
Evaluation of Low-Head Ramped Weirs for a Potamodromous Cyprinid: Effects of Substrate Addition and Discharge on Fish Passage Performance, Stress and Fatigue

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
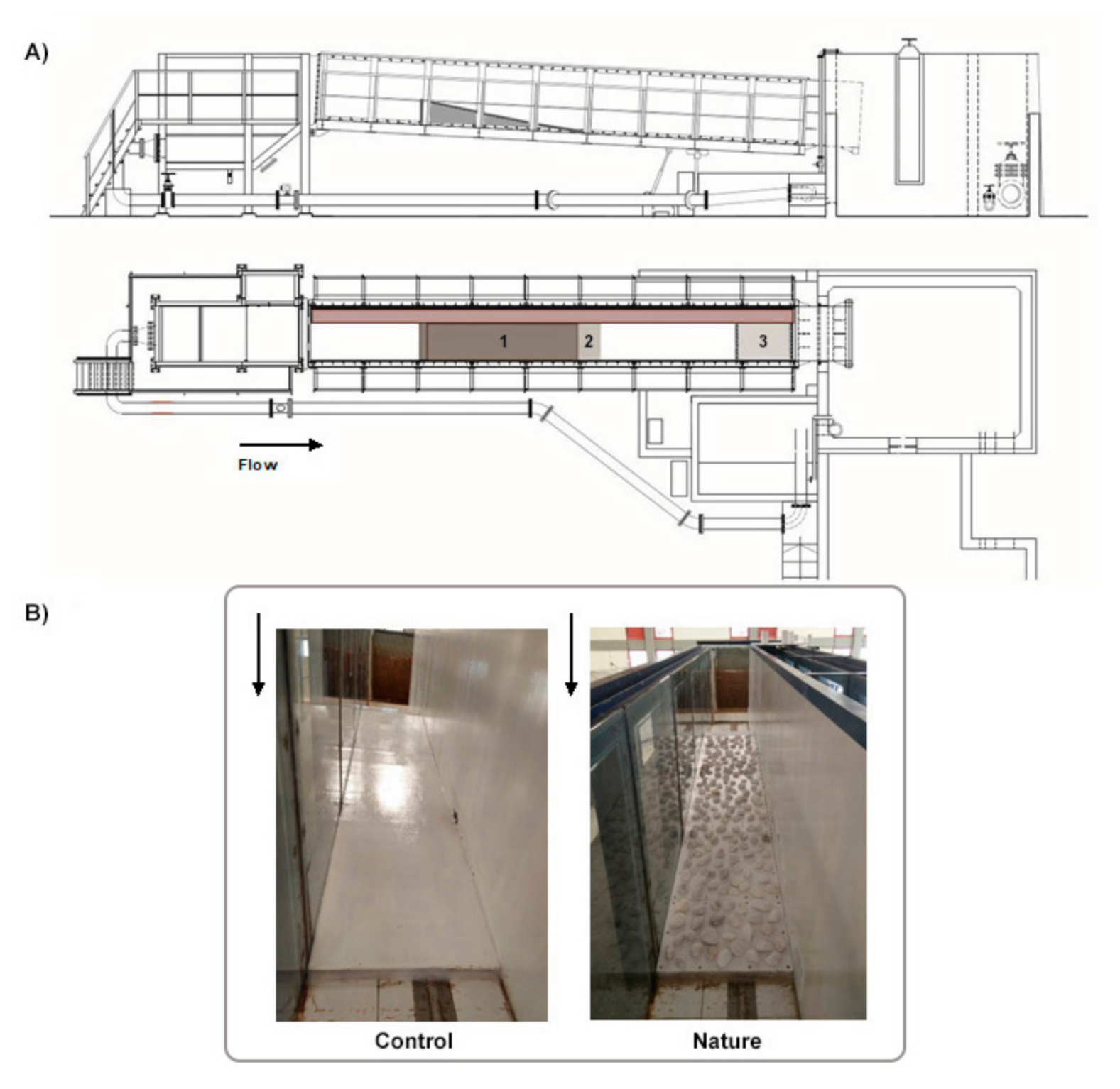
2.1. Experimental Facility
2.2. Fish Experiments
2.3. Data Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Birnie-Gauvin, K.; Franklin, P.; Wilkes, M.; Aarestrup, K. Moving beyond fitting fish into equations: Progressing the fish passage debate in the Anthropocene. Aquatic Conserv. Mar. Freshw. Ecosyst. 2019, 29, 1095–1105. [Google Scholar] [CrossRef]
- Belletti, B.; de Garcia Leaniz, C.; Jones, J.; Bizzi, S.; Börger, L.; Segura, G.; Castelletti, A.; van de Bund, W.; Aarestrup, K.; Barry, J.; et al. More than one million barriers fragment Europe’s rivers. Nature 2020, 588, 436–441. [Google Scholar] [CrossRef]
- Lucas, M.C.; Bubb, D.H.; Jang, M.-H.; Ha, K.; Masters, J.E.G. Availability of and access to critical habitats in regulated rivers: Effects of low-head barriers on threatened lampreys. Freshw. Biol. 2009, 54, 621–634. [Google Scholar] [CrossRef]
- Birnie-Gauvin, K.; Aarestrup, K.; Riis, T.M.O.; Jepsen, N.; Koed, A. Shining a light on the loss of rheophilic fish habitat in lowland rivers as a forgotten consequence of barriers, and its implications for management. Aqua. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 1345–1349. [Google Scholar] [CrossRef]
- Ovidio, M.; Philippart, J.C. The impact of small physical obstacles on upstream movements of six species of fish—Synthesis of a 5-year telemetry study in the River Meuse basin. Hydrobiologia 2002, 483, 55–69. [Google Scholar] [CrossRef]
- Calles, O.; Greenberg, L. Connectivity is a two-way street—The need for a holistic approach to fish passage problems in regulated rivers. River Res. Appl. 2009, 25, 1268–1286. [Google Scholar] [CrossRef]
- Gozlan, R.E.; Karimov, B.K.; Zadereev, E.; Kuznetsova, D.; Brucet, S. Status, trends, and future dynamics of freshwater ecosystems in Europe and Central Asia. Inland Waters 2019, 9, 78–94. [Google Scholar] [CrossRef]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [Green Version]
- Aarts, B.G.; Van Den Brink, F.W.; Nienhuis, P.H. Habitat loss as the main cause of the slow recovery of fish faunas of regulated large rivers in Europe: The transversal floodplain gradient. River Res. Appl. 2003, 20, 3–23. [Google Scholar] [CrossRef]
- Nilsson, C.; Reidy, C.A.; Dynesius, M.; Revenga, C. Fragmentation and flow regulation of the world’s large river systems. Science 2005, 308, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, S.; O’Hanley, J.R.; Newbold, L.R.; Kemp, P.S.; Diebel, M.W. A toolkit for optimizing fish passage barrier mitigation actions. J. Appl. Ecol. 2017, 54, 599–611. [Google Scholar] [CrossRef] [Green Version]
- Ordeix, M.; González, G.; Sanz-Ronda, F.J.; Santos, J.M. Restoring fish migration in the rivers of the Iberian Peninsula. In From Sea to Source 2.0. Protection and Restoration of Fish Migration in Rivers Worldwide; Brink, K., Gough, P., Royte, J., Schollema, P.P., Wanningen, H., Eds.; World Fish Migration Foundation: Groningen, The Netherlands, 2018; pp. 174–179. [Google Scholar]
- Branco, P.; Amaral, S.D.; Ferreira, M.T.; Santos, J.M. Do small barriers affect the movement of freshwater fish by increasing residency? Sci. Total Environ 2017, 581–582, 486–494. [Google Scholar] [CrossRef]
- de Garcia Leaniz, C. Weir removal in salmonid streams: Implications, challenges and practicalities. Hydrobiologia 2008, 609, 83–96. [Google Scholar] [CrossRef]
- 15. European Commission. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 establishing a framework for the community action in the field of water policy. Off. J. Eur. Comm. 2000, 327, 1–72.
- European Commission. Bringing Nature Back into Our Lives. EU 2030 Biodiversity Strategy; Document 52020DC0380; 2020. Available online: https://eur-lex.europa.eu/legalcontent/EN/TXT/?qid=1590574123338&uri=CELEX:52020 (accessed on 3 January 2021).
- Food and Agriculture Organization (FAO)/DVWK. Fish Passes—Design, Dimensions and Monitoring; FAO: Rome, Italy, 2002; p. 119. [Google Scholar]
- Baker, C.F. Effect of ramp length and slope on the efficacy of a baffled fish pass. J. Fish Biol. 2014, 84, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Baudoin, J.M.; Burgun, V.; Chanseau, M.; Larinier, M.; Ovidio, M.; Sremski, W.; Steinbach, P.; Voegtle, B. Assessing the Passage of Obstacles by Fish. Concepts, Design and Application; Onema: Paris, France, 2014; p. 200. [Google Scholar]
- Towler, B.; Mulligan, K.; Haro, A. Derivation and application of the energy dissipation factor in the design of fishways. Ecol. Eng. 2015, 83, 208–217. [Google Scholar] [CrossRef]
- Richer, E.E.; Fetherman, E.R.; Krone, E.A.; Wright, F.B.; Kondratieff, M.C. Multispecies Fish Passage Evaluation at a Rock-Ramp Fishway in a Colorado Transition Zone Stream. N. Am. J. Fish Manag. 2020, 40, 1510–1522. [Google Scholar] [CrossRef]
- Katopodis, C.; Aadland, L.P. Effective dam removal and river channel restoration approaches. Int. J. River Basin Manag. 2006, 4, 153–168. [Google Scholar] [CrossRef]
- Santos, J.M.; Branco, P.; Katopodis, C.; Ferreira, T.; Pinheiro, A. Retrofitting pool-and-weir fishways to improve passage performance of benthic fishes: Effect of boulder density and fishway discharge. Ecol. Eng. 2014, 73, 335–344. [Google Scholar] [CrossRef]
- Santos, J.M.; Reino, L.; Porto, M.; Oliveira, J.; Pinheiro, P.; Almeida, P.R.; Cortes, R.; Ferreira, M.T. Complex size-dependent habitat associations in potamodromous fish species. Aquat. Sci. 2011, 73, 233–245. [Google Scholar] [CrossRef]
- Ferreira, T.; Oliveira, J.; Caiola, N.; De Sostoa, A.; Casals, F.; Cortes, R.; Economou, A.; Zogaris, S.; Garcia-Jalon, D.; Ilhéu, M.; et al. Ecological traits of fish assemblages from Mediterranean Europe and their responses to human disturbance. Fish. Manag. Ecol. 2007, 14, 473–481. [Google Scholar] [CrossRef]
- Oliveira, J.M.; Segurado, P.; Santos, J.M.; Teixeira, A.; Ferreira, M.T.; Cortes, R.V. Modelling Stream-Fish Functional Traits in Reference Conditions: Regional and Local Environmental Correlates. PLoS ONE 2012, 7, e45787. [Google Scholar] [CrossRef]
- Kelly-Richards, S.; Silber-Coats, N.; Crootof, A.; Tecklin, D.; Bauer, C. Governing the transition to renewable energy: A review of impacts and policy issues in the small hydropower boom. Energy Policy 2017, 101, 251–264. [Google Scholar] [CrossRef]
- Solà, C.; Ordeix, M.; Pou-Rovira, Q.; Sellarès, N.; Queralt, A.; Bardina, M.; Casamitjana, A.; Munné, A. Longitudinal connectivity in hydromorphological quality assessments of rivers. The ICF index: A river connectivity index and its application to Catalan rivers. Limnetica 2011, 30, 273–292. [Google Scholar]
- Harris, J.H.; Kingsford, R.T.; Peirson, W.; Baumgartner, L.J. Mitigating the effects of barriers to freshwater fish migrations: The Australian experience. Mar. Freshw. Res. 2016, 68, 614–628. [Google Scholar] [CrossRef]
- Kemp, P.S.; O’Hanley, J.R. Procedures for evaluating and prioritising the removal of fish passage barriers: A synthesis. Fisheries Manag. Ecol. 2010, 17, 297–322. [Google Scholar] [CrossRef]
- Katopodis, C.; Gervais, R. Fish Swimming Performance Database and Analyses. Available online: http://www.dfo-mpo.gc.ca/csas-sccs/Publications/ResDocs-DocRech/2016/2016_002-eng.html (accessed on 3 January 2021).
- Newton, M.; Dodd, J.A.; Barry, J.; Boylan, P.; Adams, C.E. The impact of a small-scale riverine obstacle on the upstream migration of Atlantic Salmon. Hydrobiologia 2017, 806, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.J.; Boavida, I.; Almeida, V.; Cooke, S.J.; Pinheiro, A.N. Do artificial velocity refuges mitigate the physiological and behavioural consequences of hydropeaking on a freshwater Iberian cyprinid? Ecohydrology 2018, 11, e1983. [Google Scholar] [CrossRef]
- Romão, F.; Santos, J.M.; Katopodis, C.; Pinheiro, A.N.; Branco, P. How Does Season Affect Passage Performance and Fatigue of Potamodromous Cyprinids? An Experimental Approach in a Vertical Slot Fishway. Water 2018, 10, 395. [Google Scholar] [CrossRef] [Green Version]
- Stoot, L.J.; Cairns, N.A.; Cull, F.; Taylor, J.J.; Jeffrey, J.D.; Morin, F.; Mandelman, J.W.; Clark, T.D.; Cooke, S.J. Use of portable blood physiology point-of-care devices for basic and applied research on vertebrates: A review. Conserv. Physiol. 2014, 2, cou011. [Google Scholar] [CrossRef]
- Barton, B.A. Stress in Fishes: A diversity of responses with particular reference to changes in circulating corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef]
- Moreira, M.; Costa, M.J.; Valbuena-Castro, J.; Pinheiro, A.N.; Boavida, I. Cover or Velocity: What Triggers Iberian Barbel (Luciobarbus Bocagei) Refuge Selection under Experimental Hydropeaking Conditions? Water 2020, 12, 317. [Google Scholar] [CrossRef] [Green Version]
- Farrell, A.P. Comparisons of swimming performance in rainbow trout using constant acceleration and critical swimming speed tests. J. Fish. Biol. 2008, 72, 693–710. [Google Scholar] [CrossRef]
- White, A.J.; Schreer, J.F.; Cooke, S.J. Behavioral and physiological responses of the congeneric largemouth (Micropterus salmoides) and smallmouth bass (M. dolomieu) to various exercise and air exposure durations. Fish. Res. 2008, 89, 9–16. [Google Scholar] [CrossRef]
- Oliveira, I.C.; Alexandre, C.M.; Quintella, B.R.; Almeida, P.R. Impact of flow regulation for hydroelectric production in the movement patterns, growth and condition of a potamodromous fish species. Ecohydrology 2020, 13, e2250. [Google Scholar] [CrossRef]
- Amaral, S.D.; Quaresma, A.L.; Branco, P.; Romão, F.; Katopodis, C.; Ferreira, M.T.; Pinheiro, A.N.; Santos, J.M. Assessment of retrofitted ramped weirs to improve passage of potamodromous fish. Water 2019, 11, 2441. [Google Scholar] [CrossRef] [Green Version]
- Enders, E.C.; Gessel, M.H.; Williams, J.G. Development of successful fish passage structures for downstream migrants requires knowledge of their behavioural response to accelerating flow. Can. J. Fish. Aquat. Sci. 2009, 66, 2109–2117. [Google Scholar] [CrossRef]
- Amaral, S.D.; Branco, P.; Silva, A.T.; Katopodis, C.; Viseu, T.; Ferreira, M.T.; Pinheiro, A.N.; Santos, J.M. Upstream passage of potamodromous cyprinids over small weirs: The influence of key-hydraulic parameters. J. Ecohydraulics 2016, 1, 79–89. [Google Scholar] [CrossRef]
- Amaral, S.D.; Branco, P.; Katopodis, C.; Ferreira, M.T.; Pinheiro, A.N.; Santos, J.M. Passage Performance of Potamodromous Cyprinids over an Experimental Low-Head Ramped Weir: The Effect of Ramp Length and Slope. Sustainability 2019, 11, 1456. [Google Scholar] [CrossRef] [Green Version]
- Vogt, J.; Soille, P.; De Jager, A.; Rimaviciute, E.; Mehl, W.; Foisneau, S.; Bodis, K.; Dusart, J.; Paracchini, M.L.; Haastrup, P.; et al. A Pan-European River and Catchment Database; European Commission—Joint Research Centre—Institute for Environment and Sustainability: Luxembourg, 2007. [Google Scholar]
- [CEN] European Committee for Standardization. Water Quality: Sampling of Fish. with Electricity; The European Standard EN 14011; European Committee for Standardization: Brussels, Belgium, 2003. [Google Scholar]
- Santos, J.M.; Ferreira, M.T.; Godinho, F.N.; Bochechas, J. Efficacy of a nature-like bypass channel in a Portuguese lowland river. J. Appl. Ichthyol. 2005, 21, 381–388. [Google Scholar] [CrossRef]
- Benitez, J.-P.; Ovidio, M. The influence of environmental factors on the upstream movements of rheophilic cyprinids according to their position in a river basin. Ecol. Freshw. Fish. 2018, 27, 660–671. [Google Scholar] [CrossRef]
- Dinno, A. dunn.test: Dunn’s Test of Multiple Comparisons Using Rank Sums. R Package Version 1.2.3. 2015. Available online: http://CRAN.R-project.org/package=dunn.test (accessed on 30 November 2019).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://www.R-project.org/ (accessed on 30 November 2019).
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ For. PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). Wiley Statsref Stat. Ref. Online 2017. [Google Scholar] [CrossRef]
- Muraoka, K.; Nakanishi, S.; Kayaba, Y. Boulder arrangement on a rocky ramp fishway based on the swimming behavior of fish. Limnologica 2017, 62, 188–193. [Google Scholar] [CrossRef]
- Plesiński, K.; Bylak, A.; Radecki-Pawlik, A.; Mikołajczyk, T.; Kukuła, K. Possibilities of fish passage through the block ramp: Model-based estimation of permeability. Sci. Total Environ 2018, 631–632, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Wait, L.E.; Monk, S.K.; Rader, R.; Hotchkiss, R.H.; Belk, M.C. Effects of substrate on movement patterns and behavior of stream fish through culverts: An experimental approach. Sustainability 2019, 11, 470. [Google Scholar] [CrossRef] [Green Version]
- Montali-Ashworth, D.; Vowles, A.S.; De Almeida, G.; Kemp, P.S. Use of Cylindrical Bristle Clusters as a novel multispecies fish pass to facilitate upstream movement at gauging weirs. Ecol. Eng. 2020, 143, 105634. [Google Scholar] [CrossRef]
- Costa, M.J.; Lennox, R.J.; Katopodis, C.; Cooke, S.J. Is there evidence for flow variability as an organism-level stressor in fluvial fish? J. Ecohydraulics 2017, 2, 68–83. [Google Scholar] [CrossRef]
- Flodmark, L.E.W.; Urke, H.A.; Halleraker, J.H.; Arnekleiv, J.V.; Vollestad, L.A.; Poléo, A.B.S. Cortisol and glucose responses in juvenile brown trout subjected to a fluctuating flow regime in an artificial stream. J. Fish. Biol. 2002, 60, 238–248. [Google Scholar] [CrossRef]
- Kieffer, J.D. Limits to exhaustive exercise in fish. Comp. Biochem. Phys. A 2000, 126, 161–179. [Google Scholar] [CrossRef]
- Gleeson, T.T. Post-exercise lactate metabolism: A comparative review of sites, pathways, and regulation. Annu. Rev. Physiol. 1996, 58, 565–581. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| RD | Q | N | AP | AT | n | S | AE% | PE% | SR% | (± SD) |
|---|---|---|---|---|---|---|---|---|---|---|
| CONTROL | 55 | 20 | 572 | 317 | 20 | 14 | 55 | 4 | 70 | 19 (±15) |
| 110 | 20 | 11 | 8 | 8 | 3 | 73 | 38 | 38 | 32 (±18) | |
| NATURE | 55 | 20 | 208 | 476 | 20 | 17 | 229 | 4 | 85 | 26 (±18) |
| 110 | 20 | 47 | 27 | 12 | 8 | 57 | 30 | 67 | 19 (±15) |
| Factors | AP & AT | S | T | LACT | GLUC | |
|---|---|---|---|---|---|---|
| RD | F: | 3.412 | 4.312 | 1.769 | 0.514 | 0.118 |
| P: | 0.047 | 0.041 | 0.206 | 0.489 | 0.759 | |
| Q | F: | 26.848 | 26.950 | 16.540 | 4.441 | 3.196 |
| P: | 0.001 | 0.001 | 0.001 | 0.044 | 0.082 | |
| RD × Q | F: | 4.400 | 0.269 | 1.712 | 0.062 | 1.036 |
| P: | 0.021 | 0.626 | 0.203 | 0.821 | 0.337 | |
| Res | 76 | 76 | 76 | 76 | 76 | |
| Total | 79 | 79 | 79 | 79 | 79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amaral, S.D.; Branco, P.; Romão, F.; Ferreira, M.T.; Pinheiro, A.N.; Santos, J.M. Evaluation of Low-Head Ramped Weirs for a Potamodromous Cyprinid: Effects of Substrate Addition and Discharge on Fish Passage Performance, Stress and Fatigue. Water 2021, 13, 765. https://doi.org/10.3390/w13060765
Amaral SD, Branco P, Romão F, Ferreira MT, Pinheiro AN, Santos JM. Evaluation of Low-Head Ramped Weirs for a Potamodromous Cyprinid: Effects of Substrate Addition and Discharge on Fish Passage Performance, Stress and Fatigue. Water. 2021; 13(6):765. https://doi.org/10.3390/w13060765
Chicago/Turabian StyleAmaral, Susana D., Paulo Branco, Filipe Romão, Maria Teresa Ferreira, António N. Pinheiro, and José Maria Santos. 2021. "Evaluation of Low-Head Ramped Weirs for a Potamodromous Cyprinid: Effects of Substrate Addition and Discharge on Fish Passage Performance, Stress and Fatigue" Water 13, no. 6: 765. https://doi.org/10.3390/w13060765
APA StyleAmaral, S. D., Branco, P., Romão, F., Ferreira, M. T., Pinheiro, A. N., & Santos, J. M. (2021). Evaluation of Low-Head Ramped Weirs for a Potamodromous Cyprinid: Effects of Substrate Addition and Discharge on Fish Passage Performance, Stress and Fatigue. Water, 13(6), 765. https://doi.org/10.3390/w13060765