
Genetic Diversity of Stellate Sturgeon in the Lower Danube River: The Impact of Habitat Contraction upon a Critically Endangered Population

 ,
, 
 , , and
, , and 
Abstract
:1. Introduction
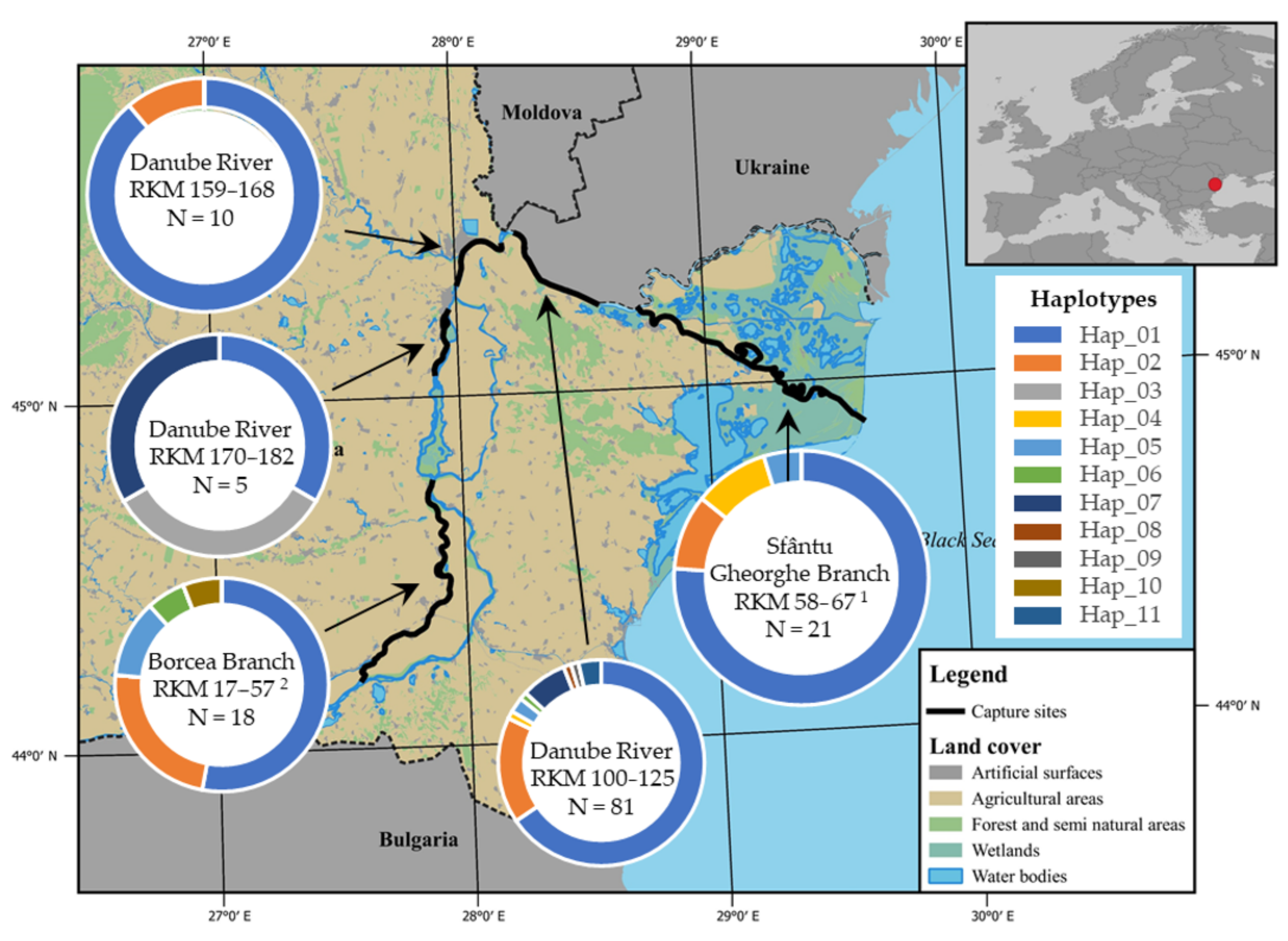
2. Materials and Methods
3. Results
4. Discussion
4.1. Genetic Variability within Cohorts of Adult Stellate Sturgeons Migrating for Spawning in the Lower Danube River and YOY on Their Downstream Migration
4.2. Genetic Population Structure of Autumn and Spring Migrant Adults
4.3. Implications for the Management of Species Conservation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qiwei, W. Acipenser stellatus. IUCN Red List Threat. Species 2010. [Google Scholar] [CrossRef]
- Bacalbasa-Dobrovici, N.; Patriche, N. Environmental studies and recovery actions for sturgeon in the Lower Danube River system. J. Appl. Ichthyol. 1999, 15, 114–115. [Google Scholar] [CrossRef]
- Vassilev, M. Lower Danube—The last refuge for surviving of sturgeon fishes in the Black Sea Region. In Proceedings of the Water Observation and Information System for Decision Support, Ohrid, Macedonia, 23–13 May 2006. [Google Scholar]
- Vecsei, P.; Peterson, D.; Suciu, R.; Artyukhin, E. Threatened fishes of the world, Acipenser stellatus, Pallas, 1771 (Acipenseridae). Environ. Biol. Fishes 2007, 78, 211–212. [Google Scholar] [CrossRef]
- Ceapa, C.; Williot, P.; Bacalbasa-Dobrovici, N. Present state and perspectives of stellate sturgeon brood fish in the Romanian part of the Danube. Int. Rev. Hydrobiol. A J. Cover. All Asp. Limnol. Mar. Biol. 2002, 87, 507–513. [Google Scholar] [CrossRef]
- Paraschiv, M.; Suciu, R.; Suciu, M. Present state of sturgeon stocks in the Lower Danube River, Romania. In Proceedings of the 36th International Conference of IAD, Austrian Committee Danube Research, Klosterneuburg, Austria, 4–8 September 2006; pp. 4–8. [Google Scholar]
- Smederevac-Lalić, M.; Jarić, I.; Višnjić-Jeftić, Ž.; Skorić, S.; Cvijanović, S.; Gačić, Z.; Lenhardt, M. Management approaches and aquaculture of sturgeons in the Lower Danube region countries. J. Appl. Ichthyol. 2011, 27, 94–100. [Google Scholar] [CrossRef]
- Norouzi, M.; Pourkazemi, M.; Fatemi, M. Application of microsatellite markers to study the genetic structure of stellate sturgeon populations (Acipenser stellatus Pallas, 1771) in the south Caspian Sea. Iran. J. Fish. Sci. 2009, 8, 73–84. [Google Scholar]
- Ryabova, G.D.; Klimonov, R.P.; Afanas’ev, V.O.; Rubtsova, K.I.; Dovgopol, G.A.; Khodorevskaya, G.F. A comparison of the spawning migration, genetic and biological parameters of stellate sturgeon from the Volga population in 1985 and 1996. Russ. J. Genet. 2006, 42, 1180–1188. [Google Scholar] [CrossRef]
- Chebanov, G.I.; Karnaukhov, M.S.; Galich, E.V.; Chmir, Y.N. Hatchery stock enhancement and conservation of sturgeon, with an emphasis on the Azov Sea populations. J. Appl. Ichthyol. 2002, 18, 463–469. [Google Scholar] [CrossRef]
- Secor, D.H.; Arefjev, V.; Nikolaev, A.; Sharov, A. Restoration of sturgeons: Lessons from the Caspian Sea sturgeon ranching programme. Fish Fish. 2000, 1, 215–230. [Google Scholar] [CrossRef]
- Ruban, G.I.; Khodorevskaya, R.P.; Shatunovskiy, M.I. Long-Term Dynamics of Sturgeon Distribution in the Northern Part of the Caspian Sea (Review). Inland Water Biol. 2019, 12, 443–451. [Google Scholar] [CrossRef]
- Ruban, G.; Khodorevskaya, R.; Shatunovskii, M. Factors influencing the natural reproduction decline in the beluga (Huso huso, Linnaeus, 1758), Russian sturgeon (Acipenser gueldenstaedtii, Brandt & Ratzeburg, 1833), and stellate sturgeon (A. stellatus, Pallas, 1771) of the Volga-Caspian basin: A review. J. Appl. Ichthyol. 2019, 35, 387–395. [Google Scholar] [CrossRef] [Green Version]
- Doukakis, P.; Birstein, V.J.; De Salle, R. Intraspecific structure within three caviar-producing sturgeons (Acipenser gueldenstaedtii, A. stellatus, and Huso huso) based on mitochondrial DNA analysis. J. Appl. Ichthyol. 2005, 21, 457–460. [Google Scholar] [CrossRef]
- Doukakis, P.; Birstein, V.J.; Ruban, G.I.; DeSalle, R. Molecular genetic analysis among subspecies of two Eurasian sturgeon species, Acipenser baerii and A. stellatus. Mol. Ecol. 1999, 8, S117–S127. [Google Scholar] [CrossRef] [PubMed]
- Dudu, A.; Georgescu, S.E.; Luca, C.; Suciu, R.; Costache, M. Microsatelitte DNA variation in the black sea stellate sturgeon, Acipenser stellatus. Sci. Pap. Anim. Sci. Biotechnol. 2008, 41, 78–82. [Google Scholar]
- Dudu, A.; Georgescu, S.E.; Costache, M. Molecular analysis of phylogeographic subspecies in three Ponto-Caspian sturgeon species. Genet. Mol. Biol. 2014, 37, 587–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holostenco, D.; Onără, D.F.; Suciu, R.; Honț, S.; Paraschiv, M. Distribution and genetic diversity of sturgeons feeding in the marine area of the Danube Delta Biosphere Reserve. Sci. Ann. Danub. Delta Inst. 2013, 19, 25–34. [Google Scholar] [CrossRef]
- Taggart, J.B.; Hynes, R.A.; Prodöuhl, P.A.; Ferguson, A. A simplified protocol for routine total DNA isolation from salmonid fishes. J. Fish Biol. 1992, 40, 963–965. [Google Scholar] [CrossRef]
- Onara, D.; Holostenco, D.; Paraschiv, M.; Suciu, R. Preliminary genetic variability of Lower Danube River young of the year (YOY) beluga sturgeon Huso huso (Linnaeus, 1758) using mt DNA markers. J. Appl. Ichthyol. 2014, 30, 1286–1289. [Google Scholar] [CrossRef]
- Hall, T.; Biosciences, I.; Carlsbad, C. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Burland, T.G. DNASTAR’s Lasergene sequence analysis software. In Bioinformatics Methods and Protocols; Misener, S., Krawetz, S.A., Eds.; Humana Press: Totowa, NJ, USA, 2000; pp. 71–91. [Google Scholar]
- Delignette-Muller, M.; Dutang, C. Fitdistrplus: An R package for fitting distributions. J. Stat. Softw. 2015, 64, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Kooperberg, C.; Stone, C. Logspline density estimation for censored data. J. Comput. Graph. Stat. 1992, 1, 301–328. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. Paleontological statistics software package for education and data analysis. Paleontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Waldman, J.R.; Hart, J.T.; Wirgin, I.I. Stock composition of the New York Bight Atlantic sturgeon fishery based on analysis of mitochondrial DNA. Trans. Am. Fish. Soc. 1996, 125, 364–371. [Google Scholar] [CrossRef]
- Nazari, S.; Pourkazemi, M.R.; Khoshkholgh, M.; Azizzade, L. Population structure and variation in Persian sturgeon (Acipenser percicus) from the Caspian Sea as determind from mitochondrial DNA sequences of the control region. Prog. Biol. Sci. 2013, 3, 67–80. [Google Scholar]
- Pourkazemi, M.; Khoshkholgh, M.; Nazari, S.; Azizzadeh Pormehr, L. Genetic relationships among collections of the Persian sturgeon, Acipenser percicus, in the south Caspian Sea detected by mitochondrial DNA Restriction fragment length polymorphisms. Casp. J. Environ. Sci. 2012, 10, 215–226. [Google Scholar]
- Hensel, K.; Holcík, J. Past and current status of sturgeons in the upper and middle Danube River. In Sturgeon Biodiversity and Conservation; Birstein, V.J., Waldman, J.R., Bemis, W.E., Eds.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1997; pp. 185–200. [Google Scholar]
- Iani, I.M.; Paraschiv, M.; Honţ, Ş.; Holostenco, D.; Toṧić, K.; Taflan, E.; Suciu, R.; Georgescu, P.L.; Iticescu, C.; Țopa, C.; et al. Glimpse of a Better Future for the Danube’s Flagship Species—First Return of Stocked Sturgeons into the River. Sci. Ann. Danub. Delta Inst. 2019, 24, 31–40. [Google Scholar]
- Onără, D.F.; Holostenco, D.; Suciu, R. Management applications of genetic structure of anadromous sturgeon populations in the Lower Danube River (LDR), Romania. Sci. Ann. Danub. Delta Inst. 2013, 19, 129–138. [Google Scholar] [CrossRef]
- Stabile, J.; Waldman, J.R.; Parauka, F.; Wirgin, I. Stock structure and homing fidelity in Gulf of Mexico sturgeon (Acipenser oxyrinchus desotoi) based on restriction fragment length polymorphism and sequence analyses of mitochondrial DNA. Genetics 1996, 144, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Somero, G.N. Thermal physiology and vertical zonation of intertidal animals: Optima, limits, and costs of living. Integr. Comp. Biol. 2002, 42, 780–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruch, R.M.; Binkowski, F.P. Spawning behavior of lake sturgeon (Acipenser fulvescens). J. Appl. Ichthyol. 2002, 18, 570–579. [Google Scholar] [CrossRef] [Green Version]
- Deguo, Y.; Qiwei, W.; Xihua, C.; Jianyi, L.; Yongjiu, Z.; Kai, W. Hydrological status of the spawning ground of Acipenser sinensis underneath the Gezhouba Dam and its relationship with the spawning runs. Acta Ecol. Sin. 2007, 27, 862–868. [Google Scholar] [CrossRef]
- He, T.; Deng, Y.; Tuo, Y.; Yang, Y.; Liang, N. Impact of the Dam Construction on the Downstream Thermal Conditions of the Yangtze River. Int. J. Environ. Res. Public Health 2020, 17, 2973. [Google Scholar] [CrossRef]
- Wan, Q.H.; Fang, S.G. The influence of the Gezhouba Reservoir on the genetic diversity of the Chinese Sturgeon, Acipenser sinensis. J. Freshw. Ecol. 2003, 18, 135–139. [Google Scholar] [CrossRef]
- Cooke, S.J.; Cech, J.J.; Glassman, J.; Simard, D.M.; Louttit, S.; Lennox, R.J.; O’Connor, C.M. Water resource development and sturgeon (Acipenseridae): State of the science and research gaps related to fish passage, entrainment, impingement and behavioural guidance. Rev. Fish Biol. Fish. 2020, 30, 219–244. [Google Scholar] [CrossRef]
- Nyeste, K.; Somogyi, D.; Sallai, Z.; Antal, L. Adatok a tokfélék (Acipenseridae) Kárpát-medencei recens előfordulásairól= Recent occurrence data of sturgeons (Acipenseridae) in the Carpathian Basin. Pisces Hung. 2020, 14, 107–114. [Google Scholar]
- Suciu, R.; Guti, G. Have sturgeons a future in the Danube River? In Proceedings of the 39th IAD Conference Living Danube, Szentendre, Hungary, 21–24 August 2012; pp. 19–30. [Google Scholar]
- Grunwald, C.; Maceda, L.; Waldman, J.; Stabile, J.; Wirgin, I. Conservation of Atlantic sturgeon Acipenser oxyrinchus oxyrinchus: Delineation of stock structure and distinct population segments. Conserv. Genet. 2008, 9, 1111–1124. [Google Scholar] [CrossRef]
- Grunwald, C.; Stabile, J.; Waldman, J.R.; Gross, R.; Wirgin, I. Population genetics of shortnose sturgeon Acipenser brevirostrum based on mitochondrial DNA control region sequences. Mol. Ecol. 2002, 11, 1885–1898. [Google Scholar] [CrossRef]
- Wirgin, I.; Grunwald, C.; Carlson, E.; Stabile, J.; Peterson, D.L.; Waldman, J. Range-wide population structure of shortnose sturgeon Acipenser brevirostrum based on sequence analysis of the mitochondrial DNA control region. Estuaries 2005, 28, 406–421. [Google Scholar] [CrossRef]
- Šarhanová, P.; Pfanzelt, S.; Brandt, R.; Himmelbach, A.; Blattner, F.R. SSR-seq: Genotyping of microsatellites using next-generation sequencing reveals higher level of polymorphism as compared to traditional fragment size scoring. Ecol. Evol. 2018, 8, 10817–10833. [Google Scholar] [CrossRef]
- Tibihika, P.D.; Curto, M.; Dornstauder-Schrammel, E.; Winter, S.; Alemayehu, H.; Waidbacher, E.; Meimberg, H. Application of microsatellite genotyping by sequencing (SSR-GBS) to measure genetic diversity of the East African Oreochromis niloticus. Conserv. Genet. 2019, 20, 357–372. [Google Scholar] [CrossRef] [Green Version]
- Curto, M.; Winter, S.; Seiter, A.; Schmid, L.; Scheicher, K.; Barthel, L.M.; Meimberg, H. Application of a SSR-GBS marker system on investigation of European Hedgehog species and their hybrid zone dynamics. Ecol. Evol. 2019, 9, 2814–2832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Káldy, J.; Mozsár, A.; Fazekas, G.; Farkas, M.; Fazekas, D.L.; Fazekas, G.L.; Goda, K.; Gyöngy, Z.; Kovács, B.; Semmens, K.; et al. Hybridization of Russian Sturgeon (Acipenser gueldenstaedtii, Brandt and Ratzeberg, 1833) and American Paddlefish (Polyodon spathula, Walbaum 1792) and evaluation of their progeny. Genes 2020, 11, 753. [Google Scholar] [CrossRef] [PubMed]
- Henry, P.; Russello, M.A. Obtaining high-quality DNA from elusive small mammals using low-tech hair snares. Eur. J. Wildl. Res. 2011, 57, 429–435. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Season | 1999 | 2000 | 2001 | 2002 * | 2003 * | 2004 * | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Upstream migration (adult) | ||||||||||||||
| Spring a | 0 | 0 | 0 | 0 | 13 3 | 5 3 | 0 | N.A. | 22 4 | N.A. | N.A. | 11 5 | 8 6 | 78 |
| Autumn b | 13 1 | 6 2 | 0 | 0 | 0 | 0 | 0 | N.A. | N.A. | N.A. | N.A. | N.A. | N.A. | |
| Downstream migration (YOY) | ||||||||||||||
| Summer c | 0 | 15 | 0 | 0 | 0 | 0 | 11 | 0 | 0 | 8 | 6 | 16 | 0 | 56 |
| Haplotype Name | Haplotype Definition | Haplotype Composition | ||
|---|---|---|---|---|
| HaeIII | BslI | BsrI | ||
| Hap_01 | AAA | A | A | A |
| Hap_02 | ABA | A | B | A |
| Hap_03 | BAA | B | A | A |
| Hap_04 | CAA | C | A | A |
| Hap_05 | AAB | A | A | B |
| Hap_06 | ACA | A | C | A |
| Hap_07 | DAA | D | A | A |
| Hap_08 | BBA | B | B | A |
| Hap_09 | AAC | A | A | C |
| Hap_10 | DBA | D | B | A |
| Hap_11 | EAA | E | A | A |
| Haplotype | Adults | YOY | ||||
|---|---|---|---|---|---|---|
| Spring | Autumn | Summer | ||||
| Female | Male | Female | Male | Unknown | ||
| Hap_01 | 16 | 25 | 0 | 4 | 11 | 33 |
| Hap_02 | 2 | 5 | 0 | 1 | 1 | 12 |
| Hap_03 | 0 | 0 | 0 | 0 | 1 | 0 |
| Hap_04 | 0 | 1 | 1 | 0 | 0 | 1 |
| Hap_05 | 1 | 2 | 0 | 0 | 0 | 2 |
| Hap_06 | 1 | 1 | 0 | 0 | 0 | 0 |
| Hap_07 | 1 | 2 | 0 | 0 | 0 | 4 |
| Hap_08 | 0 | 1 | 0 | 0 | 0 | 0 |
| Hap_09 | 0 | 0 | 0 | 0 | 0 | 1 |
| Hap_10 | 0 | 1 | 0 | 0 | 0 | 0 |
| Hap_11 | 0 | 0 | 0 | 0 | 0 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holostenco, D.N.; Ciorpac, M.; Taflan, E.; Tošić, K.; Paraschiv, M.; Iani, M.; Honț, Ș.; Suciu, R.; Rîșnoveanu, G. Genetic Diversity of Stellate Sturgeon in the Lower Danube River: The Impact of Habitat Contraction upon a Critically Endangered Population. Water 2021, 13, 1115. https://doi.org/10.3390/w13081115
Holostenco DN, Ciorpac M, Taflan E, Tošić K, Paraschiv M, Iani M, Honț Ș, Suciu R, Rîșnoveanu G. Genetic Diversity of Stellate Sturgeon in the Lower Danube River: The Impact of Habitat Contraction upon a Critically Endangered Population. Water. 2021; 13(8):1115. https://doi.org/10.3390/w13081115
Chicago/Turabian StyleHolostenco, Daniela Nicoleta, Mitică Ciorpac, Elena Taflan, Katarina Tošić, Marian Paraschiv, Marian Iani, Ștefan Honț, Radu Suciu, and Geta Rîșnoveanu. 2021. "Genetic Diversity of Stellate Sturgeon in the Lower Danube River: The Impact of Habitat Contraction upon a Critically Endangered Population" Water 13, no. 8: 1115. https://doi.org/10.3390/w13081115
APA StyleHolostenco, D. N., Ciorpac, M., Taflan, E., Tošić, K., Paraschiv, M., Iani, M., Honț, Ș., Suciu, R., & Rîșnoveanu, G. (2021). Genetic Diversity of Stellate Sturgeon in the Lower Danube River: The Impact of Habitat Contraction upon a Critically Endangered Population. Water, 13(8), 1115. https://doi.org/10.3390/w13081115