on periphytic algal community structure in a shallow tropical reservoir. Acta Limnologica Brasiliensia 2012, 24, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Great Lakes Environmental Research Lab: Great Lakes Dashboard. Available online: https://www.glerl.noaa.gov/data/dashboard/GLD_HTML5.html (accessed on 21 November 2020).
- Lofgren, B.M.; Hunter, T.S.; Wilbarger, J. Effects of using air temperature as a proxy for potential evapotranspiration in climate change scenarios of Great Lakes basin hydrology. JGLR 2011, 37, 744–752. [Google Scholar] [CrossRef]
- DeGraff, J.; Department of Geological and Mining Engineering Sciences, Michigan Technological Institute, Houghton, MI, USA. Personal communication, 2020.
- Yousef, F.; Shuchman, R.; Sayers, M.; Fahnenstiel, G.; Henareh, A. Water clarity of the Upper Great Lakes: Tracking changes between 1998–2012. J. Great Lakes Res. 2017, 43, 239–247. [Google Scholar] [CrossRef]
- Stevenson, R.J.; Bahls, L.L. Periphyton Protocols. In Rapid Bioassessment Protocols for Use in Wadeable Streams and Rivers: Periphyton, Benthic Macroinvertebrates, and Fish, 2nd ed.; Barbour, M.T., Gerritsen, J., Snyder, B.D., Stribling, J.B., Eds.; U.S. Environmental Protection Agency: Washington, DC, USA, 1999. [Google Scholar]
- Carr, J.M.; Hergenrader, G.L.; Troelstrup, N.H., Jr. A simple, inexpensive method for cleaning diatoms. Trans. Am. Microsc. Soc. 1986, 105, 152–157. [Google Scholar] [CrossRef]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae. 1 Teil: Naviculaceae. In Süsswasserflora von Mitteleuropa 1986, Band 2/1; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fisher Verlag: Jena, Germany, 1986; p. 876. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae, 2 Teil: Bacillariaceae. Epithemiaceae, Surirellaceae. In Süsswasserflora von Mitteleuropa; IEttl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fisher Verlag: Jena, Germany, 1988; Volume 2, pp. 1–596. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae, 3 Teil: Centrales, Fragilariaceae, Eunotiaceae. In Süsswasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fisher Verlag: Jena, Germany, 1991; Volume 2, pp. 1–576. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae, 4 Teil: Achnanthaceae, Kritische Erganzungen zu Navicula (Lineolatae) und Gomphonema, Gesamtliteraturverzeichnis Teil 1–4. In Süsswasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fisher Verlag: Jena, Germany, 1991; Volume 2, pp. 1–437. [Google Scholar]
- Diatoms of North America. Available online: https://diatoms.org/ (accessed on 20 December 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 21 November 2020).
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 2nd ed.; Sage: Thousand Oaks, CA, USA, 2011; Available online: http://socserv.socsci.mcmaster.ca/jfox/Books/Companion (accessed on 21 November 2020).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package 2017, Version 2.4-3. Available online: https://CRAN.R-project.org/package=vegan (accessed on 21 November 2020).
- Environmental Systems Research Institute (ESRI). ArcGIS v 10.4.1; Redlands, CA, USA, 2016. Available online: https://www.esri.com/en-us/home (accessed on 21 November 2020).
- National Oceanic and Atmospheric Association (NOAA). 2019 USACE NCMP Topobathy Lidar DEM: Lake Superior (MI). Available online: https://coast.noaa.gov/dataviewer/#/lidar/search/-9867867.477559684,5977452.048929067,-9755046.423810763,6024384.384296166/details/9185 (accessed on 21 November 2020).
- Angel, J.R.; Kunkel, K.E. The response of Great Lakes water levels to future climate scenarios with an emphasis on Lake Michigan-Huron. J. Great Lakes Res. 2010, 36, 51–58. [Google Scholar] [CrossRef]
- Hayhoe, K.; VanDorn, J.; Croley II, T.; Schlegel, N.; Wuebbles, D. Regional climate change projections for Chicao and the US Great Lakes. J. Great Lakes Res. 2010, 36, 7–21. [Google Scholar] [CrossRef]
- Bergey, E.A. Crevices as refugia for stream diatoms: Effect of crevice size on abraded substrates. Limnol. Oceanogr. 1999, 44, 1522–1529. [Google Scholar] [CrossRef]
- Kralj, K.; Plenković-Moraj, A.; Gligora, M.; Primc-Habdija, B.; Šipoš, L. Structure of periphytic community on artificial substrata: Influence of depth, slide orientation and colonization time in karstic Lake Visovačko, Croatia. Hydrobiologia 2006, 560, 249–258. [Google Scholar] [CrossRef]
- Johnson, L.E. Enhanced settlement on microtopographical high points by the intertidal red alga Halosaccion glandiforme. Limnol. Oceanogr. 1994, 39, 1893–1902. [Google Scholar] [CrossRef]
- Sekar, R.; Venugopalan, V.P.; Satpathy, K.K.; Nair, K.V.K.; Rao, V.N.R. Laboratory Studies on Adhesion of Microalgae to Hard Substrates. In Asian Pacific Phycology in the 21st Century: Prospects and Challenges; Springer: Dordrecht, The Netherlands, 2004; pp. 109–116. [Google Scholar]
- DeNicola, D.M.; McIntire, C.D. Effects of substrate relief on the distribution of periphyton in laboratory streams, i. hydrology 1. J. Phycol. 1990, 26, 624–633. [Google Scholar] [CrossRef]
- Biggs, B.J.F.; Thomsen, H.A. Disturbance of stream periphyton by perturbations in shear stress: Time to structural failure and differences in community resistance. J. Phycol. 1995, 31, 233–241. [Google Scholar] [CrossRef]
- Glasby, T.M.; Connell, S.D. Orientation and position of substrata have large effects on epibiotic assemblages. Mar. Ecol. Prog. Series 2001, 214, 127–135. [Google Scholar] [CrossRef]
- Steinman, A.D. Effects of Grazers on Freshwater Benthic Algae. In Algal Ecology: Freshwater Benthic Ecosystems; Stevenson, R.J., Bothwell, M.L., Lowe, R.L., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 341–373. [Google Scholar]
- Thomas, E.W. The Role of Wave Disturbance on Lentic, Benthic Algal Community Structure and Diversity. Master’ Thesis, Bowling Green State University, Bowling Green, OH, USA, August 2007. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Minimum (m) | Maximum (m) | Reference | |
|---|---|---|---|
| Historic Data (1918–2020) | 182.72 | 183.91 | Great Lakes Dashboard [17] |
| Current Study (2019) | 183.86 | ||
| Future Projections (2068–2105) | 182.42 | 183.74 | Lofgren et al. [18] |
| 183.22 | 183.47 | Angel et al. [34] | |
| 183.24 | 183.27 | Hayhoe et al. [35] |
| Site/ Depth (m) | Nitrate (mg L−1) | Total Phosphorus (ug L−1) | Temperature (C) | Dissolved Oxygen (mg L−1) | Conductivity (μs cm−1) |
|---|---|---|---|---|---|
| BP | 0.33 ± 0.00 (9) | 52.89 ± 3.94 (9) | 19.78 ± 0.27 (9) | 11.59 ± 0.40 (9) | 88.02 ± 0.38 (9) |
| 0 | 0.32 ± 0.01 (3) | 50.67 ± 2.60 (3) | 20.57 ± 0.18 (3) | 10.50 ± 0.14 (3) | 88.67 ± 0.78 (3) |
| 1 | 0.33 ± 0.01 (3) | 57.33 ± 6.89 (3) | 19.30 ± 0.61 (3) | 11.72 ± 0.56 (3) | 87.97 ± 0.80 (3) |
| 2 | 0.33 ± 0.00 (3) | 50.67 ± 10.84 (3) | 19.47 ± 0.09 (3) | 12.54 ± 0.70 (3) | 87.43 ± 0.32 (3) |
| HP | 0.33 ± 0.01 (9) | 50.67 ± 3.84 (9) | 21.30 ± 0.14 (9) | 10.17 ± 0.25 (9) | 91.51 ± 0.22 (3) |
| 0 | 0.35 ± 0.01 (3) | 55.00 ± 7.55 (3) | 21.63 ± 0.30 (3) | 9.22 ± 0.14 (3) | 90.97 ± 0.29 (3) |
| 1 | 0.31 ± 0.02 (3) | 46.00 ± 8.89 (3) | 21.30 ± 0.15 (3) | 10.83 ± 0.26 (3) | 92.23 ± 0.23 (3) |
| 2 | 0.32 ± 0.00 (3) | 51.00 ± 4.58 (3) | 20.97 ± 0.09 (3) | 10.47 ± 0.09 (3) | 91.33 ± 0.15 (3) |
| ML | 0.32 ± 0.00 (9) | 58.11 ± 5.09 (9) | 20.88 ± 0.61 (9) | 9.88 ± 0.13 (9) | 93.42 ± 1.73 (3) |
| 0 | 0.31 ± 0.00 (3) | 65.33 ± 8.35 (3) | 23.30 ± 0.11 (3) | 9.61 ± 0.06 (3) | 97.03 ± 4.60 (3) |
| 1 | 0.32 ± 0.00 (3) | 49.67 ± 3.48 (3) | 19.90 ± 0.11 (3) | 9.77 ± 0.06 (3) | 93.07 ± 0.98 (3) |
| 2 | 0.32 ± 0.01 (3) | 59.33 ± 12.91 (3) | 19.43 ± 0.12 (3) | 10.25 ± 0.28 (3) | 89.90 ± 0.23 (3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woller-Skar, M.M.; Locher, A.; Audia, E.; Thomas, E.W. Changing Water Levels in Lake Superior, MI (USA) Impact Periphytic Diatom Assemblages in the Keweenaw Peninsula. Water 2021, 13, 253. https://doi.org/10.3390/w13030253
Woller-Skar MM, Locher A, Audia E, Thomas EW. Changing Water Levels in Lake Superior, MI (USA) Impact Periphytic Diatom Assemblages in the Keweenaw Peninsula. Water. 2021; 13(3):253. https://doi.org/10.3390/w13030253
Chicago/Turabian StyleWoller-Skar, M. Megan, Alexandra Locher, Ellen Audia, and Evan W. Thomas. 2021. "Changing Water Levels in Lake Superior, MI (USA) Impact Periphytic Diatom Assemblages in the Keweenaw Peninsula" Water 13, no. 3: 253. https://doi.org/10.3390/w13030253
APA StyleWoller-Skar, M. M., Locher, A., Audia, E., & Thomas, E. W. (2021). Changing Water Levels in Lake Superior, MI (USA) Impact Periphytic Diatom Assemblages in the Keweenaw Peninsula. Water, 13(3), 253. https://doi.org/10.3390/w13030253