
Multi-Biomarker Responses of Asian Clam Corbicula fluminea (Bivalvia, Corbiculidea) to Cadmium and Microplastics Pollutants


 , ,
, ,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Experimental Design
2.3. Cd Quantification in Water and Bivalve Organs
2.4. MPs Scanning Electron Microscopy plus Energy-Dispersive X-Ray Spectroscopy (SEM/EDS) Analysis
2.5. Biochemical Analyses
2.6. Statistical Analyses
3. Results
3.1. Cadmium Accumulation
3.2. Characterization of MPs Particles
3.3. Oxidative Stress
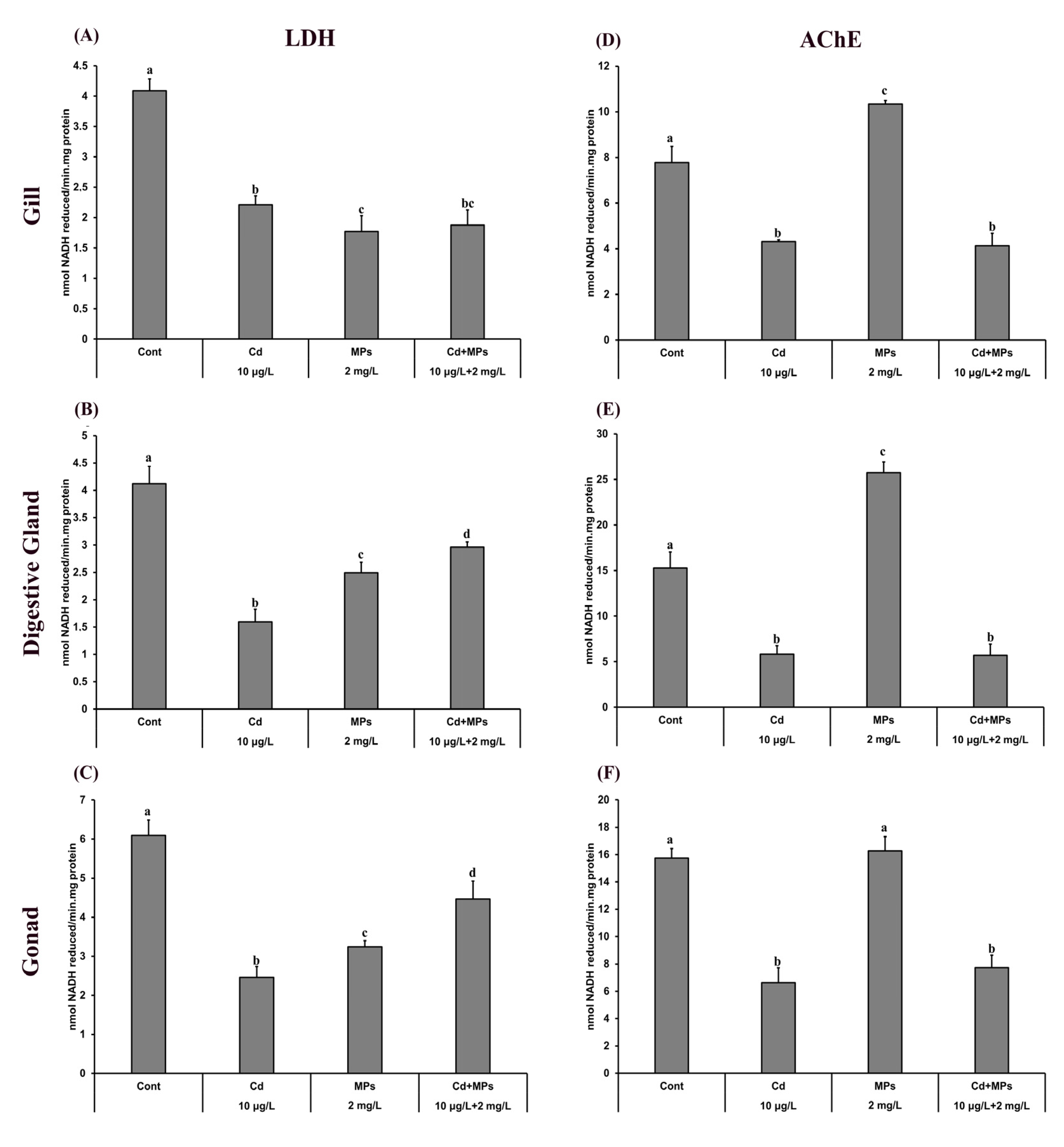
3.4. Metabolic Changes
3.5. Neurotoxicity
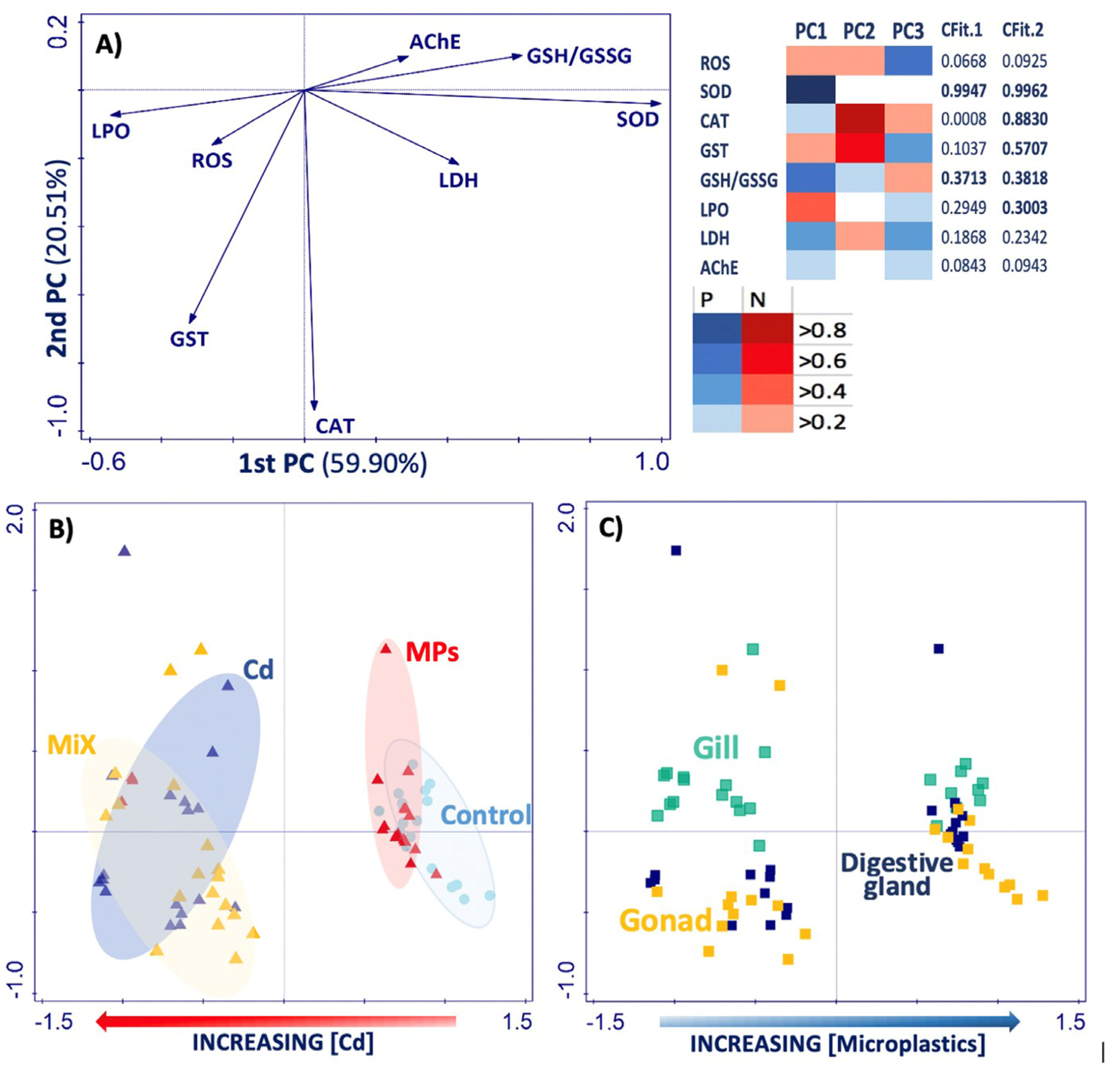
3.6. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Su, L.; Cai, H.; Kolandhasamy, P.; Wu, C.; Rochman, C.R.; Shi, H. Using the Asian clam as an indicator of microplastic pollution in freshwater ecosystems. Environ. Pollut. 2018, 234, 347–355. [Google Scholar] [CrossRef]
- Ortmann, A.E. The destruction of the freshwater fauna in western Pennsylvania. Proc. Am. Philos. Soc. 1909, 48, 90–110. [Google Scholar]
- Boening, D.W. An evaluation of bivalves as biomonitors of heavy metals pollution in marine waters. Environ. Monit. Assess. 1999, 55, 459–470. [Google Scholar] [CrossRef]
- Jara-Marini, M.E.; Tapia-Alcaraz, J.N.; Dumer-Gutierrez, J.A.; García-Rico, L.; García-Hernandez, J.; Paez-Osuna, F. Comparative bioaccumulation of trace metals using six filter feeder organisms in a coastal lagoon ecosystem (of the central-east Gulf of California). Environ. Monit. Assess. 2013, 185, 1071–1085. [Google Scholar] [CrossRef] [PubMed]
- Koch, I.; McPherson, K.; Smith, P.; Easto, L.; Doe, K.G.; Reimer, K.J. Arsenic bioaccessibility and speciation in clams and seaweed from a contaminated marine environment. Mar. Pollut. Bull. 2007, 54, 586–594. [Google Scholar] [CrossRef]
- Rochman, C.M.; Tahir, A.; Williams, S.L.; Baxa, D.V.; Lam, R.; Miller, J.T.; Teh, F.C.; Werorilangi, S.; Teh, S.J. Anthropogenic debris in seafood: Plastic debris and fibers from textiles in fish and bivalves sold for human consumption. Sci. Rep. 2015, 5, 10. [Google Scholar] [CrossRef]
- Seltenrich, N. New link in the food chain? Marine plastic pollution and seafood safety. Environ. Health Perspect. 2015, 123, A34–A41. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Feng, C. Biological toxicity response of Asian Clam (Corbicula fluminea) to pollutants in surface water and sediment. Sci. Total Environ. 2018, 631–632, 56–70. [Google Scholar] [CrossRef]
- Sous, R.; Antunes, C.; Guilhermino, L. Ecology of the invasive Asian clam Corbicula fluminea (Muller, 1774) in aquatic ecosystems: An overview. Ann. Limnol. Int. J. Limnol. 2008, 44, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Rochman, C.M.; Parnis, J.M.; Browne, M.A.; Serrato, S.; Reiner, E.J.; Robson, M.; Young, T.; Diamond, M.L.; Teh, S.J. Direct and indirect effects of different types of microplastics on freshwater prey (Corbicula fluminea) and their predator (Acipenser transmontanus). PLoS ONE 2017, 12, e0187664. [Google Scholar] [CrossRef]
- Sow, A.Y.; Dee, K.H.; Lee, S.W.; Eh Rak, A.A.L. An Assessment of Heavy Metals Toxicity in Asian Clam, Corbicula fluminea, from Mekong River, Pa Sak River, and Lopburi River, Thailand. Sci. World J. 2019, 2019, 1615298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, C.T.; Souza, M.M. The influence of lead on different proteins in gill cells from the freshwater bivalve, Corbicula fluminea, from defense to repair biomarkers. Arch. Environ. 2011, 62, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Fraysse, B.; Baudin, J.-P.; Garnier-Laplace, J.; Adam, C.; Boudou, A. Effects of Cd and Zn waterborne exposure on the uptake and depuration of 57Co, 110mAg and 134Cs by the Asiatic clam (Corbicula fluminea) and the zebra mussel (Dreissena polymorpha)—whole organism study. Environ. Pollut. 2002, 118, 297–306. [Google Scholar] [CrossRef]
- Zhang, H.; Reynolds, M. Cadmium exposure in living organisms: A short review. Sci. Total Environ. 2019, 678, 761–767. [Google Scholar] [CrossRef]
- Orisakwe, O.E. 11—Other heavy metals: Antimony, cadmium, chromium and mercury. In Toxicity of Building Materials, Woodhead Publishing Series in Civil and Structural Engineering; Pacheco-Torgal, F., Jalali, S., Fucic, A., Eds.; Woodhead Publishing: Cambridge, UK, 2012; pp. 297–333. [Google Scholar] [CrossRef]
- Loganathan, P.; Hedley, M.J.; Grace, N.D. Pasture Soils Contaminated with Fertilizer-Derived Cadmium and Fluorine: Livestock Effects. In Reviews of Environmental Contamination and Toxicology; Whitacre, D.M., Ed.; Springer: New York, NY, USA, 2008; pp. 29–66. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Ilahi, I. Environmental Chemistry and Ecotoxicology of Hazardous Heavy Metals: Environmental Persistence, Toxicity, and Bioaccumulation. J. Chem. 2019, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Vale, G.; Franco, C.; Diniz, M.S.; dos Santos, M.M.C.; Domingos, R.F. Bioavailability of cadmium and biochemical responses on the freshwater bivalve Corbicula fluminea—The role of TiO₂ nanoparticles. Ecotoxicol. Environ. Saf. 2014, 109, 161–168. [Google Scholar] [CrossRef]
- Ivar do Sul, J.A.; Costa, M.F. The present and future of microplastic pollution in the marine environment. Environ. Pollut. 2014, 185, 352–364. [Google Scholar] [CrossRef]
- Andrady, A.L. Microplastics in the marine environment. Mar. Pollut. Bull. 2011, 62, 1596–1605. [Google Scholar] [CrossRef]
- Moore, C.J. Synthetic polymers in the marine environment: A rapidly increasing, long-term threat. Environ. Res. 2008, 108, 131–139. [Google Scholar] [CrossRef]
- Guilhermino, L.; Vieira, L.R.; Ribeiro, D.; Tavares, A.S.; Cardoso, V.; Alves, A.; Almeida, J.M. Uptake and effects of the antimicrobial florfenicol, microplastics and their mixtures on freshwater exotic invasive bivalve Corbicula fluminea. Sci. Total Environ. 2018, 622–623, 1131–1142. [Google Scholar] [CrossRef]
- Oliveira, P.; Lopes-Lima, M.; Machado, J.; Guilhermino, L. Comparative sensitivity of European native (Anodonta anatina) and exotic (Corbicula fluminea) bivalves to mercury. Estuar. Coast. Shelf Sci. 2015, 167, 191–198. [Google Scholar] [CrossRef]
- Holmes, L.A.; Turner, A.; Thompson, R.C. Adsorption of trace metals to plastic resin pellets in the marine environment. Environ. Pollut. 2012, 160, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Rochman, C.M.; Hentschel, B.T.; Teh, S.J. Long-term sorption of metals is similar among plastic types: Implications for plastic debris in aquatic environments. PLoS ONE 2014, 9, e85433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashton, K.; Holmesk, L.; Turner, A. Association of metals with plastic production pellets in the marine environment. Mar. Pollut. Bull. 2010, 60, 2050–2055. [Google Scholar] [CrossRef]
- Eisler, R. Eisler’s Encyclopedia of Environmentally Hazardous Priority Chemicals, 1st ed.; Elsevier Science: Amsterdam, The Netherlands, 2007; p. 983. [Google Scholar]
- Wirnkor, V.A.; Ngozi, V.E.; Ebere, E.C. Microplastics, an emerging concern: A review of analytical techniques for detecting and quantifying microplatics. Anal. Methods Environ. Chem. J. 2019, 2, 15–32. [Google Scholar] [CrossRef]
- Minor, E.C.; Lin, R.; Burrows, A.; Cooney, E.M.; Grosshuesch, S.; Lafrancois, B. An analysis of microlitter and microplastics from Lake Superior beach sand and surface-water. Sci. Total Environ. 2020, 744, 140824. [Google Scholar] [CrossRef]
- Wagner, J.; Wang, Z.-M.; Ghosal, S.; Rochman, C.; Gassel, M.; Wall, S. Novel method for the extraction and identification of microplastics in ocean trawl and fish gut matrices. Anal. Methods 2017, 9, 1479–1490. [Google Scholar] [CrossRef]
- Wang, Z.M.; Wagner, J.; Ghosal, S.; Bedi, G.; Wall, S. SEM/EDS and optical microscopy analyses of microplastics in ocean trawl and fish guts. Sci. Total Environ. 2017, 603–604, 616–626. [Google Scholar] [CrossRef]
- Hummel, D.O. Atlas of Plastics Additives: Analysis by Spectrometric Methods; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar] [CrossRef]
- Escartín, E.; Porte, C. The use of cholinesterase and carboxylesterase activities from Mytilus galloprovincialis in pollution monitoring. Environ. Toxicol. Chem. 1997, 16, 2090–2095. [Google Scholar] [CrossRef]
- Galloway, T.S.; Millward, N.; Browne, M.A.; Depledge, M.H. Rapid assessment of organophosphorous/carbamate exposure in the bivalve mollusc Mytilus edulis using combined esterase activities as biomarkers. Aquat. Toxicol. 2002, 61, 169–180. [Google Scholar] [CrossRef]
- Deng, J.; Yu, L.; Liu, C.; Yu, K.; Shi, X.; Yeung, L.W.Y.; Lam, P.K.S.; Wu, R.S.S.; Zhou, B. Hexabromocyclododecane-induced developmental toxicity and apoptosis in zebrafish embryos. Aquat. Toxicol. 2009, 93, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Durak, I.; Yurtarslanl, Z.; Canbolat, O.; Akyol, Ö. A methodological approach to superoxide dismutase (SOD) activity assay based on inhibition of nitroblue tetrazolium (NBT) reduction. Clinic. Chim. Acta 1993, 214, 103–104. [Google Scholar] [CrossRef]
- Claiborne, A. Catalase activity. In CRC Handbook of Methods in Oxygen Radical Research; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 283–284. [Google Scholar]
- Habig, W.H.; Jakoby, W.B. Assays for differentiation of glutathione S-Transferases. In Methods in Enzymology, Detoxication and Drug Metabolism: Conjugation and Related Systems; Academic Press: Cambridge, MA, USA, 1981; pp. 398–405. [Google Scholar] [CrossRef]
- Gartaganis, S.P.; Patsoukis, N.E.; Nikolopoulos, D.K.; Georgiou, C.D. Evidence for oxidative stress in lens epithelial cells in pseudoexfoliation syndrome. Eye 2007, 21, 1406–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Fuentes, G.; Rubio-Escalante, F.J.; Noreña-Barroso, E.; Escalante-Herrera, K.S.; Schlenk, D. Impacts of oxidative stress on acetylcholinesterase transcription, and activity in embryos of zebrafish (Danio rerio) following Chlorpyrifos exposure. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2015, 172–173, 19–25. [Google Scholar] [CrossRef]
- Domingues, I.; Oliveira, R.; Lourenço, J.; Grisolia, C.K.; Mendo, S.; Soares, A.M.V.M. Biomarkers as a tool to assess effects of chromium (VI): Comparison of responses in zebrafish early life stages and adults. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 152, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Félix, L.M.; Vidal, A.M.; Serafim, C.; Valentim, A.M.; Antunes, L.M.; Monteiro, S.M.; Matos, M.; Coimbra, A.M. Ketamine induction of p53-dependent apoptosis and oxidative stress in zebrafish (Danio rerio) embryos. Chemosphere 2018, 201, 730–739. [Google Scholar] [CrossRef]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar] [CrossRef]
- Bertin, G.; Averbeck, D. Cadmium: Cellular effects, modifications of biomolecules, modulation of DNA repair and genotoxic consequences (a review). Biochimie 2006, 88, 1549–1559. [Google Scholar] [CrossRef]
- Liu, J.; Qian, S.Y.; Guo, Q.; Jiang, J.; Waalkes, M.P.; Mason, R.P.; Kadiiska, K.B. Cadmium generates reactive oxygen- and carbon-centered radical species in rats: Insights from in vivo spin-trapping studies. Free Radic. Biol. Med. 2008, 45, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Donaghy, L.; Hong, H.-K.; Jauzein, C.; Choi, K.-S. The known and unknown sources of reactive oxygen and nitrogen species in haemocytes of marine bivalve molluscs. Fish Shellfish Immunol. 2015, 42, 91–97. [Google Scholar] [CrossRef]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Livingstone, D.R. Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organisms. Mar. Pollut. Bull. 2001, 42, 656–666. [Google Scholar] [CrossRef]
- Vlahogianni, T.H.; Valavanidis, A. Heavy-metal effects on lipid peroxidation and antioxidant defence enzymes in mussels Mytilus galloprovinciali. J. Chem. Ecol. 2007, 23, 361–371. [Google Scholar] [CrossRef]
- Casalino, E.; Calzaretti, G.; Sblano, C.; Landriscina, C. Molecular inhibitory mechanisms of antioxidant enzymes in rat liver and kidney by cadmium. Toxicology 2002, 179, 37–50. [Google Scholar] [CrossRef]
- Ravikumar, A.; Arun, P.; Devi, K.V.; Augustine, J.; Kurup, P.A. Isoprenoid pathway and free radical generation and damage in neuropsychiatric disorders. Indian J. Exp. Biol. 2000, 38, 438–446. [Google Scholar] [PubMed]
- Ribeiro, F.; Garcia, A.R.; Pereira, B.P.; Fonseca, M.; Mestre, N.C.; Fonseca, T.G.; Ilharco, L.M.; Bebianno, M.J. Microplastics effects in Scrobicularia plana. Mar. Pollut. Bull. 2017, 122, 379–391. [Google Scholar] [CrossRef]
- Borković-Mitić, S.; Pavlović, S.; Perendija, B.; Despotović, S.; Gavrić, J.; Gačić, Z.; Saičić, Z. Influence of some metal concentrations on the activity of antioxidant enzymes and concentrations of vitamin E and SH-groups in the digestive gland and gills of the freshwater bivalve Unio tumidus from the Serbian part of Sava River. Ecol. Indic. 2013, 32, 212–221. [Google Scholar] [CrossRef]
- Martins, M.; Costa, P.M.; Ferreira, A.M.; Costa, M.H. Comparative DNA damage and oxidative effects of carcinogenic and non-carcinogenic sediment-bound PAHs in the gills of a bivalve. Aquat. Toxicol. 2013, 142–143, 85–95. [Google Scholar] [CrossRef]
- Fernández, B.; Campillo, J.A.; Martínez-Gómez, C.; Benedicto, J. Antioxidant responses in gills of mussel (Mytilus galloprovincialis) as biomarkers of environmental stress along the Spanish Mediterranean coast. Aquat. Toxicol. 2010, 99, 186–197. [Google Scholar] [CrossRef]
- Rocher, B.; Le Goff, J.; Peluhet, L.; Briand, M.; Manduzio, H.; Gallois, J.; Devier, M.H.; Geffard, O.; Gricourt, L.; Augagneur, S.; et al. Genotoxicant accumulation and cellular defence activation in bivalves chronically exposed to waterborne contaminants from the Seine River. Aquat. Toxicol. 2006, 79, 65–77. [Google Scholar] [CrossRef]
- Won, E.-J.; Kim, R.O.; Rhee, J.S.; Park, G.S.; Lee, J.; Shin, K.H.; Lee, Y.M.; Lee, J.S. Response of glutathione S-transferase (GST) genes to cadmium exposure in the marine pollution indicator worm, Perinereis nuntia. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 154, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Selvam, S.; Jesuraja, K.; Venkatramanan, S.; Roy, P.D.; Jeyanthi Kumari, V. Hazardous microplastic characteristics and its role as a vector of heavy metal in groundwater and surface water of coastal south India. J. Hazard. Mater. 2021, 402, 123786. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, D.; Almeida, A.; Calisto, V.; Esteves, V.I.; Schneider, R.J.; Wrona, F.J.; Soares, A.M.V.M.; Figueira, E.; Freitas, R. Caffeine impacts in the clam Ruditapes philippinarum: Alterations on energy reserves, metabolic activity and oxidative stress biomarkers. Chemosphere 2016, 160, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Coppola, F.; Almeida, A.; Henriques, B.; Soares, A.M.V.M.; Figueira, E.; Pereira, E.; Freitas, R. Biochemical impacts of Hg in Mytilus galloprovincialis under present and predicted warming scenarios. Sci. Total Environ. 2017, 601–602, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Almeida, E.A.; Miyamoto, S.; Bainy, A.C.D.; de Medeiros, M.H.G.; Di Mascio, P. Protective effect of phospholipid hydroperoxide glutathione peroxidase (PHGPx) against lipid peroxidation in mussels Perna perna exposed to different metals. Mar. Pollut. Bull. 2004, 49, 386–392. [Google Scholar] [CrossRef]
- O’Donovan, S.; Mestre, N.C.; Abel, S.; Fonseca, T.G.; Carteny, C.C.; Cormier, B.; Keiter, S.H.; Bebianno, M.J. Ecotoxicological Effects of Chemical Contaminants Adsorbed to Microplastics in the Clam Scrobicularia plana. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Cheung, C.C.C.; Siu, W.H.L.; Richardson, B.J.; De Luca-Abbott, S.B.; Lam, P.K.S. Antioxidant responses to benzo[a]pyrene and Aroclor 1254 exposure in the green-lipped mussel, Perna viridis. Environ. Pollut. 2004, 128, 393–403. [Google Scholar] [CrossRef]
- Karthikeyan, J.; Bavani, G. Effect of cadmium on lactate dehyrogenase isoenzyme, succinate dehydrogenase and Na(+)-K(+)-ATPase in liver tissue of rat. J. Environ. Biol. 2009, 30, 895–898. [Google Scholar]
- Kumar, N.; Krishnani, K.K.; Singh, N.P. Oxidative and cellular stress as bioindicators for metal contamination in freshwater mollusk Lamellidens marginalis. Environ. Sci. Pollut. Res. 2017, 24, 16137–16147. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Zhang, N.; Jin, S.-R.; Chen, Z.-Z.; Gao, J.-Z.; Liu, Y.; Liu, H.-P.; Xu, Z. Microplastics have a more profound impact than elevated temperatures on the predatory performance, digestion and energy metabolism of an Amazonian cichlid. Aquat. Toxicol. 2018, 195, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Lionetto, M.G.; Caricato, R.; Calisi, A.; Giordano, M.E.; Schettino, T. Acetylcholinesterase as a biomarker in environmental and occupational medicine: New insights and future perspectives. Biomed. Res. Int. 2013, 321213. [Google Scholar] [CrossRef] [PubMed]
- Matozzo, V.; Fabrello, J.; Masiero, L.; Ferraccioli, F.; Finos, L.; Pastore, P.; Di Gangi, I.M.; Bogialli, S. Ecotoxicological risk assessment for the herbicide glyphosate to non-target aquatic species: A case study with the mussel Mytilus galloprovincialis. Environ. Pollut. 2018, 233, 623–632. [Google Scholar] [CrossRef]
- Sun, W.; Chen, L.; Zheng, W.; Wei, X.; Wu, W.; Duysen, E.G.; Jiang, W. Study of acetylcholinesterase activity and apoptosis in SH-SY5Y cells and mice exposed to ethanol. Toxicology 2017, 384, 33–39. [Google Scholar] [CrossRef]
- Zhang, X.J.; Yang, L.; Zhao, Q.; Caen, J.P.; He, H.Y.; Jin, Q.H.; Guo, L.H.; Alemany, M.; Zhang, L.Y.; Shi, Y.F. Induction of acetylcholinesterase expression during apoptosis in various cell types. Cell Death Differ. 2002, 9, 790–800. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Cd Levels | Control | Cd | MPs | Cd + MPs |
|---|---|---|---|---|
| Water (µg L−1) | 0.35 ± 0.01 | 6.73 ± 0.10 | 0.29 ± 0.10 | 4.94 ± 0.24 |
| Clam (µg Cd g−1 LW) | 0.51 ± 0.20 | 1.35 ± 0.12 | 0.70 ± 0.05 | 1.86 ± 0.37 |
| Biochemical Parameters | Analyzed Organs | Control | Cd | MPs | Cd + MPs |
|---|---|---|---|---|---|
| ROS µmol DCF/mg of protein | Gill | 20.06 ± 3.7 a | 31.25 ± 4.5 b | 18.65 ± 1.7 a | 29.76 ± 3.8 b |
| Digestive gland | 49.2 ± 5.2 a | 103.0 ± 9.8 b | 51.8 ± 4.8 a | 62.8 ± 9.6 c | |
| Gonad | 72.1 ± 4.7 a | 54.0 ± 4.0 b | 41.5 ± 6.5 c | 53.3 ± 6.2 b | |
| SOD U/mg of protein | Gill | 661.3 ± 75.5 a | 3.8 ± 0.8 b | 351.6 ± 12.5 c | 5.7 ± 1.3 b |
| Digestive gland | 483.0 ± 19.7 a | 6.9 ± 1.0 b | 465.7 ± 28.6 a | 13.1 ± 2.3 b | |
| Gonad | 2040.4 ± 194.8 a | 5.4 ± 1.3 b | 406.7 ± 25.8 c | 3.4 ± 0.4 b | |
| CAT U/mg of protein | Gill | 65.5 ± 9.3 a | 72.8 ± 7.4 a | 91.6 ± 10.4 b | 84.0 ± 6.6 b |
| Digestive gland | 71.2 ± 9.1 a | 237.2 ± 21.3 b | 91.0 ± 6.5 c | 111.3 ± 3.8 d | |
| Gonad | 315.9 ± 31.6 a | 309.9 ± 35.0 a | 91.4 ± 8.4 b | 544.4 ± 67.3 c | |
| GST nmol CDNB/min.mg of protein | Gill | 10.6 ± 1.6 a | 22.7 ± 2.0 b | 11.0 ± 4.6 a | 39.7 ± 3.5 c |
| Digestive gland | 25.5 ± 3.1 a | 173.0 ± 14.7 b | 33.5 ± 1.5 a | 220.6 ± 10.9 c | |
| Gonad | 44.0 ± 10.2 a | 159.3 ± 19.8 b | 29.0 ± 2.8 a | 135.9 ± 11.2 c | |
| Reduced/Oxidized Glutathione Ratio | Gill | 0.8 ± 0.03 a | 0.1 ± 0.01 b | 1.2 ± 0.1 c | 0.1 ± 0.02 b |
| Digestive gland | 0.6 ± 0.02 a | 0.1 ± 0.01 b | 0.6 ± 0.08 a | 0.1 ± 0.01 b | |
| Gonad | 1.1 ± 0.1 a | 0.1 ± 0.02 b | 1.6 ± 0.2 c | 0.08 ± 0.03 b | |
| LPO nmol MDA/mg protein | Gill | 0.2 ± 0.03 a | 0.7 ± 0.08 b | 0.4 ± 0.05 c | 1.0 ± 0.1 d |
| Digestive gland | 0.3 ± 0.03 a | 1.2 ± 0.1 b | 0.4 ± 0.1 a | 0.8 ± 0.03 c | |
| Gonad | 0.5 ± 0.05 a | 0.7 ± 0.04 b | 0.3 ± 0.03 c | 0.3 ± 0.01 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parra, S.; Varandas, S.; Santos, D.; Félix, L.; Fernandes, L.; Cabecinha, E.; Gago, J.; Monteiro, S.M. Multi-Biomarker Responses of Asian Clam Corbicula fluminea (Bivalvia, Corbiculidea) to Cadmium and Microplastics Pollutants. Water 2021, 13, 394. https://doi.org/10.3390/w13040394
Parra S, Varandas S, Santos D, Félix L, Fernandes L, Cabecinha E, Gago J, Monteiro SM. Multi-Biomarker Responses of Asian Clam Corbicula fluminea (Bivalvia, Corbiculidea) to Cadmium and Microplastics Pollutants. Water. 2021; 13(4):394. https://doi.org/10.3390/w13040394
Chicago/Turabian StyleParra, Susana, Simone Varandas, Dércia Santos, Luís Félix, Lisete Fernandes, Edna Cabecinha, Jesús Gago, and Sandra Mariza Monteiro. 2021. "Multi-Biomarker Responses of Asian Clam Corbicula fluminea (Bivalvia, Corbiculidea) to Cadmium and Microplastics Pollutants" Water 13, no. 4: 394. https://doi.org/10.3390/w13040394
APA StyleParra, S., Varandas, S., Santos, D., Félix, L., Fernandes, L., Cabecinha, E., Gago, J., & Monteiro, S. M. (2021). Multi-Biomarker Responses of Asian Clam Corbicula fluminea (Bivalvia, Corbiculidea) to Cadmium and Microplastics Pollutants. Water, 13(4), 394. https://doi.org/10.3390/w13040394