
Dynamic of Dominant Plant Communities in Kettle Holes (Northeast Germany) during a Five-Year Period of Extreme Weather Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. UAS Data Acquisition and Image Processing
2.3. Vegetation Data, Kettle Hole and Water Area
2.4. Meteorological Data
2.5. Statistical Analysis
3. Results
3.1. Meteorological and Hydrological Conditions
3.2. Effect of Dry and Wet Periods on Dominant Plant Communities
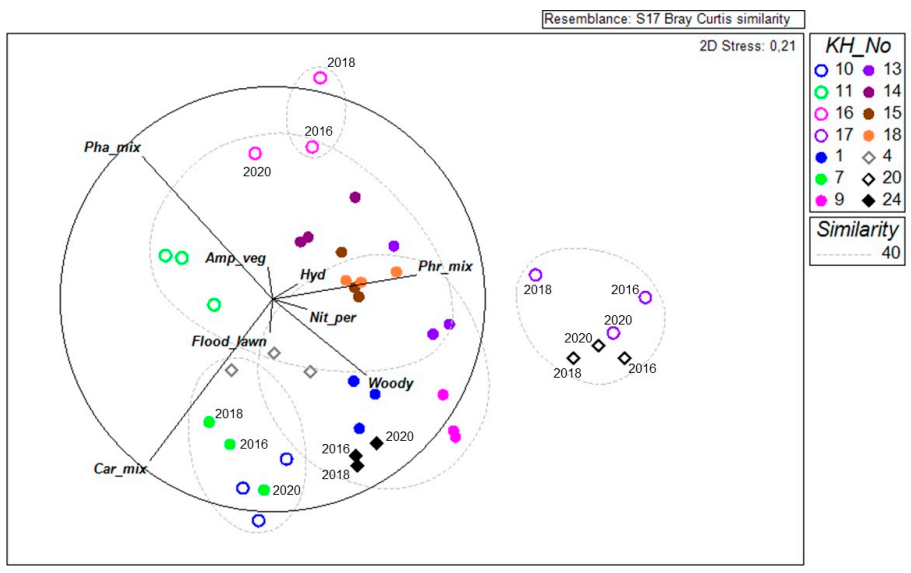
3.3. Interannual Variability in Dominant Plant Community Composition
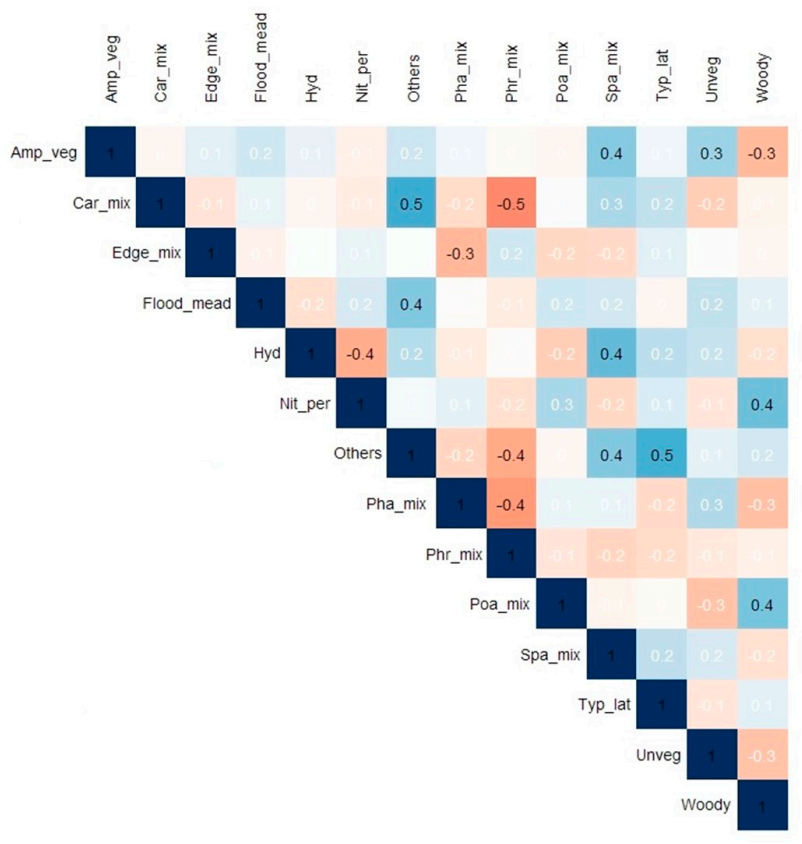
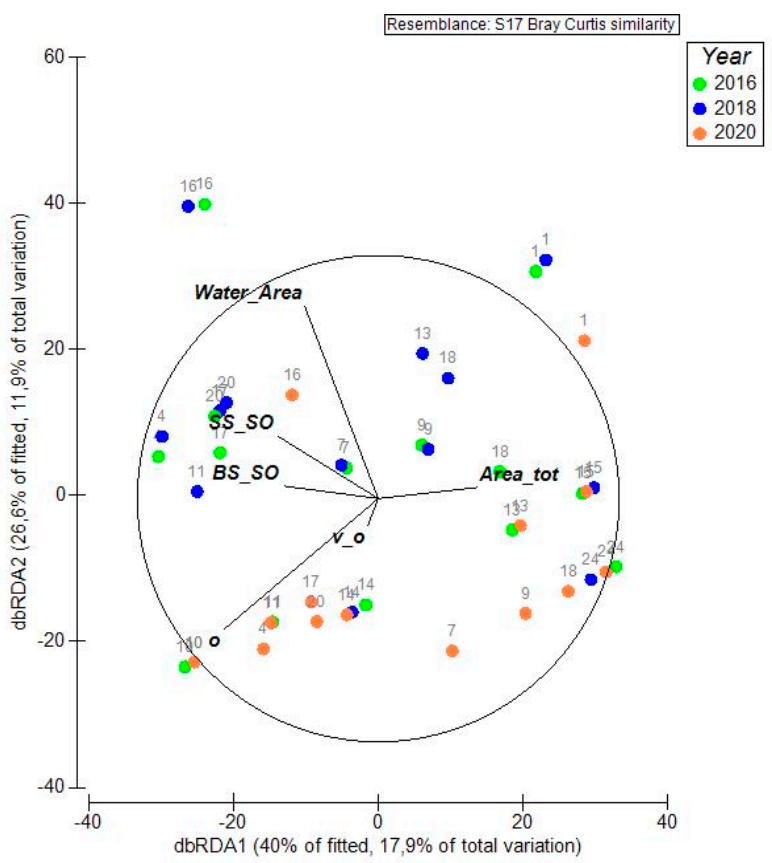
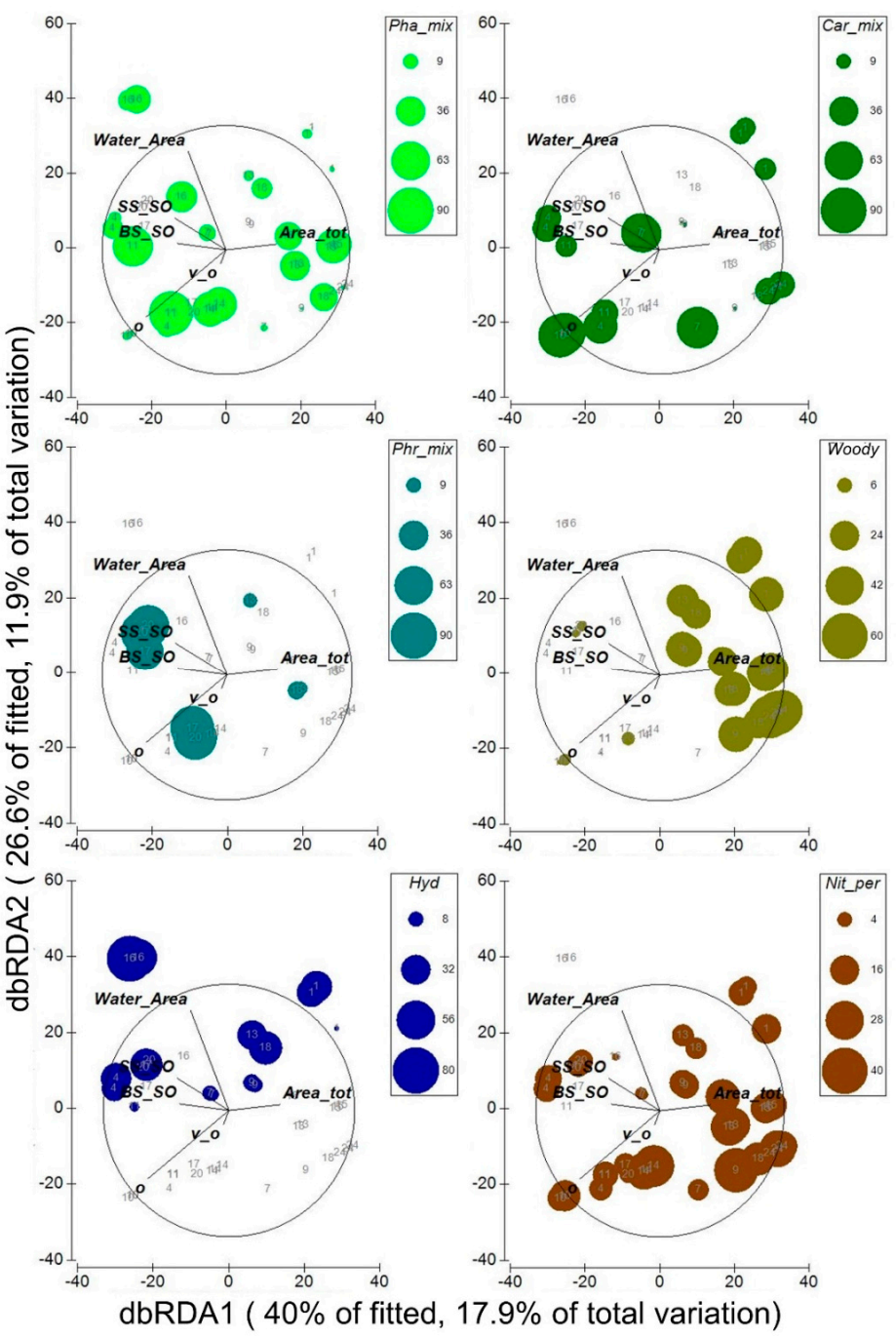
3.4. Relation between Dominant Plant Communities, Interannual Scale and Environmental Variables
4. Discussion
4.1. Different Groups of Dominant Plant Communities Are Differently Affected by Dry and Wet Periods
4.2. Interannual Variability of Kettle Hole Vegetation Increases during Extreme Weather Conditions
4.3. Future Perspectives for Kettle Hole Occurrence in Central European Pleistocene Landscapes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeffries, M.J. Flood, drought and the inter-annual variation to the number and size of ponds and small wetlands in an English lowland landscape over three years of weather extremes. Hydrobiologia 2016, 768, 255–272. [Google Scholar] [CrossRef] [Green Version]
- Thornhill, I.A.; Biggs, J.; Hill, M.J.; Briers, R.; Gledhill, D.; Wood, P.J.; Gee, J.H.R.; Ledger, M.; Hassall, C. The functional response and resilience in small waterbodies along land-use and environmental gradients. Glob. Chang. Biol. 2018, 24, 3079–3092. [Google Scholar] [CrossRef]
- Marlene, P.; Kalettka, T.; Onandia, G.; Balla, D.; Lischeid, G.; Pätzig, M.; Thomas, K.; Gabriela, O.; Dagmar, B.; Gunnar, L. How much information do we gain from multiple-year sampling in natural pond research? Limnologica 2020, 80, 125728. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Riley, W.D.; Potter, E.C.; Biggs, J.; Collins, A.L.; Jarvie, H.P.; Jones, J.I.; Kelly-Quinn, M.; Ormerod, S.J.; Sear, D.A.; Wilby, R.L.; et al. Small Water Bodies in Great Britain and Ireland: Ecosystem function, human-generated degradation, and options for restorative action. Sci. Total. Environ. 2018, 645, 1598–1616. [Google Scholar] [CrossRef]
- Winter, T.C.; Rosenberry, D.O. Hydrology of Prairie Pothole Wetlands during Drought and Deluge: A 17-Year Study of the Cottonwood Lake Wetland Complex in North Dakota in the Perspective of Longer Term Measured and Proxy Hydrological Records. Clim. Chang. 1998, 40, 189–209. [Google Scholar] [CrossRef]
- Valk, A.G.V.D. Water-level fluctuations in North American prairie wetlands. Hydrobiologia 2005, 539, 171–188. [Google Scholar] [CrossRef]
- Hayashi, M.; Van Der Kamp, G.; Rosenberry, D.O. Hydrology of Prairie Wetlands: Understanding the Integrated Surface-Water and Groundwater Processes. Wetlands 2016, 36, 237–254. [Google Scholar] [CrossRef]
- Van Der Valk, A.G.; Davis, C.B. The Role of Seed Banks in the Vegetation Dynamics of Prairie Glacial Marshes. Ecology 1978, 59, 322–335. [Google Scholar] [CrossRef]
- Davies, B.; Biggs, J.; Williams, P.; Whitfield, M.; Nicolet, P.; Sear, D.; Bray, S.; Maund, S. Comparative biodiversity of aquatic habitats in the European agricultural landscape. Agric. Ecosyst. Environ. 2008, 125, 1–8. [Google Scholar] [CrossRef]
- Onandia, G.; Lischeid, G.; Kalettka, T.; Kleeberg, A.; Omari, M.; Premke, K.; Arhonditsis, G.B. Biogeochemistry of natural ponds in agricultural landscape: Lessons learned from modeling a kettle hole in Northeast Germany. Sci. Total. Environ. 2018, 634, 1615–1630. [Google Scholar] [CrossRef]
- Kazanjian, G.; Flury, S.; Attermeyer, K.; Kalettka, T.; Kleeberg, A.; Premke, K.; Köhler, J.; Hilt, S. Primary production in nutrient-rich kettle holes and consequences for nutrient and carbon cycling. Hydrobiologia 2017, 806, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Johnson, W.C.; Millett, B.V.; Gilmanov, T.; Voldseth, R.A.; Guntenspergen, G.R.; Naugle, D.E. Vulnerability of north-ern prairie wetlands to climate change. Bioscience 2005, 55, 863–872. [Google Scholar] [CrossRef]
- Johnson, W.C.; Werner, B.; Guntenspergen, G.R. Non-linear responses of glaciated prairie wetlands to climate warming. Clim. Chang. 2015, 134, 209–223. [Google Scholar] [CrossRef]
- Mushet, D.M.; McKenna, O.P.; McLean, K.I. Alternative stable states in inherently unstable systems. Ecol. Evol. 2019, 10, 843–850. [Google Scholar] [CrossRef]
- Johnson, W.C.; Boettcher, S.E.; Poiani, K.A.; Guntenspergen, G.R. Influence of weather extremes on the water levels of glaciated prairie wetlands. Wetlands 2004, 24, 385–398. [Google Scholar] [CrossRef]
- Bauder, E.T. Inundation effects on small-scale plant distributions in San Diego, California vernal pools. Aquat. Ecol. 2000, 34, 43–61. [Google Scholar] [CrossRef]
- Altenfelder, S.; Raabe, U.; Albrecht, H. Effects of water regime and agricultural land use on diversity and species com-position of vascular plants inhabiting temporary ponds in northeastern Germany. Tuexenia 2014, 34, 145–162. [Google Scholar] [CrossRef]
- Rocarpin, P.; Gachet, S.; Metzner, K.; Saatkamp, A. Moisture and soil parameters drive plant community assembly in Mediterranean temporary pools. Hydrobiol. 2015, 781, 55–66. [Google Scholar] [CrossRef]
- Stewart, R.E.; Kantrud, H.A. Classification of Natural Ponds and Lakes in the Glaciated Prairie Region; Bureau of Sport Fisheries and Wildlife, U.S. Fish and Wildlife Service: Washington, DC, USA, 1971.
- Kantrud, H.A.; Krapu, G.L.; Swanson, G.A. Prairie basin wetlands of the Dakotas—A community profile. Biol. Rep. 1989, 85, 1–111. [Google Scholar]
- Van der Valk, A. Succession in Wetlands: A Gleasonian Approach. Ecology 1981, 62, 688–696. [Google Scholar] [CrossRef] [Green Version]
- Seabloom, E.W.; Moloney, K.A.; van der Valk, A.G. Constraints on the establishment of plants along a fluctuating water-depth gradient. Ecology 2001, 82, 2216–2232. [Google Scholar] [CrossRef]
- Euliss, N.H.; LaBaugh, J.W.; Fredrickson, L.H.; Mushet, D.M.; Laubhan, M.K.; Swanson, G.A.; Winter, T.C.; Rosenberry, D.O.; Nelson, R.D. The wetland continuum: A conceptual framework for interpreting biological studies. Wetlands 2004, 24, 448–458. [Google Scholar] [CrossRef]
- Mushet, D.M.; McKenna, O.P.; La Baugh, J.W.; Euliss, N.H.; Rosenberry, D.O. Accommodating State Shifts within the Conceptual Framework of the Wetland Continuum. Wetlands 2018, 38, 647–651. [Google Scholar] [CrossRef]
- Sayer, C.D.; Andrews, K.; Shilland, E.; Edmonds, N.; Edmonds-Brown, V.; Patmore, I.R.; Emson, D.; Axmacher, J.C. The role of pond management for biodiversity conservation in an agricultural landscape. Aquat. Conserv. Mar. Freshw. Ecosyst. 2012, 22, 626–638. [Google Scholar] [CrossRef]
- Lewis-Phillips, J.; Brooks, S.J.; Sayer, C.D.; Patmore, I.R.; Hilton, G.M.; Harrison, A.; Robson, H.; Axmacher, J.C. Ponds as insect chimneys: Restoring overgrown farmland ponds benefits birds through elevated productivity of emerging aquatic insects. Biol. Conserv. 2020, 241, 108253. [Google Scholar] [CrossRef]
- Lewis-Phillips, J.; Brooks, S.; Sayer, C.D.; McCrea, R.; Siriwardena, G.; Axmacher, J.C. Pond management enhances the local abundance and species richness of farmland bird communities. Agric. Ecosyst. Environ. 2019, 273, 130–140. [Google Scholar] [CrossRef]
- Johnson, W.C.; Poiani, K.A. Climate Change Effects on Prairie Pothole Wetlands: Findings from a Twenty-five Year Numerical Modeling Project. Wetlands 2016, 36, 273–285. [Google Scholar] [CrossRef]
- McKenna, O.P.; Mushet, D.M.; Rosenberry, D.O.; LaBaugh, J.W. Evidence for a climate-induced ecohydrological state shift in wetland ecosystems of the southern Prairie Pothole Region. Clim. Chang. 2017, 145, 273–287. [Google Scholar] [CrossRef]
- Van Der Valk, A.G.; Volin, J.C.; Wetzel, P.R. Predicted Changes in Interannual Water-Level Fluctuations Due to Climate Change and Its Implications for the Vegetation of the Florida Everglades. Environ. Manag. 2015, 55, 799–806. [Google Scholar] [CrossRef]
- Van Der Valk, A.; Mushet, D.M. Interannual Water-level Fluctuations and the Vegetation of Prairie Potholes: Potential Impacts of Climate Change. Wetlands 2016, 36, 397–406. [Google Scholar] [CrossRef]
- Vasić, F.; Paul, C.; Strauss, V.; Helming, K. Ecosystem Services of Kettle Holes in Agricultural Landscapes. Agronomy 2020, 10, 1326. [Google Scholar] [CrossRef]
- Lehsten, D.; Von Asmuth, J.R.; Kleyer, M. Simulation of Water Level Fluctuations in Kettle Holes Using a Time Series Model. Wetlands 2011, 31, 511–520. [Google Scholar] [CrossRef]
- Pätzig, M.; Kalettka, T.; Glemnitz, M.; Berger, G. What governs macrophyte species richness in kettle hole types? A case study from Northeast Germany. Limnologica 2012, 42, 340–354. [Google Scholar] [CrossRef]
- Raulings, E.J.; Morris, K.; Roache, M.C.; Boon, P.I. The importance of water regimes operating at small spatial scales for the diversity and structure of wetland vegetation. Freshw. Biol. 2010, 55, 701–715. [Google Scholar] [CrossRef]
- Kalettka, T.; Rudat, C. Hydrogeomorphic types of glacially created kettle holes in North-East Germany. Limnologica 2006, 36, 54–64. [Google Scholar] [CrossRef] [Green Version]
- Wich, S.A.; Koh, L.P. Conservation Drones: Mapping and Monitoring Biodiversity; Oxford University Press: New York, NY, USA, 2018. [Google Scholar]
- Díaz-Delgado, R.; Mücher, S. Editorial of Special Issue “Drones for Biodiversity Conservation and Ecological Monitoring”. Drones 2019, 3, 47. [Google Scholar] [CrossRef] [Green Version]
- Chabot, D.; Bird, D.M. Small unmanned aircraft: Precise and convenient new tools for surveying wetlands. J. Unmanned Veh. Syst. 2013, 1, 15–24. [Google Scholar] [CrossRef]
- Husson, E.; Hagner, O.; Ecke, F. Unmanned aircraft systems help to map aquatic vegetation. Appl. Veg. Sci. 2014, 17, 567–577. [Google Scholar] [CrossRef]
- Verch, G. Weather Data 2015, Dedelow, Germany; Leibniz Centre for Agricultural Landscape Research (ZALF): Müncheberg, Germany, 2018. [Google Scholar] [CrossRef]
- Verch, G. Weather Data 2016, Dedelow, Germany; Leibniz Centre for Agricultural Landscape Research (ZALF): Müncheberg, Germany, 2019. [Google Scholar] [CrossRef]
- Verch, G. Weather Data 2017, Dedelow, Germany; Leibniz Centre for Agricultural Landscape Research (ZALF): Müncheberg, Germany, 2020. [Google Scholar] [CrossRef]
- Verch, G. Weather Data 2018, Dedelow, Germany; Leibniz Centre for Agricultural Landscape Research (ZALF): Müncheberg, Germany, 2021. [Google Scholar] [CrossRef]
- Verch, G. Weather Data 2019, Dedelow, Germany; Leibniz Centre for Agricultural Landscape Research (ZALF): Müncheberg, Germany, 2021. [Google Scholar] [CrossRef]
- Verch, G. Weather Data 2020, Dedelow, Germany; Leibniz Centre for Agricultural Landscape Research (ZALF): Müncheberg, Germany, 2021. [Google Scholar] [CrossRef]
- Richter, D. Ergebnisse Methodischer Untersuchungen zu Korrektur des Systematischen Meßfehlers des Hellmann-Niederschlagsmessers (Berichte des Deutschen Wetterdienstes); Selbstverlag des Deutschen Wetterdienstes: Offenbach, Germany, 1995. [Google Scholar]
- Wendling, U.; Müller, J.; Schwede, K. Ergebnisse von Verdunstungsmessungen über Gras mit einem offline-Datenerfassungssystem. Z. Meteorol. 1984, 34, 190–202. [Google Scholar]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The Meta-Analisys of Response Ratios in Experimental Ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Revelle, W. psych: Procedures for Psychological, Psychometric, and Personality Research; R Package Version 2.0.12; Northwestern University: Evanston, IL, USA, 2020; Available online: https://CRAN.R-project.org/package=psych (accessed on 20 December 2020).
- Hallett, L.; Avolio, M.; Carroll, I.T.; Jones, S.K.; MacDonald, A.A.; Flynn, D.F.B.; Slaughter, P.; Ripplinger, J.; Collins, S.L.; Gries, C.; et al. codyn: Community Dynamics Metrics. R Package Version 2.0.5. 2020. Available online: https://github.com/NCEAS/codyn (accessed on 18 December 2020). [CrossRef]
- Anderson, K.J. Temporal Patterns in Rates of Community Change during Succession. Am. Nat. 2007, 169, 780–793. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statisitcal Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Akaike, H. Information theory as an extension of the maximum likelihood principle. In Proceedings of the Second International Symposium on Information Theory, Tsahkadsor, Armenia, 2–8 September 1973; pp. 267–281. [Google Scholar]
- Erwin, K.L. Wetlands and global climate change: The role of wetland restoration in a changing world. Wetl. Ecol. Manag. 2009, 17, 71–84. [Google Scholar] [CrossRef]
- Frey, W.; Lösch, R. Lehrbuch der Geobotanik—Pflanze und Vegetation in Raum und Zeit, 2nd ed.; Spektrum Akademischer Verlag: München, Germany, 2004. [Google Scholar]
- Chapin, F.S.; Matson, P.A.; Vitousek, P.M. Principles of Terrestrial Ecosystem Ecology, 2nd ed.; Springer Science + Business Media: Berlin/Heidelberg, Germnay, 2011. [Google Scholar]
- Jeffries, M. The spatial and temporal heterogeneity of macrophyte communities in thirty small, temporary ponds over a period of ten years. Ecography 2008, 31, 765–775. [Google Scholar] [CrossRef]
- Rhazi, L.; Grillas, P.; Rhazi, M.; Aznar, J.-C. Ten-year dynamics of vegetation in a Mediterranean temporary pool in western Morocco. Hydrobiologia 2009, 634, 185–194. [Google Scholar] [CrossRef]
- Francová, K.; Šumberová, K.; Kučerová, A.; Šorf, M.; Grill, S.; Exler, N.; Vrba, J. Drivers of plant species composition of ecotonal vegetation in two fishpond management types. Wetl. Ecol. Manag. 2021, 29, 93–110. [Google Scholar] [CrossRef]
- Galatowitsch, S.M.; Anderson, N.O.; Ascher, P.D. Invasiveness in wetland plants in temperate North America. Wetlands 1999, 19, 733–755. [Google Scholar] [CrossRef]
- Mulhouse, J.M.; De Steven, D.; Lide, R.F.; Sharitz, R.R. Effects of dominant species on vegetation change in Carolina bay wetlands following a multi-year drought1. J. Torrey Bot. Soc. 2005, 132, 411–420. [Google Scholar] [CrossRef]
- Wilcox, D.A. Implications of hydrologic variability on the succession of plants in Great Lakes wetlands. Aquat. Ecosyst. Health Manag. 2004, 7, 223–231. [Google Scholar] [CrossRef]
- Oertli, B.; Joye, D.A.; Castella, E.; Juge, R.; Cambin, D.G.; Lachavanne, J.-B. Does size matter? The relationship between pond area and biodiversity. Biol. Conserv. 2002, 104, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Van Leeuwen, C.H.A.; Sarneel, J.M.; Van Paassen, J.; Rip, W.J.; Bakker, E.S. Hydrology, shore morphology and species traits affect seed dispersal, germination and community assembly in shoreline plant communities. J. Ecol. 2014, 102, 998–1007. [Google Scholar] [CrossRef] [Green Version]
- Buth, M.; Kahlenborn, W.; Savelsberg, J.; Becker, N.; Bubeck, P.; Kabisch, S.; Kofler, C. Germany’s vulnerability to Climate Change—Summary. Dessau-Roßlau 2015, 11, 2015. [Google Scholar]
- Shin, H.R.; Kneitel, J.M. Warming interacts with inundation timing to influence the species composition of California vernal pool communities. Hydrobiologica 2019, 843, 93–105. [Google Scholar] [CrossRef]
- Bourgeois, B.; Vanasse, A.; González, E.; Andersen, R.; Poulin, M. Threshold dynamics in plant succession after tree planting in agricultural riparian zones. J. Appl. Ecol. 2016, 53, 1704–1713. [Google Scholar] [CrossRef]
- Rheinhardt, R.D.; Faser, K. Relationship between hydrology and zonation of freshwater swale wetlands on lower hatteras island, North Carolina, USA. Wetlands 2001, 21, 265–273. [Google Scholar] [CrossRef]
- Frielinghaus, M.; Vahrson, W.-G. Soil translocation by water erosion from agricultural cropland into wet depressions (morainic kettle holes). Soil Tillage Res. 1998, 46, 23–30. [Google Scholar] [CrossRef]
- Kneitel, J.M.; Lessin, C.L. Ecosystem-phase interactions: Aquatic eutrophication decreases terrestrial plant diversity in California vernal pools. Oecologia 2009, 163, 461–469. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KH_No | VegType 2016 | HGM | KA | KA_C | SS_C | LP_2011–2017 | LSOT_2011–2017 |
|---|---|---|---|---|---|---|---|
| 1 | Edge type with shore woods and sedges | SS-S | 2271 ± 89 | ≤0.32 | ≤40 | 5_permanent | 1_no overflow |
| 4 | Edge type with sedges | BS-SO | 3224 ± 143 | ≤1.0 | ≤20 | 4_semi-permanent | 3_fully overflowing |
| 7 | Full reed type with sedges | SS-S | 1878 ±32 | ≤0.32 | ≤30 | 3_periodic | 1_no overflow |
| 9 | Full reed type with reeds and shore woods | SS-S | 2910 ± 70 | ≤0.32 | ≤30 | 4_semi-permanent | 1_no overflow |
| 10 | Full reed type with sedges | SS-SO | 789 ± 25 | ≤0.10 | ≤20 | 2_episodic | 2_partly overflowing |
| 11 | Full reed type with reed canary grass | SS-SO | 1286 ± 8 | ≤0.32 | ≤20 | 3_periodic | 3_fully overflowing |
| 13 | Edge type with reed canary grass | SS-S | 1017 ± 27 | ≤0.10 | ≤40 | 3_periodic | 1_no overflow |
| 14 | Full reed type with reed canary grass | SS-S | 379 ± 23 | ≤0.10 | ≤40 | 2_episodic | 1_no overflow |
| 15 | Full reed type with shore woods and reed canary grass | SS-S | 1560 ± 55 | ≤0.32 | ≤40 | 2_episodic | 1_no overflow |
| 16 | Edge type with reed canary grass | SS-SO | 1014 ± 52 | ≤0.10 | ≤30 | 4_semi-permanent | 3_fully overflowing |
| 17 | Full reed type with reed | SS-SO | 1660 ± 93 | ≤0.32 | ≤20 | 4_semi-permanent | 2_partly overflowing |
| 18 | Full reed type with shore woods and reed canary grass | SS-S | 3852 ± 17 | ≤1.0 | ≤30 | 4_semi-permanent | 1_no overflow |
| 20 | Full reed type with reed | BS-SO | 4644 ± 356 | ≤1.0 | ≤20 | 3_periodic | 2_partly overflowing |
| 24 | Full reed type with shore woods and sedges | BS-S | 4834 ± 369 | ≤1.0 | ≤30 | 3_periodic | 1_no overflow |
| KH_No | pH | EC | Eh | O2 | SRP | NH4-N | TN | TP | CH |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 7.6 ± 0.6 (4) | 286 ± 36 (4) | 426 ± 56 (3) | 10.7 ± 3.3 (4) | 0.026 ± 0.030 (4) | 0.212 ± 0.277 (4) | 1.51 ± 0.40 (4) | 0.09 ± 0.03 (4) | 9.7 ± 1.7 (3) |
| 4 | 8.0 ± 0.4 (4) | 452 ± 50 (4) | 431 ± 24 (3) | 10.9 ± 2.4 (4) | 0.076 ± 0.123 (4) | 0.054 ± 0.029 (4) | 2.73 ± 0.62 (4) | 0.18 ± 0.18 (4) | 13.3 ± 8.5 (3) |
| 7 | 7.4 ± 0.3 (5) | 611 ± 253 (5) | 347 ± 195 (4) | 14.3 ± 9.1 (5) | 0.075 ± 0.115 (5) | 0.072 ± 0.070 (5) | 4.15 ± 1.97 (5) | 0.23 ± 0.18 (5) | 9.0 ± 2.9 (4) |
| 9 | 6.5 ± 1.2 (4) | 744 ± 564 (4) | 340 ± 160 (3) | 13.2 ± 7.6 (4) | 0.521 ± 0.504 (4) | 1.671 ± 1.557 (4) | 4.55 ± 2.90 (4) | 0.77 ± 0.46 (4) | 5.5 ± 1.7 |
| 10 | 7.0 ± 0.5 (4) | 433 ± 199 (4) | 414 ± 19 (3) | 12.2 ± 2.7 (4) | 0.078 ± 0.046 (4) | 0.122 ± 0.095 (4) | 2.50 ± 1.35 (4) | 0.17 ± 0.10 (4) | 7.5 ± 4.4 (4) |
| 11 | 7.2 (1) | 459 (1) | 203 (1) | 9.2 (1) | Na | Na | Na | Na | 10.0 (1) |
| 13 | 7.5 (1) | 871 (1) | 259 (1) | 3.7 (1) | 0.068 (1) | 0.126 (1) | 2.65 (1) | 0.30 (1) | 21.0 (1) |
| 14 | 6.7 (1) | 155 (1) | 379 (1) | 13.7 (1) | 0.264 (1) | 0.204 (1) | 2.76 (1) | 0.42 (1) | 4.0 (1) |
| 15 | 6.7 (1) | 175 (1) | 456 (1) | 13.2 (1) | 0.455 (1) | 0.128 (1) | 1.99 (1) | 0.60 (1) | 5.0 (1) |
| 16 | 8.5 ± 0.44 (5) | 653 ± 166 (5) | 395 ± 50 (4) | 14.0 ± 5.3 (5) | 0.050 ± 0.049 (5) | 0.055 ± 0.042 (5) | 3.33 ± 1.86 (5) | 0.11 ± 0.08 (5) | 10 ± 2.8 (4) |
| 17 | 7.6 ± 0.6 (5) | 686 ± 124 (5) | 450 ± 31 (4) | 13.6 ± 3.1 (5) | 0.027 ± 0.030 (5) | 0.097 ± 0.065 (5) | 4.28 ± 2.66 (5) | 0.15 ± 0.10 (5) | 11.5 ± 3.3 (4) |
| 18 | 7.5 ± 0.7 (5) | 676 ± 248 (5) | 388 ± 72 (4) | 14.3 ± 8.4 (5) | 0.407 ± 0.356 (5) | 1.219 ± 1.617 (5) | 14.11 ± 10.82 (5) | 0.89 ± 0.61 (5) | 11.8 ± 7.7 (4) |
| 20 | 7.0 ± 0.5 (4) | 740 ± 124 (4) | 396 ± 14 (3) | 6.8 ± 3.9 (4) | 0.235 ± 0.331 (4) | 0.830 ± 1.416 (4) | 5.48 ± 3.15 (4) | 0.40 ± 0.51 (4) | 19.0 ± 5.2 (3) |
| 24 | 7.1 ± 0.8 (2) | 296 ± 162 (2) | 480 (1) | 7.1 ± 4.4 (2) | 0.155 ± 0.187 (2) | 0.027 ± 0.189 (2) | 2.63 ± 0.86 (2) | 0.26 ± 0.25 (2) | 7.0 ± 3.9 (2) |
| Abbreviation | Dominant Plant Community | Dominant Species/Taxa Included | |
|---|---|---|---|
| 1 | Amp_veg | Amphibious vegetation | Alisma plantago-aquatica L., Oenanthe aquatica (L.) Poiret, Rorippa amphibia (L.) Besser, Rorippa palustris (L.) Besser, Rumex maritimus L., amphibious seedlings (not identifiable) |
| 2 | Car_mix | Carex mix | Carex riparia Curtis, Carex spec. |
| 3 | Edge_mix | Edge mix | Poaceae (not further identified), but including Elymus repens (L.) Gould s. str., Arrhenatherum elatius (L.) P. Beauv. ex J. Presl & C. Presl, Bromus sterilis L., non-dominant stands of Phalaris arundinacea L., non-dominant stands of Nitrophilous perennials (see group 5), unidentified segetal species |
| 4 | Hyd | Hydrophytes | Lemna minor L., Spirodela polyrhiza (L.) Schleid., Potamogeton natans L., Batrachium spec. (characterized by floating leaves); Ceratophyllum submersum L. (potential area for submersed hydrophytes, characterized as open water area) |
| 5 | Nit_per | Nitrophilous perennials | Arctium lappa L., Cirsium arvense (L.) Scop., Crisium vulgaris (Savi) Ten., Galeopsis speciosa Mill., Urtica dioica L. s. l., Rumex obtusifolius L., Rumex spec. (not R. maritimus) |
| 6 | Flood | Flood-meadow | Alopecurus aequalis Sobol., Glyceria fluitans (L.) R. Br. |
| 7 | Others | Others | Anthriscus sylvestris (L.) Hoffm., Iris pseudacorus L., Juncus effusus L., Juncus spec. |
| 8 | Pha_mix | Phalaris mix | Phalaris arundinacea L. |
| 9 | Poa_mix | Poa mix | Poa palustris L., Poa trivialis L. |
| 10 | Phr_mix | Phragmites mix | Phragmites australis (Cav.) Trin. ex Steud. |
| 11 | Woody | Woody plants | Humulus lupulus L., Fraxinus excelsior L., Rosa spec., Salix alba L., Salix cinerea L., Salix spec., Sambucus nigra L., Sorbus aucuparia L., unidentified woody species |
| 12 | Spa_mix | Sparganium mix | Sparganium emersum Rehmann |
| 13 | Typ_mix | Typha latifolia | Typha latifolia L. |
| 14 | Unveg | Unvegetated | No vegetation |
| Marginal Test | Relationship | |||||
|---|---|---|---|---|---|---|
| Variable | Pseudo-F | P | Prop. | dbRDA1 | dbRDA2 | dbRDA3 |
| Area_tot | 3.401 | 0.004 | 7.8 | 0.409 | 0.046 | 0.726 |
| Water_Area | 4.962 | 0.000 | 11.0 | −0.306 | 0.793 | 0.358 |
| BS-S | 2.397 | 0.027 | 5.7 | |||
| BS-SO | 2.295 | 0.039 | 5.4 | −0.393 | 0.051 | −0.145 |
| SS-S | 4.077 | 0.002 | 9.3 | |||
| SS-SO | 3.451 | 0.006 | 7.9 | −0.414 | 0.255 | −0.192 |
| o | 6.345 | 0.000 | 13.7 | −0.641 | −0.538 | 0.515 |
| vo | 2.842 | 0.222 | 6.6 | −0.043 | −0.111 | −0.146 |
| s | 1.413 | 0.017 | 3.4 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pätzig, M.; Düker, E. Dynamic of Dominant Plant Communities in Kettle Holes (Northeast Germany) during a Five-Year Period of Extreme Weather Conditions. Water 2021, 13, 688. https://doi.org/10.3390/w13050688
Pätzig M, Düker E. Dynamic of Dominant Plant Communities in Kettle Holes (Northeast Germany) during a Five-Year Period of Extreme Weather Conditions. Water. 2021; 13(5):688. https://doi.org/10.3390/w13050688
Chicago/Turabian StylePätzig, Marlene, and Eveline Düker. 2021. "Dynamic of Dominant Plant Communities in Kettle Holes (Northeast Germany) during a Five-Year Period of Extreme Weather Conditions" Water 13, no. 5: 688. https://doi.org/10.3390/w13050688
APA StylePätzig, M., & Düker, E. (2021). Dynamic of Dominant Plant Communities in Kettle Holes (Northeast Germany) during a Five-Year Period of Extreme Weather Conditions. Water, 13(5), 688. https://doi.org/10.3390/w13050688