
A Morphological and Molecular Analysis of a Bloom of the Filamentous Green Alga Pithophora




Abstract
:1. Introduction
2. Materials and Methods
3. Results
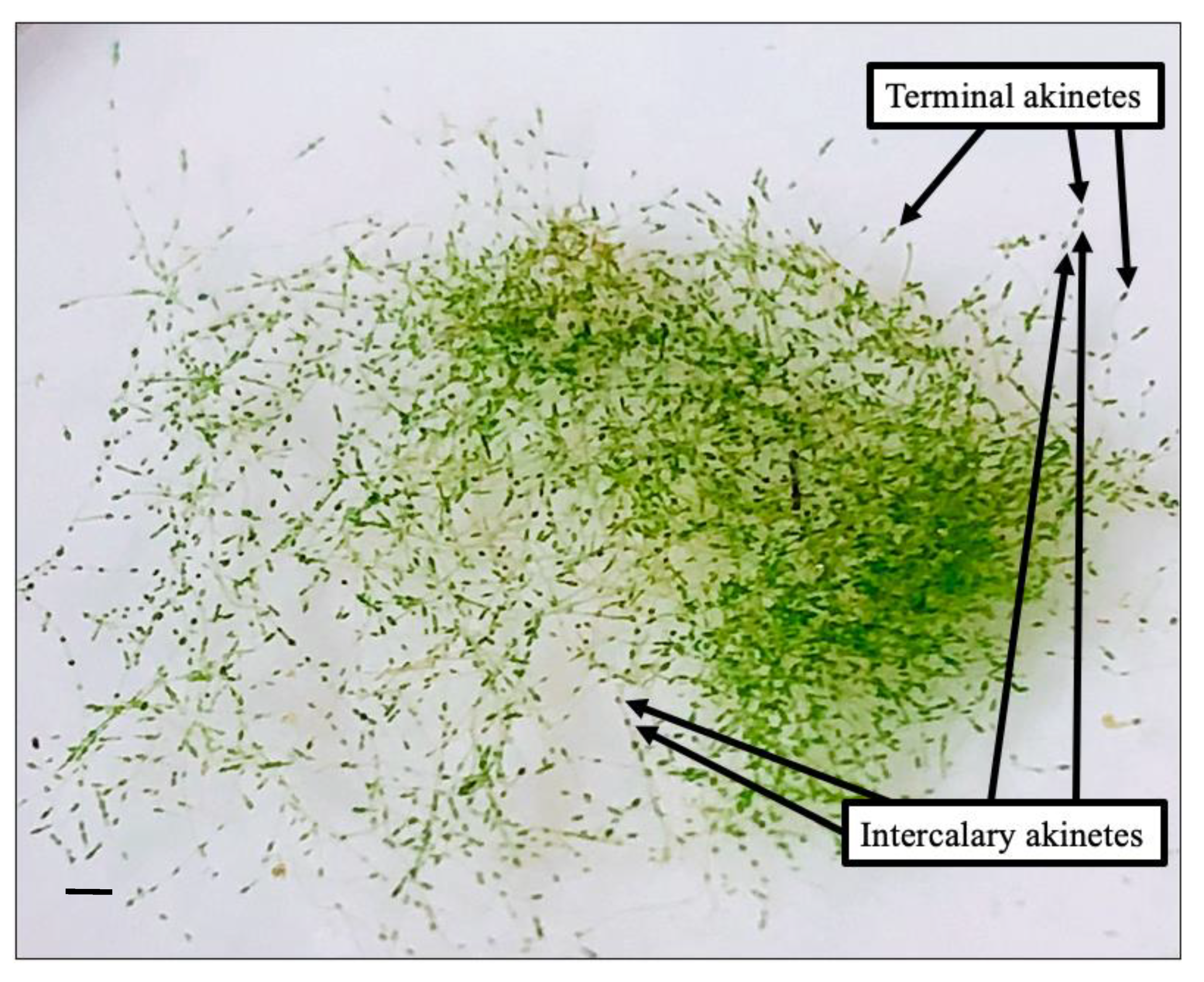
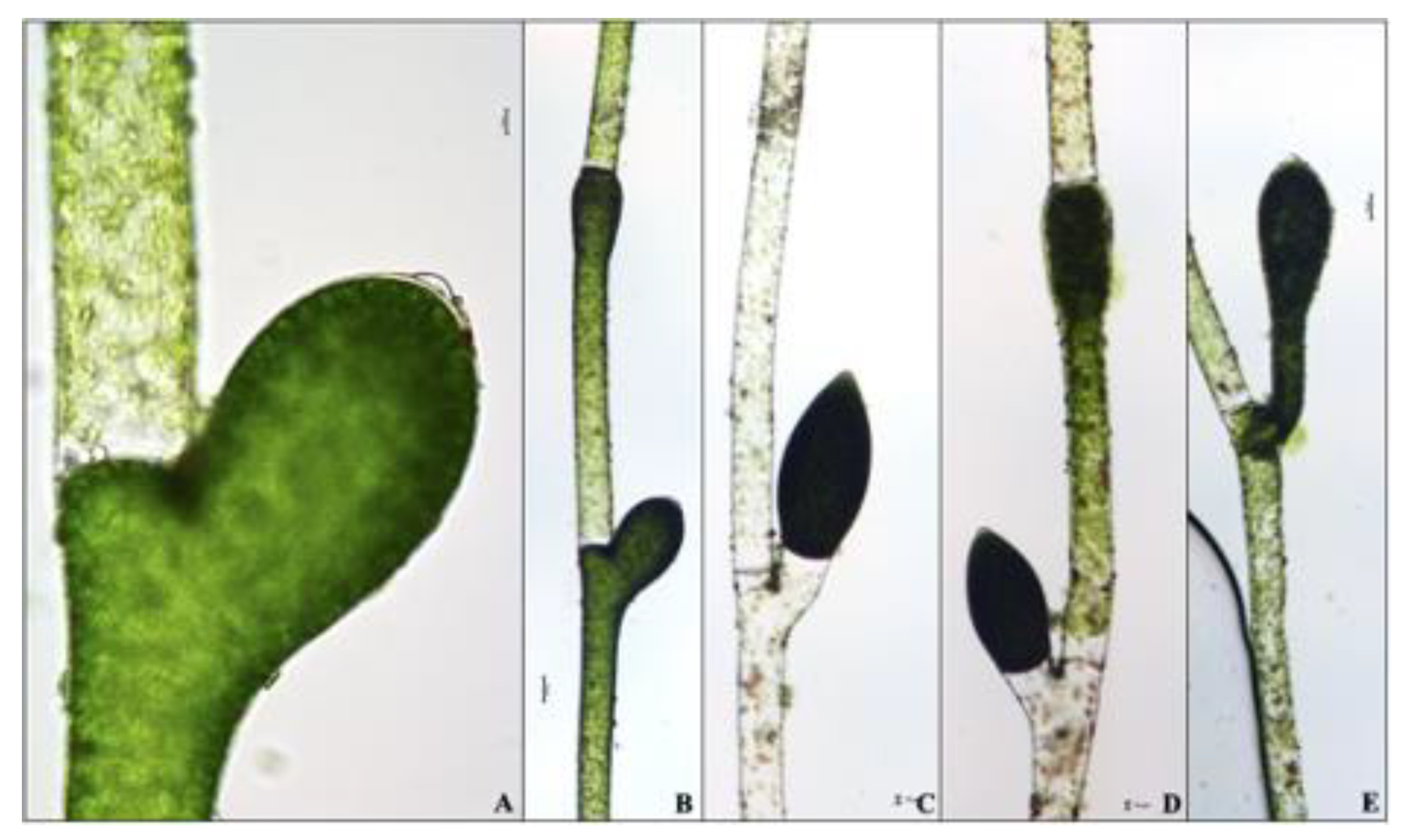
3.1. Morphological Analyses
3.2. DNA Sequences
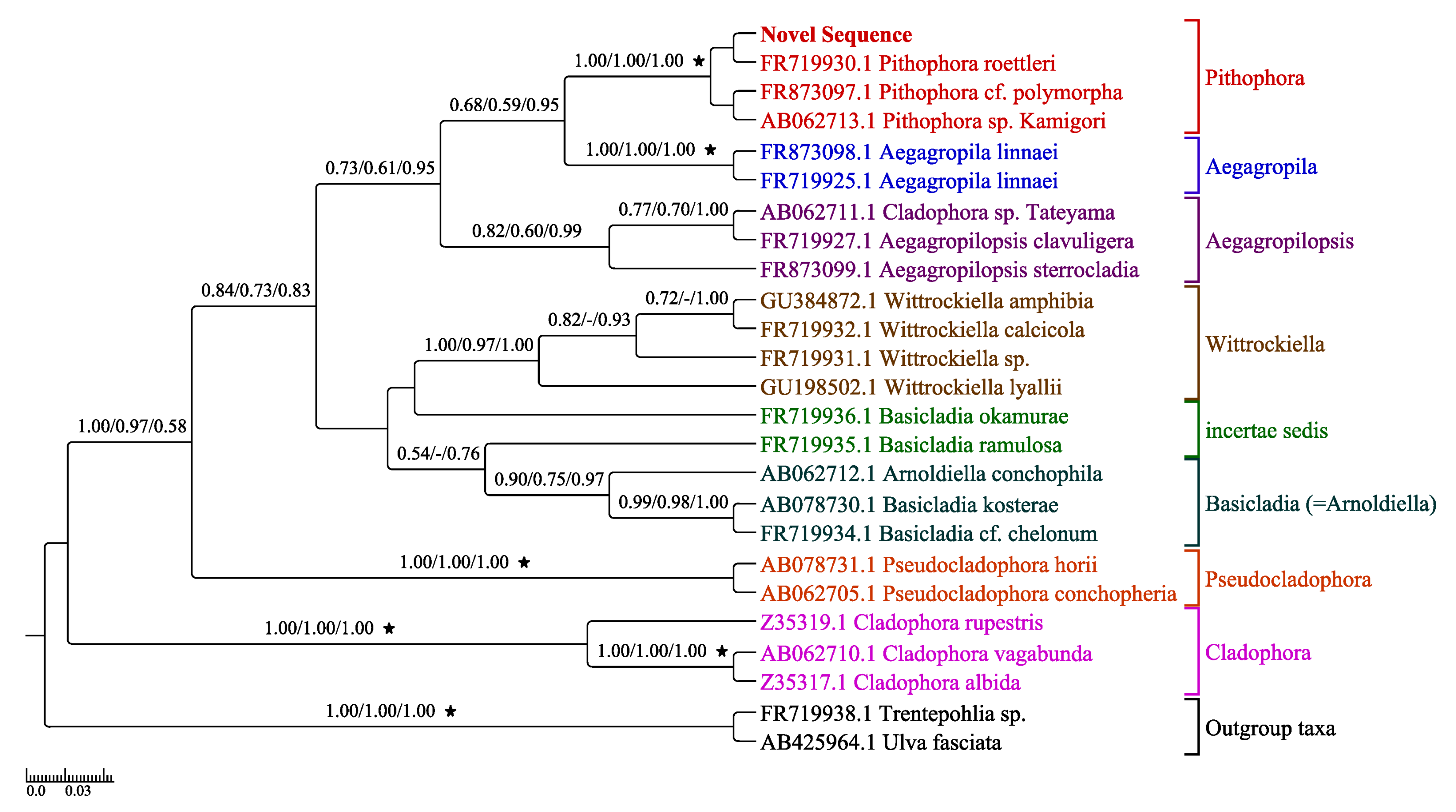
3.3. Phylogenetic Analysis
3.4. Morphological and Molecular Phylogenetic Analyses Congruence
4. Discussion
4.1. Morphological Analyses
4.2. Molecular Phylogenetic Analyses
4.3. Consensual Interpretation
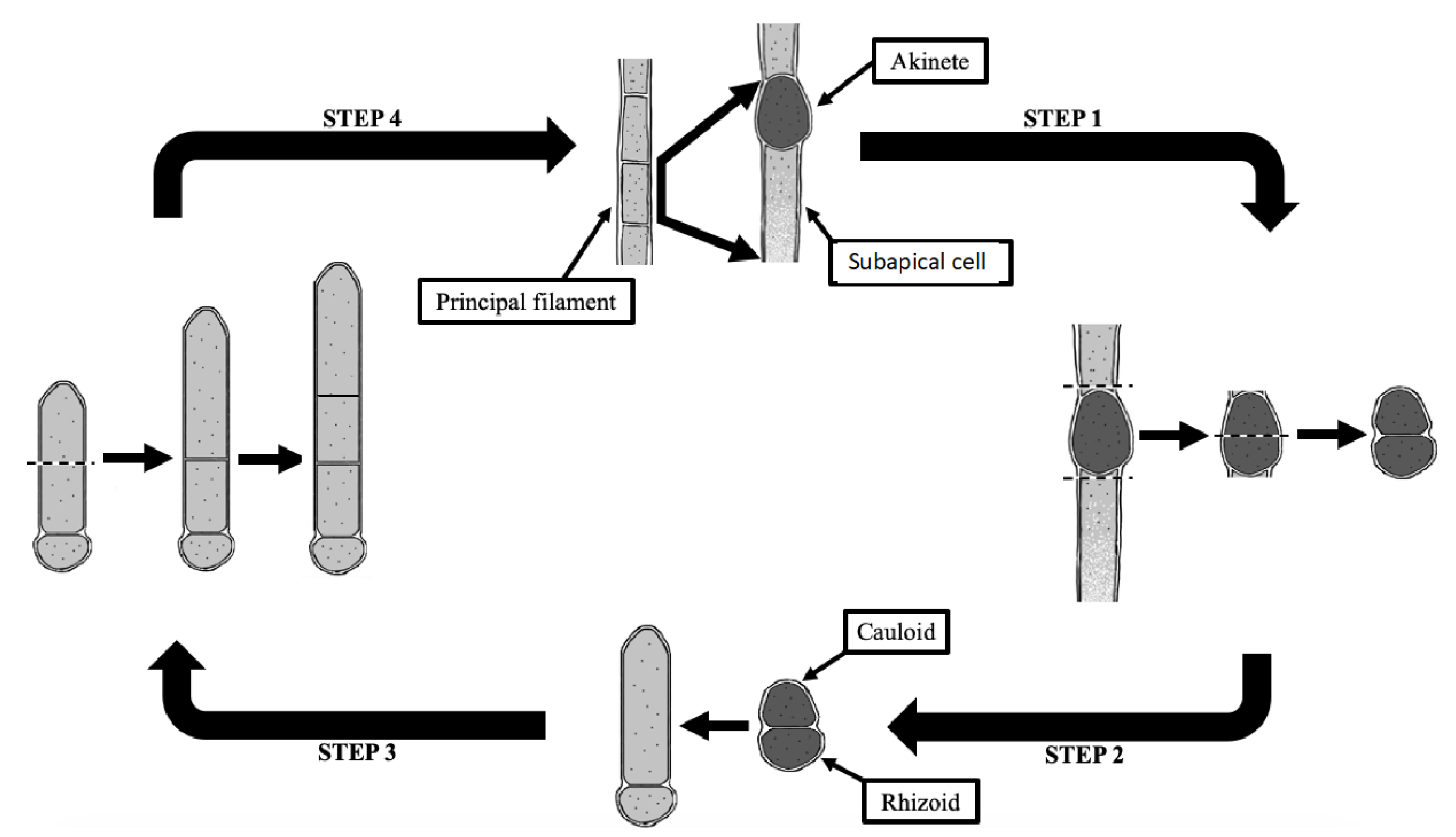
4.4. New Schematic Diagram
4.5. Inferred Ecology of Floating Mats
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mittelbach, G.G. Community Ecology; Sinauer Associates, Inc. Publishers: Sunderland, MA, USA, 2012. [Google Scholar]
- Oyedeji, A.A.; Abowei, J.F.N. The classification, distribution, control and economic important of aquatic plants. Intern. J. Fish. Aquat. Stud. 2012, 1, 118–128. [Google Scholar]
- Awasthi, A.K. Textbook of Algae; Vikas Publishing House: New Delhi, India, 2015; pp. 169–174. [Google Scholar]
- Fritsch, F.E. The subaerial and freshwater algal flora of the tropics. A phytogeographical and ecological study. Ann. Bot. 1907, 21, 235–275. [Google Scholar] [CrossRef]
- Pearlmutter, N.L.; Lembi, C.A. Structure and composition of Pithophora oedogonia (Chlorophyta) cell wall. J. Phycol. 1980, 16, 602–616. [Google Scholar] [CrossRef]
- Boedeker, C.; Leliaert, F.; Zuccarello, G.C. Molecular phylogeny of the Cladophoraceae (Cladophorales, Ulvophyceae), with the resurrection of Acrocladus Nägeli and Willeella Børgesen, and the description of Lurbica Gen. Nov. and Pseudoprhizoclonium gen. nov. J. Phycol. 2016, 52, 905–928. [Google Scholar] [CrossRef]
- Zulkifly, S.B.; Graham, J.M.; Young, E.B.; Mayer, R.J.; Piotroeski, M.J.; Smith, I.; Graham, L.E. The genus Cladophora Kützing (Ulvophyceae) as a globally distributed ecological engineer. J. Phycol. 2013, 49, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Prescott, G.W. Algae of the Western Great Lakes Area; WM. C. Brown Co. Inc.: Dubuque, Iowa, 1962; pp. 134–144. [Google Scholar]
- Saunders, L.L.; Kilham, S.S.; Fairchild, G.W.R.; Verb, R. Effects of small-scale environmental variation on metaphyton condition and community composition. Freshw. Biol. 2012, 57, 1884–1895. [Google Scholar] [CrossRef]
- Power, M.; Lowe, R.; Furey, F.; Welter, J.; Limm, M.; Finlay, J.; Bode, C.; Chang, S.; Goodrich, M.; Sculley, J. Algal mats and insect emergence in rivers under Mediterranean climates: Towards photogrammetric surveillance. Freshw. Biol. 2009, 54, 2101–2115. [Google Scholar] [CrossRef]
- Ansari, W.A.; Kashyap, S.P.; Prajapati, R.; Neelam, A. 2 Morpho-physiological and genetic variation of the algal collected from different water bodies of Varanasi and Chandauli region using ISSR molecular marker. Int. J. Tech. Res. Appl. 2015, 3, 56–61. [Google Scholar]
- Valiela, I.; McClelland, J.; Hauxwell, J.; Behr, P.J.; Hersh, D.; Foreman, K. Macroalgal blooms in shallow estuaries: Controls and ecophysiological and ecosystem consequences. Limnol. Oceanogr. 1997, 42, 1105–1118. [Google Scholar] [CrossRef] [Green Version]
- Dodds, W.K.; Gudder, D.A. The ecology of Cladophora. J. Phyc. 1992, 28, 415–427. [Google Scholar] [CrossRef]
- Boedeker, C. Phylogenetic, Taxonomic and Biogeographical Studies in the Pithophoraceae (Cladophorales, Chlorophyta); Wöhrmann Print Service: Zutphen, The Netherlands; Netherlands Centre for Biodiversity Naturalis: Leiden, The Netherlands, 2010. [Google Scholar]
- Leliaert, F.; Boedeker, C. Cladophorales. In The Green Seaweeds of Britain and Ireland; Brodie, J., Maggs, C.A., John, D.M., Eds.; The British Phycological Society: London, UK, 2007; pp. 131–183. [Google Scholar]
- Algarte, V.M.; Bichoff, A.; Dunck, B.; Rodrigues, L. First record of Pithophora oedogonia (Montagne) Wittrock (Pithophoraceae) in the Upper Paraná River floodplain, Brazil. Check List 2015, 11, 1722. [Google Scholar] [CrossRef] [Green Version]
- Boedeker, C.; Kelly, C.J.O.; Star, C.J.W.; Leliaert, F. Molecular phylogeny and taxonomy of the Aegagropila clade (Cladophorales, Ulvophyceae), including the description of Aegagropilopsis Gen. Nov. and Pseudocladophora gen. nov. J. Phycol. 2012, 48, 808–825. [Google Scholar] [CrossRef] [PubMed]
- Wittrock, V.B. On the Development and Systematic Arrangement of the Pithophoraceae, a New Order of Algae Nova Acta Regiae Societatis Scientiarum Upsaliensis Ser. 3, extra ordinem. Editum 1877, 19, 1–80. [Google Scholar]
- Leliaert, F.; Smith, D.R.; Moreau, H.; Herron, M.D.; Verbruggen, H.; Delwiche, C.F.; de Clerck, O. Phylogeny and molecular evolution of the green algae. CRC Crit. Rev. Plant Sci. 2012, 31, 1–46. [Google Scholar] [CrossRef] [Green Version]
- van den Hoek, C.; Mann, D.G.; Jahns, H.M. Algae an Introduction to Phycology; Cambridge Press University: New York, NY, USA, 1995. [Google Scholar]
- Chaudhary, B.R.; Singh, H.V. Influence of certain environmental factors of spore germination and spore differentiation in Pithophora oedogonia. Folia Microbiol. 1987, 32, 339–344. [Google Scholar] [CrossRef]
- Sukenik, A.; Maldener, I.; Delhaye, T.; Viner-Mozzini, Y.; Sela, D.; Bormans, M. Carbon assimilation and accumulation of cyanophycin during the development of dormant cells (akinetes) in the cyanobacterium Aphanizomenon ovalisporum. Front. Microbiol. 2015, 6, 1067. [Google Scholar] [CrossRef] [Green Version]
- O’Neal, S.W.; Lembi, C.A. Physiological changes during germination of Pithophora oedogonia (Chlorophyceae) akinetes. J. Phyc. 1983, 19, 193–199. [Google Scholar] [CrossRef]
- Collins, F.S. The Green Algae of North America; Tufts College Press: Medford, MA, USA, 1909. [Google Scholar]
- Kapraun, D.F. Nuclear DNA content estimates in green algal lineages: Chlorophyta and Streptophyta. Ann. Bot. 2007, 99, 677–701. [Google Scholar] [CrossRef] [PubMed]
- Del Cortona, A.; Leliaert, F.; Bogaert, K.A.; Verbrugeen, H.; Vandepoele, K.; Clerck, O. The plastid genome in Cladophorales green algae is encoded by hairpin chromosomes. Curr. Biol. 2017, 27, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Meade, M.J.; Proulex, G.C.R.; Manoylov, K.M.; Cahoon, A.B. Chloroplast mRNAs are 3′ polyuridlylated in the green alga Pithophora roettleri (Cladophorales). J. Phycol. 2020, 56, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetic analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MRBAYES 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, B.G. Phylogenetic Trees Made Easy: A How-to Manual, 4th ed.; Sinauer Associates, Inc.: Sunerland, MA, USA, 2011. [Google Scholar]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, S.; Entwisle, T.J. Non-marine algae of Australia: 6. Cladophoraceae (Chlorophyta). Telopea 2004, 10, 731–748. [Google Scholar]
- Hanyuda, T.; Wakana, I.; Arai, S.; Miyaji, K.; Watano, Y.; Ueda, K. Phylogenetic relationships within Cladophorales (Ulvophyceae, Chlorophyta) inferred from 18S rRNA gene sequences with special reference to Aegagropila linnaei. J. Phycol. 2002, 38, 564–571. [Google Scholar] [CrossRef]
- Lembi, C.A.; Pearlmutter, N.L.; Spencer, D.F. Life Cycle, Ecology, and Management Considerations of the Green Filamentous Algal, Pithophora; Indiana Water Res. Center Technology Reports, Paper 130; Purdue University: West Lafayette, IN, USA, 1980. [Google Scholar]
- NOAA National Centers for Environmental Information, State of the Climate: National Climate Report for October 2016. Available online: https://www.ncdc.noaa.gov/sotc/national/201610 (accessed on 15 January 2020).
- Paerl, H.W.; Huisman, J. Blooms like it hot. Am. Assoc. Adv. Sci. 2008, 320, 57–58. [Google Scholar] [CrossRef] [Green Version]
- Sundbäck, K.; Carlson, L.; Nilsson, C.; Jönsson, B.; Wulff, A.; Odmark, S. Response of benthic microbial mats to drifting green algal mats. Aquat. Microb. Ecol. 1996, 10, 195–208. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | LSU | SSU |
|---|---|---|
| Aegagropila | ||
| Aegagropila linnaei Kützing | EU655697.1 | FR719925.1 |
| EU655701.1 | FR873098.1 | |
| Aegagropilopsis | ||
| Cladophora clavuligera Grunow | FR719939.1 | FR719927.1 |
| Cladophora sterrocladia Skuja | FR719940.1 | FR873099.1 |
| Cladophora sp. ‘Tateyama’ | / | AB062711.1 |
| Basicladia (=Arnoldiella) | ||
| Arnoldiella conchophila V.V. Mill. | / | AB062712.1 |
| Basicladia cf. chelonum (Collins) | FR719947.1 | FR719934.1 |
| W.E. Hoffm. et. Tilden | ||
| Basicladia kosterae (C. Hoek) Garbary | FR719946.1 | AB078730.1 |
| Cladophora | ||
| Cladophora albida (Nees) Kützing | AM503433.1 | Z35317.1 |
| Cladophora rupestris (Linnaeus) Kützing | AJ544764.1 | Z35319.1 |
| Cladophora vagabunda (Linnaeus) | AJ544760.1 | AB062710.1 |
| van den Hoek | ||
| incertae sedis | ||
| Basicladia okamurae (Ueda) Garbary | FR719949.1 | FR719936.1 |
| Basicladia ramulosa Ducker | FR719948.1 | FR719935.1 |
| Pithophora | ||
| Pithophora sp. ‘Kamigori’ | / | AB062713.1 |
| Pithophora cf. polymorpha Wittrock | FR873096.1 | FR873097.1 |
| Pithophora roettleri (Roth) Wittrock | FR719942.1 | FR719930.1 |
| Pseudocladophora | ||
| Cladophora conchopheria Sakai | FR719951.1 | AB062705.1 |
| Cladophora horii C. Hoek et. Chihara | AJ544728.1 | AB078731.1 |
| Wittrockiella | ||
| Wittrockiella amphibia (Collins) | GU384873.1 | GU384872.1 |
| Boedeker et. G.I. Hansen | ||
| Cladophorella calcicola F.E. Fritsch | FR719944.1 | FR719932.1 |
| Wittrockiella layallii (Harvey) | GU198503.1 | GU198502.1 |
| C. Hoek, Ducker et. Womersley | ||
| Wittrockiella sp. | FR719943.1 | FR719931.1 |
| Outgroup taxa | ||
| Trentepohlia sp. | FR719952.1 | FR719938.1 |
| Ulva fasciata Delile | AJ544726.1 | AB425964.1 |
| Function | Gene | Organelle | GenBank Accession Number | Percent Identity to Pithophora sp. (UTEX 787) |
|---|---|---|---|---|
| atpA | Plastid | not archived | nd | |
| ATP synthase | atpB | Plastid | MN049190 | 94.3% |
| atpH | Plastid | MN049185 | 97.2% | |
| Cytochrome b6f complex | ||||
| petA | Plastid | MN049188 | 85.9% | |
| petB | Plastid | MN049187 | 93.7% | |
| Photosystem I | ||||
| psaA | Plastid | not archived | nd | |
| psaB | Plastid | MN049189 | 76.2% | |
| Photosystem II | ||||
| psbA | Plastid | not archived | nd | |
| psbB | Plastid | not archived | nd | |
| psbC | Plastid | MN049186 | 90.3% | |
| Rubisco | ||||
| rbcL | Plastid | MN049191 | 88.3% | |
| Cytochrome c oxidase subunit | ||||
| cox1 | Mitochondria | MN049192 | nd | |
| Ribosome | ||||
| SSU, ITS1, 5.8S, ITS2, LSU | Nucleus | MN017042 | 100% * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lor, B.; Zohn, M.; Meade, M.J.; Cahoon, A.B.; Manoylov, K.M. A Morphological and Molecular Analysis of a Bloom of the Filamentous Green Alga Pithophora. Water 2021, 13, 760. https://doi.org/10.3390/w13060760
Lor B, Zohn M, Meade MJ, Cahoon AB, Manoylov KM. A Morphological and Molecular Analysis of a Bloom of the Filamentous Green Alga Pithophora. Water. 2021; 13(6):760. https://doi.org/10.3390/w13060760
Chicago/Turabian StyleLor, Blia, Merry Zohn, Marcus J. Meade, A. Bruce Cahoon, and Kalina M. Manoylov. 2021. "A Morphological and Molecular Analysis of a Bloom of the Filamentous Green Alga Pithophora" Water 13, no. 6: 760. https://doi.org/10.3390/w13060760
APA StyleLor, B., Zohn, M., Meade, M. J., Cahoon, A. B., & Manoylov, K. M. (2021). A Morphological and Molecular Analysis of a Bloom of the Filamentous Green Alga Pithophora. Water, 13(6), 760. https://doi.org/10.3390/w13060760