
Variation in Xylem Hydraulic Structure and Function of Two Mangrove Species across a Latitudinal Gradient in Eastern Australia



 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sites and Plant Materials
2.2. Maximum Vessel Length
2.3. Stem Vulnerability Curve
2.4. Stem Hydraulic Conductivity and Huber Value
2.5. Wood Density and Sapwood Water Content
2.6. Water Potential and Hydraulic Safety Margin
2.7. Wood Anatomy
2.8. Statistics Analysis
3. Results
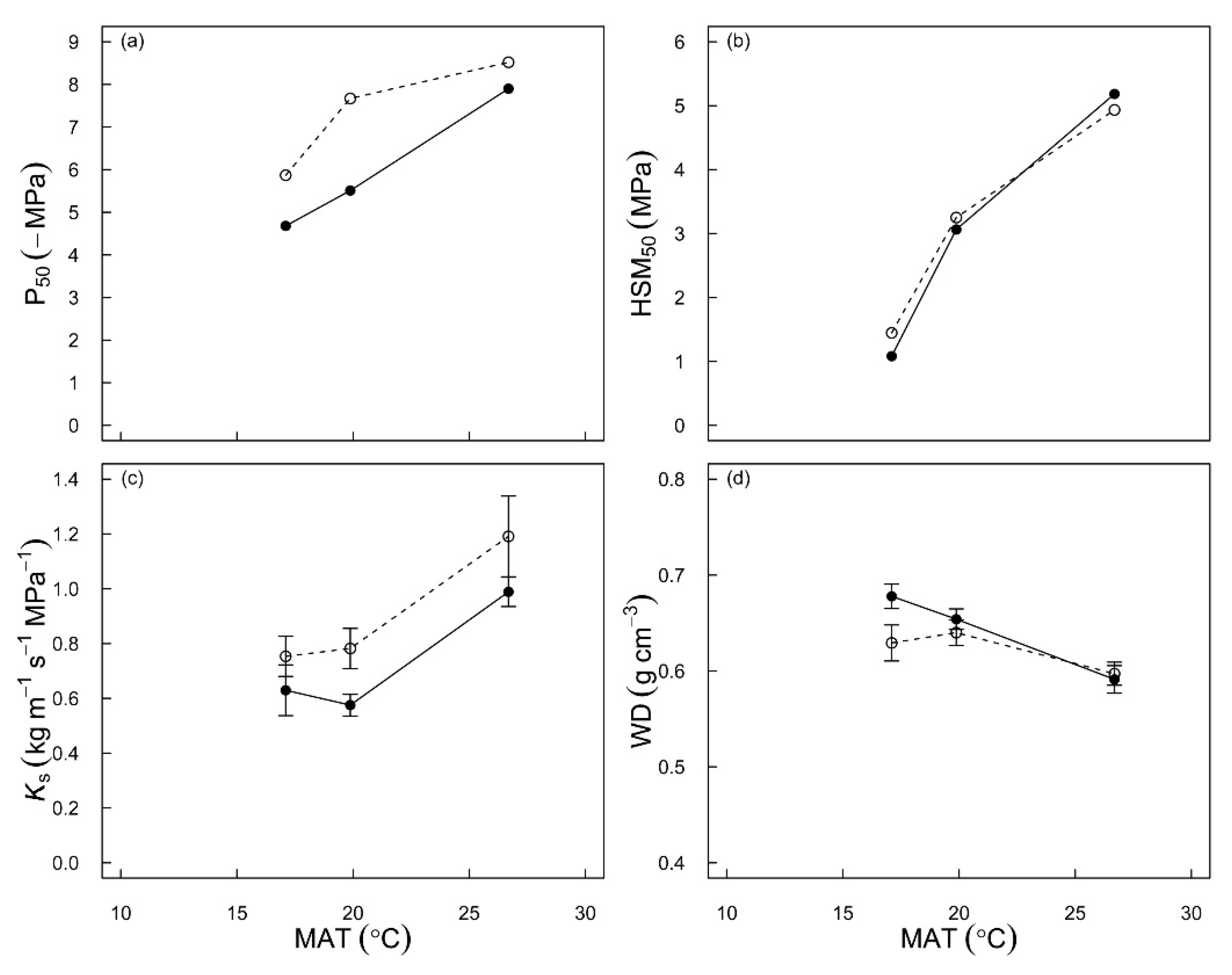
3.1. Changes in Xylem Hydraulic Traits along the Latitudinal Gradient
3.2. Difference in Hydraulic Traits and Plasticity between Two Species
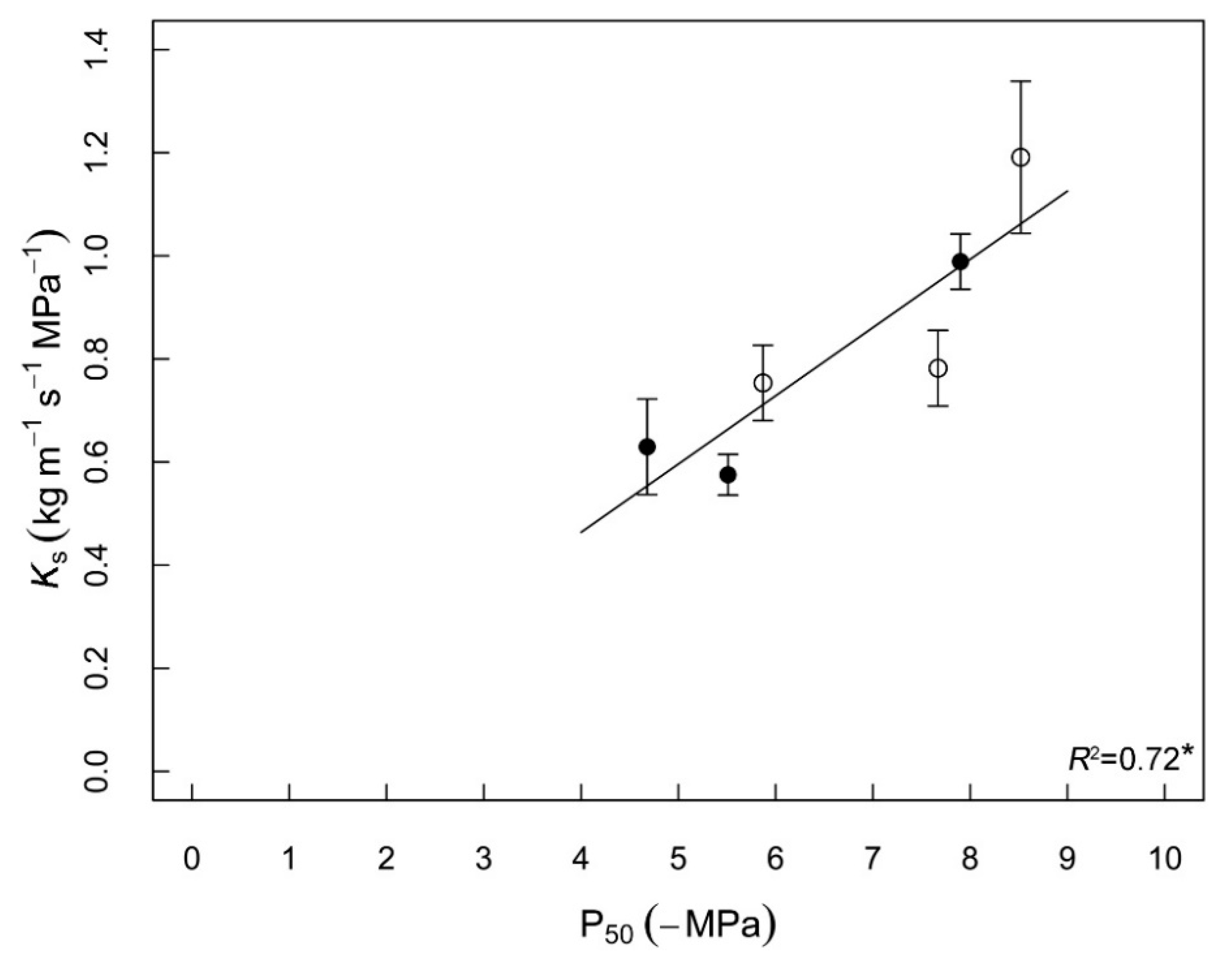
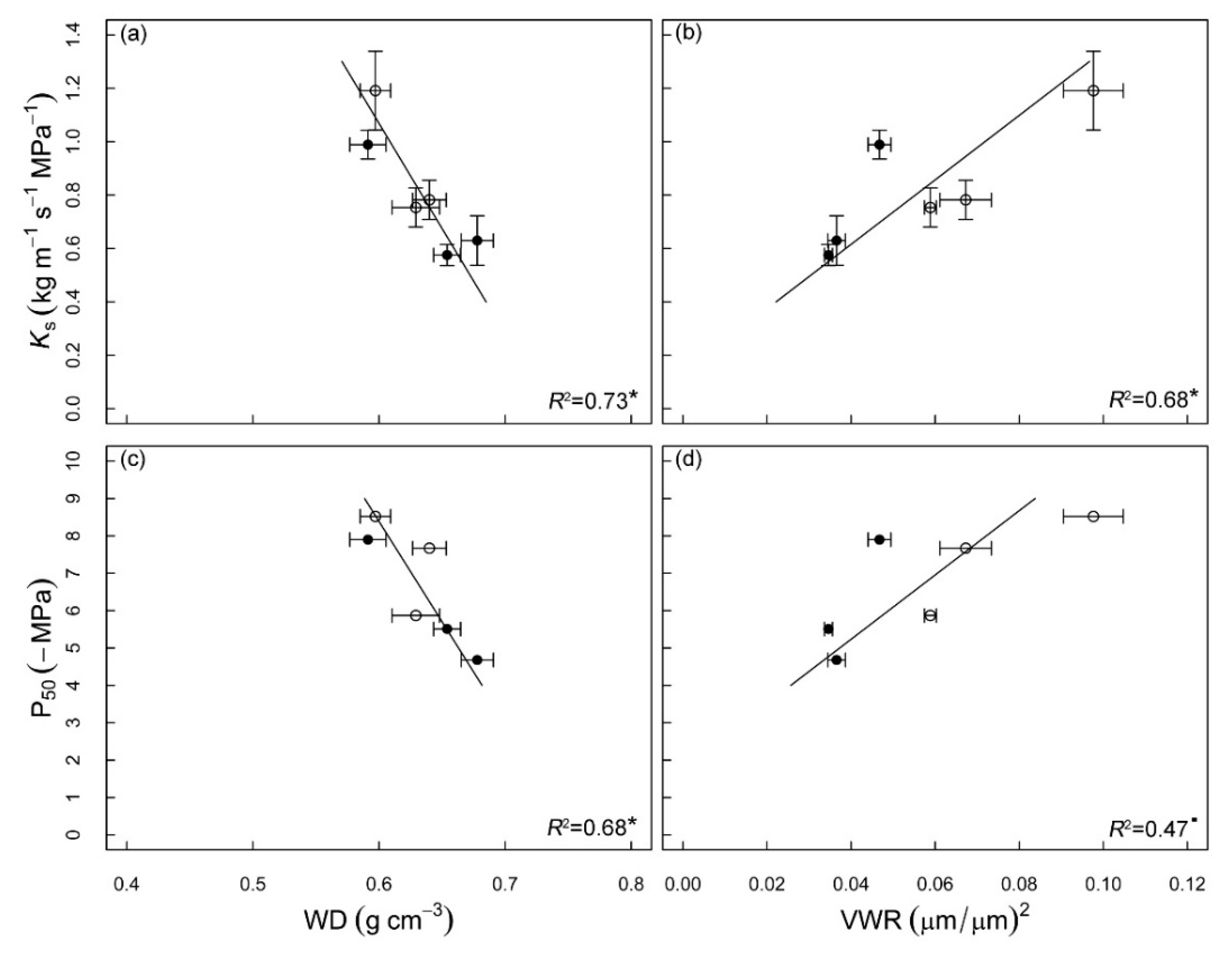
3.3. Correlations between Hydraulic Traits
4. Discussion
4.1. Variation in Hydraulic Traits along the Latitudinal Gradient
4.2. Different Hydraulic Plasticity between Two Species
4.3. Implications for Mangrove Response to Climate Change
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duke, N.C.; Ball, M.C.; Ellison, J.C. Factors influencing biodiversity and distributional gradients in mangroves. Glob. Ecol. Biogeogr. Lett. 1998, 7, 27; [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.T.; Stanton, D.E.; Schmitz, N.; Farquhar, G.D.; Ball, M.C. Growth responses of the mangrove Avicennia marina to salinity: Development and function of shoot hydraulic systems require saline conditions. Ann. Bot. 2015, 115, 397–407; [Google Scholar] [CrossRef] [PubMed]
- Reef, R.; Lovelock, C.E. Regulation of water balance in mangroves. Ann. Bot. 2015, 115, 385–395; [Google Scholar] [CrossRef] [Green Version]
- Quisthoudt, K.; Schmitz, N.; Randin, C.F.; Dahdouh-Guebas, F.; Robert, E.M.R.; Koedam, N. Temperature variation among man-grove latitudinal range limits worldwide. Trees 2012, 26, 1919–1931; [Google Scholar] [CrossRef]
- Nguyen, H.T.; Meir, P.; Sack, L.; Evans, J.R.; Oliveira, R.S.; Ball, M.C. Leaf water storage increases with salinity and aridity in the mangrove Avicennia marina: Integration of leaf structure, osmotic adjustment and access to multiple water sources. Plant Cell Environ. 2017, 40, 1576–1591; [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barraclough, A.D.; Cusens, J.; Zweifel, R.; Leuzinger, S. Environmental drivers of stem radius change and heterogeneity of stem radial water storage in the mangrove Avicennia marina (Forssk.) Vierh. Agric. For. Meteorol. 2020, 280, 107764; [Google Scholar] [CrossRef]
- Fazlioglu, F.; Chen, L. Introduced non-native mangroves express better growth performance than co-occurring native man-groves. Sci. Rep. 2020, 10, 1–11; [Google Scholar] [CrossRef]
- Sobrado, M.A. Relation of water transport to leaf gas exchange properties in three mangrove species. Trees 2000, 14, 258–262; [Google Scholar] [CrossRef]
- Lovelock, C.E.; Feller, I.C.; Ball, M.C.; Bettina, M.J.E.; Ewe, M.L. Differences in plant function in phosphorus- and nitro-gen-limited mangrove ecosystems. New Phytol. 2006, 172, 514–522; [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755; [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyree, M.T.; Davis, S.D.; Cochard, H. Biophysical perspectives of xylem evolution: Is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction? IAWA J. 1994, 15, 335–360; [Google Scholar] [CrossRef] [Green Version]
- Melcher, P.J.; Goldstein, G.; Meinzer, F.C.; Yount, D.E.; Jones, T.J.; Holbrook, N.M.; Huang, C.X. Water relations of coastal and estuarine Rhizophora mangle: Xylem pressure potential and dynamics of embolism formation and repair. Oecologia 2001, 126, 182–192; [Google Scholar] [CrossRef]
- Ewers, F.W.; Lopez-Portillo, J.; Angeles, G.; Fisher, J.B. Hydraulic conductivity and embolism in the mangrove tree La-guncularia racemosa. Tree Physiol. 2004, 24, 1057–1062; [Google Scholar] [CrossRef] [Green Version]
- Jiang, G.; Goodale, U.M.; Liu, Y.; Hao, G.; Cao, K. Salt management strategy defines the stem and leaf hydraulic charac-teristics of six mangrove tree species. Tree Physiol. 2017, 37, 389–401; [Google Scholar] [CrossRef] [PubMed]
- Stuart, S.A.; Choat, B.; Martin, K.C.; Holbrook, N.M.; Ball, M.C. The role of freezing in setting the latitudinal limits of mangrove forests. New Phytol. 2007, 173, 576–583; [Google Scholar] [CrossRef] [Green Version]
- Sperry, J.S.; Hacke, U.G.; Pittermann, J. Size and function in conifer tracheids and angiosperm vessels. Am. J. Bot. 2006, 93, 1490–1500; [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spec-trum. Ecol. Lett. 2009, 12, 351–366; [Google Scholar] [CrossRef] [PubMed]
- Verheyden, A.; De Ridder, F.; Schmitz, N.; Beeckman, H.; Koedam, N. High-resolution time series of vessel density in Kenyan mangrove trees reveal a link with climate. New Phytol. 2005, 167, 425–435; [Google Scholar] [CrossRef]
- Gleason, S.M.; Westoby, M.; Jansen, S.; Choat, B.; Hacke, U.G.; Pratt, R.B.; Bhaskar, R.; Brodribb, T.J.; Bucci, S.J.; Cao, K.; et al. Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species. New Phytol. 2016, 209, 123–136; [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Morin, H.; Deslauriers, A.; Plourde, P. Predicting xylem phenology in black spruce under climate warming. Glob. Chang. Biol. 2011, 17, 614–625; [Google Scholar] [CrossRef]
- Gricar, J.; Prislan, P.; Gryc, V.; Vavrcik, H.; de Luis, M.; Cufar, K. Plastic and locally adapted phenology in cambial sea-sonality and production of xylem and phloem cells in Picea abies from temperate environments. Tree Physiol. 2014, 34, 869–881; [Google Scholar] [CrossRef] [Green Version]
- Inoue, S.; Dang, Q.; Man, R.; Tedla, B. Northward migration of trembling aspen will increase growth but reduce resistance to drought-induced xylem cavitation. Botany 2019, 97, 627–638; [Google Scholar] [CrossRef]
- García-Cervigón, A.I.; Fajardo, A.; Caetano-Sánchez, C.; Camarero, J.J.; Olano, J.M. Xylem anatomy needs to change, so that conductivity can stay the same: Xylem adjustments across elevation and latitude in Nothofagus pumilio. Ann. Bot. 2020, 125, 1101–1112; [Google Scholar] [CrossRef]
- Puchi, P.F.; Castagneri, D.; Rossi, S.; Carrer, M. Wood anatomical traits in black spruce reveal latent water constraints on the boreal forest. Glob. Chang. Biol. 2020, 26, 1767–1777; [Google Scholar] [CrossRef]
- Girard, M.; Rossi, S.; Morin, H. Mapping events: Cambium phenology across the latitudinal distribution of black spruce. IAWA J. 2015, 36, 270–285; [Google Scholar] [CrossRef]
- Schuldt, B.; Knutzen, F.; Delzon, S.; Jansen, S.; Muller-Haubold, H.; Burlett, R.; Clough, Y.; Leuschner, C. How adaptable is the hydraulic system of European beech in the face of climate change-related precipitation reduction? New Phytol. 2016, 210, 443–458; [Google Scholar] [CrossRef] [Green Version]
- Bourne, A.E.; Creek, D.; Peters, J.M.R.; Ellsworth, D.S.; Choat, B. Species climate range influences hydraulic and stomatal traits in Eucalyptus species. Ann. Bot. 2017, 120, 123–133; [Google Scholar] [CrossRef] [PubMed]
- Macedo, T.M.; Lima, H.C.D.; de Souza, N.D.; Gonçalves, A.C.; Costa, C.G.; Barros, C.F. Intraspecific variation of Pau-brasilia echinata (Fabaceae) wood along a latitudinal gradient in Brazil. Flora 2019, 258, 151437; [Google Scholar] [CrossRef]
- Pfautsch, S.; Harbusch, M.; Wesolowski, A.; Smith, R.; Macfarlane, C.; Tjoelker, M.G.; Reich, P.B.; Adams, M.A. Climate determines vascular traits in the ecologically diverse genus Eucalyptus. Ecol. Lett. 2016, 19, 240–248; [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Sack, L.; Holbrook, N.M. Diversity of hydraulic traits in nine cordia species growing in tropical forests with contrasting precipitation. New Phytol. 2007, 175, 686–698; [Google Scholar] [CrossRef] [PubMed]
- Madrid, E.N.; Armitage, A.R.; LÃ Pez-Portillo, J. Avicennia germinans (black mangrove) vessel architecture is linked to chilling and salinity tolerance in the Gulf of Mexico. Front. Plant Sci. 2014, 5, 503; [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López, R.; Cano, F.J.; Choat, B.; Cochard, H.; Gil, L. Plasticity in vulnerability to cavitation of Pinus canariensis occurs only at the driest end of an aridity gradient. Front. Plant Sci. 2016, 7, 769; [Google Scholar] [CrossRef] [Green Version]
- Lamy, J.; Delzon, S.; Bouche, P.S.; Alia, R.; Vendramin, G.G.; Cochard, H.; Plomion, C. Limited genetic variability and phenotypic plasticity detected for cavitation resistance in a Mediterranean pine. New Phytol. 2014, 201, 874–886; [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cao, K.; Fan, Z.; Zhang, J. Potential hydraulic efficiency in angiosperm trees increases with growth-site tem-perature but has no trade-off with mechanical strength. Glob. Ecol. Biogeogr. 2013, 22, 971–981; [Google Scholar] [CrossRef]
- Morris, H.; Gillingham, M.A.F.; Plavcová, L.; Gleason, S.M.; Olson, M.E.; Coomes, D.A.; Fichtler, E.; Klepsch, M.M.; Mar-tínez-Cabrera, H.I.; McGlinn, D.J.; et al. Vessel diameter is related to amount and spatial arrangement of axial pa-renchyma in woody angiosperms. Plant Cell Environ. 2018, 41, 245–260; [Google Scholar] [CrossRef] [PubMed]
- He, P.; Gleason, S.M.; Wright, I.J.; Weng, E.; Liu, H.; Zhu, S.; Lu, M.; Luo, Q.; Li, R.; Wu, G.; et al. Growing-season tem-perature and precipitation are independent drivers of global variation in xylem hydraulic conductivity. Glob. Chang. Biol. 2020, 26, 1833–1841; [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Blackman, C.J.; Choat, B.; Duursma, R.A.; Rymer, P.D.; Medlyn, B.E.; Tissue, D.T. Tree hydraulic traits are coor-dinated and strongly linked to climate-of-origin across a rainfall gradient. Plant Cell Environ. 2018, 41, 646–660; [Google Scholar] [CrossRef]
- Wu, Y.; Ricklefs, R.E.; Huang, Z.; Zan, Q.; Yu, S. Winter temperature structures mangrove species distributions and as-semblage composition in China. Glob. Ecol. Biogeogr. 2018, 27, 1492–1506; [Google Scholar] [CrossRef]
- Gilman, E.L.; Ellison, J.; Duke, N.C.; Field, C. Threats to mangroves from climate change and adaptation options: A review. Aquat. Bot. 2008, 89, 237–250; [Google Scholar] [CrossRef]
- Duke, N.C.; Kovacs, J.M.; Griffiths, A.D.; Preece, L.; Hill, D.J.E.; van Oosterzee, P.; Mackenzie, J.; Morning, H.S.; Burrows, D. Large-scale dieback of mangroves in Australia’s Gulf of Carpentaria: A severe ecosystem response, coincidental with an unu-sually extreme weather event. Mar. Freshw. Res. 2017, 68, 1816–1829; [Google Scholar] [CrossRef]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and distribution of mangrove forests of the world using earth observation satellite data. Glob. Ecol. Biogeogr. 2011, 20, 154–159; [Google Scholar] [CrossRef]
- Lovelock, C.E.; Feller, I.C.; Reef, R.; Hickey, S.; Ball, M.C. Mangrove dieback during fluctuating sea levels. Sci. Rep. 2017, 7, 1680; [Google Scholar] [CrossRef]
- Duke, N.C. Mangrove Floristics and Biogeography. In Tropical Mangrove Ecosystems Robertson, A.I.; Alongi, D.M., Ed.; American Geophysical Union: Washington, DC, USA, 1992; Volume 41, pp. 63–100. [Google Scholar]
- Marine Science Australia. Available online: https://www.ausmarinescience.com/marine-science-basics/oceanography-of-australia/ (accessed on 28 December 2020).
- Sea Surface Salinity Remote Sensing Project. Available online: http://www.salinityremotesensing.ifremer.fr/sea-surface-salinity/salinity-distribution-at-the-ocean-surface (accessed on 28 December 2020).
- WorldClim. Available online: https://www.worldclim.org/ (accessed on 28 May 2020).
- Whitley, R.; Evans, B.; Pauwels, J.; Hutchinson, M.; Xu, T.; Wang, H. 2014. Potential evapotranspiration (approximated), an-nual precipitation: eMAST-R-Package 2.0, 0.01 degree, Australian coverage, 1970–2012, Macquarie University, Sydney, Australia. Obtained from http://dap.nci.org.au, made available by the Ecosystem Modelling and Scaling Infrastructure (eMAST, http://www.emast.org.au). Facility of the Terrestrial Ecosystem Research Network (TERN, http://www.tern.org.au) (accessed on 5 May 2016).
- Yang, J.; Medlyn, B.E.; De Kauwe, M.G.; Duursma, R.A. Applying the concept of ecohydrological equilibrium to predict steady state leaf area index. J. Adv. Model. Earth Syst. 2018, 10, 1740–1758; [Google Scholar] [CrossRef]
- Bureau of Meteorology. Available online: http://www.bom.gov.au (accessed on 19 May 2020).
- Ewers, F.W.; Fisher, J.B. Techniques for measuring vessel lengths and diameters in stems of woody plants. Am. J. Bot. 1989, 76, 645–656; [Google Scholar] [CrossRef]
- Cohen, S. Air method measurements of apple vessel length distributions with improved apparatus and theory. J. Exp. Bot. 2003, 54, 1889–1897; [Google Scholar] [CrossRef] [PubMed]
- Scholz, A.; Klepsch, M.; Karimi, Z.; Jansen, S. How to quantify conduits in wood? Front. Plant Sci. 2013, 4, 56; [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Portillo, J.; Ewers, F.W.; Angeles, G. Sap salinity effects on xylem conductivity in two mangrove species. Plant Cell Environ. 2005, 28, 1285–1292; [Google Scholar] [CrossRef]
- Sperry, J.S.; Donnelly, J.R.; Tyree, M.T. A method for measuring hydraulic conductivity and embolism in xylem. Plant Cell Environ. 1988, 11, 35–40; [Google Scholar] [CrossRef]
- Wheeler, J.K.; Huggett, B.A.; Tofte, A.N.; Rockwell, F.E.; Holbrook, N.M. Cutting xylem under tension or supersaturated with gas can generate PLC and the appearance of rapid recovery from embolism. Plant Cell Environ. 2013, 36, 1938–1949; [Google Scholar] [CrossRef] [PubMed]
- Torres-Ruiz, J.M.; Jansen, S.; Choat, B.; McElrone, A.J.; Cochard, H.; Brodribb, T.J.; Badel, E.; Burlett, R.; Bouche, P.S.; Brodersen, C.R.; et al. Direct X-ray microtomography observation confirms the induction of embolism upon xylem cutting under tension. Plant Physiol. 2015, 167, 40–43; [Google Scholar] [CrossRef] [Green Version]
- Choat, B.; Drayton, W.M.; Brodersen, C.; Matthews, M.A.; Shackel, K.A.; Wada, H.; McElrone, A.J. Measurement of vulnerability to water stress-induced cavitation in grapevine: A comparison of four techniques applied to a long-vesseled species. Plant Cell Environ. 2010, 33, 1502–1512; [Google Scholar] [CrossRef] [PubMed]
- Cochard, H.; Badel, E.; Herbette, S.; Delzon, S.; Choat, B.; Jansen, S. Methods for measuring plant vulnerability to cavi-tation: A critical review. J. Exp. Bot. 2013, 64, 4779–4791; [Google Scholar] [CrossRef] [Green Version]
- Melcher, P.; Warchocki, S. The impact of xylem wounding on the measure of stem hydraulic resistance. In Botanical Society of America; Providence, RI, USA, 2010. [Google Scholar]
- Melcher, P.J.; Michele Holbrook, N.; Burns, M.J.; Zwieniecki, M.A.; Cobb, A.R.; Brodribb, T.J.; Choat, B.; Sack, L. Meas-urements of stem xylem hydraulic conductivity in the laboratory and field. Methods Ecol. Evol. 2012, 3, 685–694; [Google Scholar] [CrossRef]
- Chave, J.; Muller-Landau, H.C.; Baker, T.R.; Easdale, T.A.; ter Steege, H.; Webb, C.O. Regional and phylogenetic variation of wood density across 2456 Neotropical tree species. Ecol. Appl. 2006, 16, 2356–2367; [Google Scholar] [CrossRef] [Green Version]
- William, R.L.A.; Klein, T.; Bartlett, M.; Sack, L.; Adam, F.A.P.; Choat, B.; Jansen, S. Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought-induced tree mortality across the globe. Proc. Natl. Acad. Sci. USA 2016, 113, 5024–5029; [Google Scholar] [CrossRef] [Green Version]
- Tyree, M.T.; Zimmermann, M.H. Xylem Structure and the Ascent of Sap, 2nd ed.; Springer: Berlin, Heidelberg, Germany, 2002; p. 283. [Google Scholar]
- Lens, F.; Sperry, J.S.; Christman, M.A.; Choat, B.; Rabaey, D.; Jansen, S. Testing hypotheses that link wood anatomy to cavitation resistance and hydraulic conductivity in the genus Acer. New Phytol. 2011, 190, 709–723; [Google Scholar] [CrossRef] [Green Version]
- Duursma, R.; Choat, B. fitplc—An R package to fit hydraulic vulnerability curves. J. Plant Hydraul. 2017, 4, 002; [Google Scholar] [CrossRef]
- Tyree, M.T.; Sperry, J.S. Vulnerability of xylem to cavitation and embolism. Annu. Rev. Plant Physiol. Plant Molec. Biol. 1989, 40, 19–36; [Google Scholar] [CrossRef]
- Maherali, H.; DeLucia, E.H. Xylem conductivity and vulnerability to cavitation of ponderosa pine growing in contrasting climates. Tree Physiol. 2000, 20, 859–867; [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lens, F.; Espino, S.; Karimi, Z.; Klepsch, M.; Schenk, H.J.; Schmitt, M.; Schuldt, B.; Jansen, S. Intervessel pit membrane thickness as a key determinant of embolism resistance in angiosperm xylem. IAWA J. 2016, 37, 152–171; [Google Scholar] [CrossRef] [Green Version]
- Hacke, U.G.; Sperry, J.S.; Pockman, W.T.; Davis, S.D.; McCulloh, K.A. Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 2001, 126, 457–461; [Google Scholar] [CrossRef] [PubMed]
- Janssen, T.A.J.; Hölttä, T.; Fleischer, K.; Naudts, K.; Dolman, H. Wood allocation trade-offs between fiber wall, fiber lumen, and axial parenchyma drive drought resistance in neotropical trees. Plant Cell Environ. 2020, 43, 965–980; [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeber, S.; Leuschner, C.; Köhler, L.; Arias-Aguilar, D.; Schuldt, B. The importance of hydraulic conductivity and wood density to growth performance in eight tree species from a tropical semi-dry climate. For. Ecol. Manag. 2014, 330, 126–136; [Google Scholar] [CrossRef]
- Schumann, K.; Leuschner, C.; Schuldt, B. Xylem hydraulic safety and efficiency in relation to leaf and wood traits in three temperate Acer species differing in habitat preferences. Trees 2019, 33, 1475–1490; [Google Scholar] [CrossRef]
- Schmitz, N.; Verheyden, A.; Beeckman, H.; Kairo, J.G.; Koedam, N. Influence of a salinity gradient on the vessel characters of the mangrove species Rhizophora mucronata. Ann. Bot. 2006, 98, 1321–1330; [Google Scholar] [CrossRef]
- Liu, H.; Ye, Q.; Gleason, S.M.; He, P.; Yin, D. Weak tradeoff between xylem hydraulic efficiency and safety: Climatic sea-sonality matters. New Phytol. 2021, 229, 1440–1452; [Google Scholar] [CrossRef]
- Gleason, S.M.; Westoby, M.; Jansen, S.; Choat, B.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Hacke, U.G.; Jacobsen, A.L.; Johnson, D.M.; et al. On research priorities to advance understanding of the safety-efficiency tradeoff in xylem. New Phytol. 2016, 211, 1156–1158; [Google Scholar] [CrossRef] [Green Version]
- Lovelock, C.E.; Krauss, K.W.; Osland, M.J.; Reef, R.; Ball, M.C. The physiology of mangrove trees with changing climate. In Tropical Tree Physiology; Goldstein, G., Santiago, L.S., Eds.; Springer: Cham, Switzerland, 2016; pp. 149–179. [Google Scholar]
- Feller, I.C.; Lovelock, C.E.; Berger, U.; McKee, K.L.; Joye, S.B.; Ball, M.C. Biocomplexity in Mangrove Ecosystems. Annu. Rev. Mar. Sci. 2010, 2, 395–417; [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, E.M.R.; Koedam, N.; Beeckman, H.; Schmitz, N. A safe hydraulic architecture as wood anatomical explanation for the difference in distribution of the mangroves Avicennia and Rhizophora. Funct. Ecol. 2009, 23, 649–657; [Google Scholar] [CrossRef]
- Schmitz, N.; Robert, E.M.R.; Verheyden, A.; Kairo, J.G.; Beeckman, H.; Koedam, N. A patchy growth via successive and simulta-neous cambia: Key to success of the most widespread mangrove species Avicennia marina? Ann. Bot. 2007, 101, 49–58; [Google Scholar] [CrossRef]
- Robert, E.M.R.; Jambia, A.H.; Schmitz, N.; De Ryck, D.J.R.; De Mey, J.; Kairo, J.G.; Dahdouh-Guebas, F.; Beeckman, H.; Koedam, N. How to catch the patch? A dendrometer study of the radial increment through successive cambia in the mangrove Avicennia. Ann. Bot. 2014, 113, 741–752; [Google Scholar] [CrossRef]
- Carlquist, S. Successive cambia revisited: Ontogeny, histology, diversity, and functional significance. J. Torrey Bot. Soc. 2007, 134, 301–332; [Google Scholar] [CrossRef]
- Robert, E.M.R.; Schmitz, N.; Boeren, I.; Driessens, T.; Herremans, K.; De Mey, J.; Van de Casteele, E.; Beeckman, H.; Koedam, N. Successive cambia: A developmental oddity or an adaptive structure? PLoS ONE 2011, 6, e16558; [Google Scholar] [CrossRef] [Green Version]
- Santini, N.S.; Schmitz, N.; Lovelock, C.E. Variation in wood density and anatomy in a widespread mangrove species. Trees 2012, 26, 1555–1563; [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Wheeler, J.K.; Castro, L. Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiol. 2006, 26, 689–701; [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S. Functional and ecological xylem anatomy. Perspect. Plant Ecol. Evol. Syst. 2001, 4, 97–115. [Google Scholar] [CrossRef] [Green Version]
- Choat, B.; Cobb, A.R.; Jansen, S. Structure and function of bordered pits: New discoveries and impacts on whole-plant hydraulic function. New Phytol. 2008, 177, 608–626; [Google Scholar] [CrossRef] [PubMed]
- Asbridge, E.; Lucas, R.; Accad, A.; Dowling, R. Mangrove response to environmental changes predicted under varying climates: Case studies from Australia. Curr. For. Rep. 2015, 1, 178–194; [Google Scholar] [CrossRef] [Green Version]
- Santiago, L.S.; Goldstein, G.; Meinzer, F.C.; Fisher, J.B.; Machado, K.; Woodruff, D.; Jones, T. Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees. Oecologia 2004, 140, 543–550; [Google Scholar] [CrossRef] [PubMed]
- Fazlioglu, F.; Wan, J.S.H.; Chen, L. Latitudinal shifts in mangrove species worldwide: Evidence from historical occurrence records. Hydrobiologia 2020, 847, 4111–4123; [Google Scholar] [CrossRef]
- Cavanaugh, K.C.; Kellner, J.R.; Forde, A.J.; Gruner, D.S.; Parker, J.D.; Rodriguez, W.; Feller, I.C. Poleward expansion of mangroves is a threshold response to decreased frequency of extreme cold events. Proc. Natl. Acad. Sci. USA 2014, 111, 723–727; [Google Scholar] [CrossRef] [Green Version]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52–58; [Google Scholar] [CrossRef]
- Feher, L.C.; Osland, M.J.; Griffith, K.T.; Grace, J.B.; Howard, R.J.; Stagg, C.L.; Enwright, N.M.; Krauss, K.W.; Gabler, C.A.; Day, R.H.; et al. Linear and nonlinear effects of temperature and precipitation on ecosystem properties in tidal saline wetlands. Ecosphere 2017, 8, e01956; [Google Scholar] [CrossRef]
- Osland, M.J.; Feher, L.C.; Griffith, K.T.; Cavanaugh, K.C.; Enwright, N.M.; Day, R.H.; Stagg, C.L.; Krauss, K.W.; Howard, R.J.; Grace, J.B.; et al. Climatic controls on the global distribution, abundance, and species richness of mangrove forests. Ecol. Monogr. 2017, 87, 341–359; [Google Scholar] [CrossRef] [Green Version]
- Gabler, C.A.; Osland, M.J.; Grace, J.B.; Stagg, C.L.; Day, R.H.; Hartley, S.B.; Enwright, N.M.; From, A.S.; McCoy, M.L.; McLeod, J.L. Macroclimatic change expected to transform coastal wetland ecosystems this century. Nat. Clim. Chang. 2017, 7, 142–147; [Google Scholar] [CrossRef]
- Steppe, K.; Vandegehuchte, M.W.; Van de Wal, B.A.E.; Hoste, P.; Guyot, A.; Lovelock, C.E.; Lockington, D.A. Direct uptake of canopy rainwater causes turgor-driven growth spurts in the mangrove Avicennia marina. Tree Physiol. 2018, 38, 979–991; [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Coordinates | MAT | Pmin | PS | MAP | AI | PET |
|---|---|---|---|---|---|---|---|
| Marlow | 33° 32′ 54″ S, 151° 11′ 54″ E | 17.10 | 56 | 28.99 | 864.57 | 1.55 | 1338.85 |
| MBRS | 27° 24′ 25″ S, 153° 26′ 18″ E | 19.88 | 43 | 35.09 | 1200.01 | 1.24 | 1491.93 |
| DRO | 16° 5′ 60″ S, 145° 27′ 59″ E | 24.69 | 28 | 98.08 | 2920.08 | 0.54 | 1567.29 |
| Species | Site | P12 | P50 | P88 | HSM50 | HSM88 | Ψpd | Ψmd-leaf | Ψmd-stem |
|---|---|---|---|---|---|---|---|---|---|
| Avicennia marina | Marlow | 3.86[3.29, 4.56] | 5.87[5.46, 6.24] | 7.75[7.11, 8.37] | 1.45 | 3.33 | 3.14 ± 0.05a | 5.32 ± 0.13a | 4.42 ± 0.04a |
| MBRS | 5.13[4.59, 5.53] | 7.67[7.39, 7.88] | 10.02[9.57, 10.56] | 3.25 | 5.6 | 2.20 ± 0.22b | 5.19 ± 0.05a | 4.42 ± 0.28a | |
| DRO | 6.20[4.64, 8.15] | 8.52[7.98, 9.40] | 10.51[9.41, 14.55] | 4.93 | 6.92 | 3.24 ± 0.15a | 4.72 ± 0.15b | 3.59 ± 0.11b | |
| Aegiceras corniculatum | Marlow | 2.06[0.90, 4.59] | 4.68[3.57, 6.01] | 8.08[6.54, 13.15] | 1.08 | 4.48 | 2.90 ± 0.12a | 4.40 ± 0.13a | 3.60 ± 0.05a |
| MBRS | 1.90[1.04, 2.92] | 5.51[4.97, 6.31] | 11.13[8.82, 20.05] | 3.05 | 8.67 | 1.97 ± 0.06b | 3.25 ± 0.07c | 2.46 ± 0.09b | |
| DRO | 4.92[3.71, 5.64] | 7.90[7.51, 9.34] | 10.81[9.87, 17.01] | 5.18 | 8.09 | 2.14 ± 0.05b | 3.79 ± 0.06b | 2.72 ± 0.05b |
| Species | Site | KLx104 | KS | MVL | SWC | DH | VD | WD | VWR | HVx104 |
|---|---|---|---|---|---|---|---|---|---|---|
| Avicennia marina | Marlow | 2.83 ± 0.51a | 0.75 ± 0.07b | 23.00 ± 6.55a | 0.96 ± 0.05a | 40.41 ± 1.97a | 74.83 ± 8.82a | 0.63 ± 0.02a | 0.059 ± 0.0014b | 4.40 ± 0.59a |
| MBRS | 2.56 ± 0.24a | 0.78 ± 0.07ab | 27.27 ± 4.57a | 0.95 ± 0.04a | 37.11 ± 1.22a | 85.08 ± 4.62a | 0.64 ± 0.01a | 0.067 ± 0.0061b | 4.08 ± 0.63a | |
| DRO | 3.84 ± 0.52a | 1.19 ± 0.15a | 30.00 ± 2.10a | 1.01 ± 0.03a | 35.20 ± 1.00a | 86.53 ± 6.82a | 0.60 ± 0.01a | 0.098 ± 0.0071a | 3.33 ± 0.41a | |
| Aegiceras corniculatum | Marlow | 1.68 ± 0.20b | 0.63 ± 0.09b | 22.53 ± 1.37a | 0.78 ± 0.02a | 21.78 ± 0.81a | 310.54 ± 8.02b | 0.68 ± 0.01a | 0.037 ± 0.0021b | 2.79 ± 0.35b |
| MBRS | 1.66 ± 0.37b | 0.58 ± 0.04b | 25.93 ± 4.63a | 0.82 ± 0.02a | 20.75 ± 0.40a | 318.12 ± 19.30b | 0.65 ± 0.01a | 0.035 ± 0.0010b | 2.72 ± 0.28b | |
| DRO | 4.02 ± 0.73a | 0.99 ± 0.05a | 27.77 ± 4.77a | 0.86 ± 0.04a | 21.80 ± 0.41a | 402.90 ± 22.08a | 0.59 ± 0.01b | 0.047 ± 0.0027a | 4.64 ± 0.44a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, X.; Choat, B.; Zhang, Y.-J.; Guan, X.-Y.; Shi, W.; Cao, K.-F. Variation in Xylem Hydraulic Structure and Function of Two Mangrove Species across a Latitudinal Gradient in Eastern Australia. Water 2021, 13, 850. https://doi.org/10.3390/w13060850
Jiang X, Choat B, Zhang Y-J, Guan X-Y, Shi W, Cao K-F. Variation in Xylem Hydraulic Structure and Function of Two Mangrove Species across a Latitudinal Gradient in Eastern Australia. Water. 2021; 13(6):850. https://doi.org/10.3390/w13060850
Chicago/Turabian StyleJiang, Xin, Brendan Choat, Yong-Jiang Zhang, Xin-Yi Guan, Wen Shi, and Kun-Fang Cao. 2021. "Variation in Xylem Hydraulic Structure and Function of Two Mangrove Species across a Latitudinal Gradient in Eastern Australia" Water 13, no. 6: 850. https://doi.org/10.3390/w13060850
APA StyleJiang, X., Choat, B., Zhang, Y. -J., Guan, X. -Y., Shi, W., & Cao, K. -F. (2021). Variation in Xylem Hydraulic Structure and Function of Two Mangrove Species across a Latitudinal Gradient in Eastern Australia. Water, 13(6), 850. https://doi.org/10.3390/w13060850