
A First Glimpse on Cold-Adapted PCB-Oxidizing Bacteria in Edmonson Point Lakes (Northern Victoria Land, Antarctica)

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Area
2.2. Sampling and Preliminary Treatment of Samples
2.3. Bacterial Isolation and Characterization
2.3.1. Set-Up of Bacterial Cultures
2.3.2. Bacterial Growth in the Presence of Aroclor 1242
2.3.3. 16S rRNA Gene Amplification
2.3.4. Sequencing and Analysis of 16S rRNA Genes
2.3.5. Screening for the bphA Gene
3. Results
3.1. Bacterial Isolation
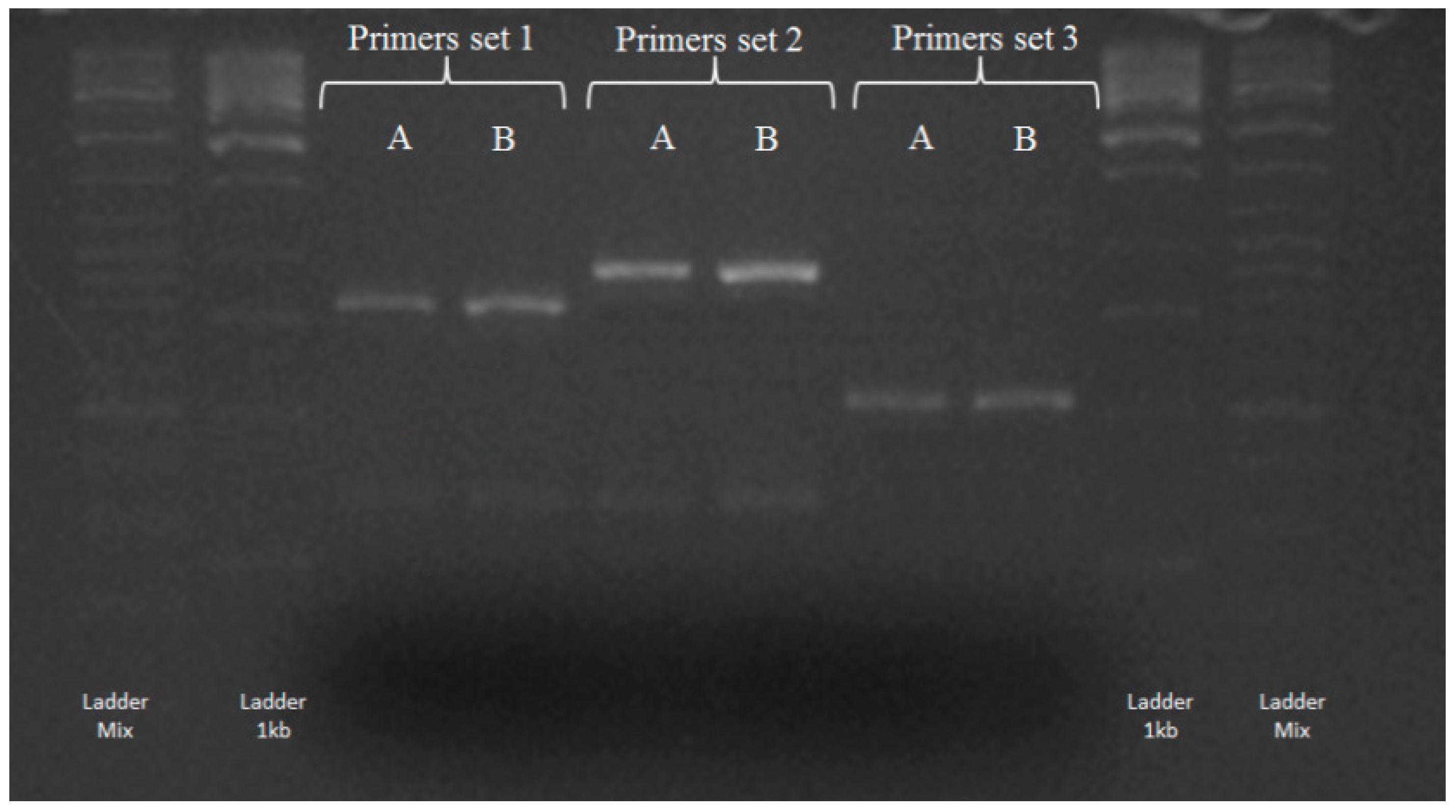
3.2. Screening for bphA Gene
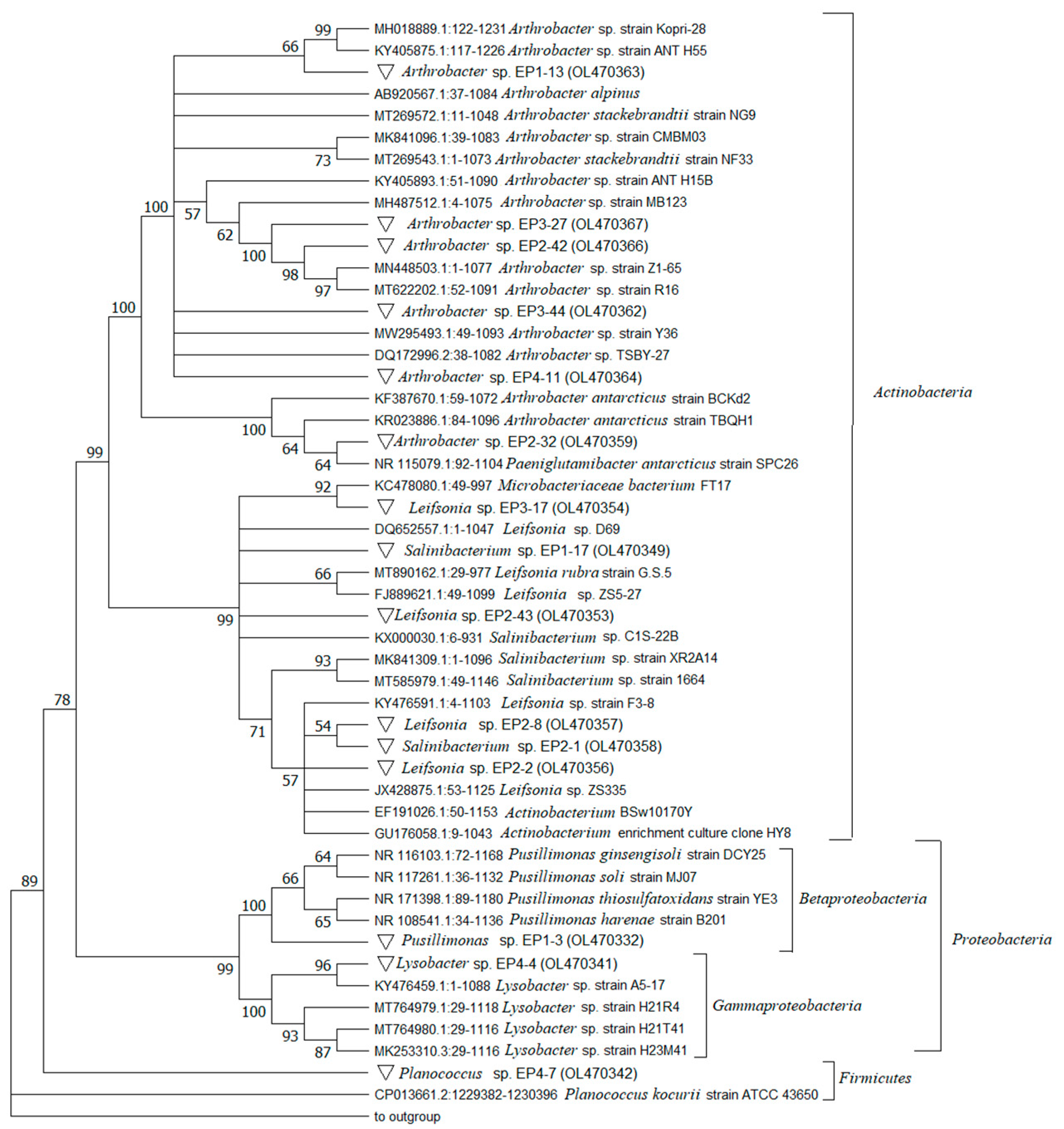
3.3. Bacterial Identification
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lo Giudice, A.; Caruso, G.; Rizzo, C.; Papale, M.; Azzaro, M. Bacterial communities versus anthropogenic disturbances in the Antarctic coastal marine environment. Environ. Sustain. 2019, 2, 297–310. [Google Scholar] [CrossRef]
- Gambaro, A.; Manodori, L.; Zangrando, R.; Cincinelli, A.; Capodaglio, G.; Cescon, P. Atmospheric PCB concentrations at Terra Nova Bay, Antarctica. Environ. Sci. Technol. 2005, 39, 9406–9411. [Google Scholar] [CrossRef]
- Bhardwaj, L.; Chauhan, A.; Ranjan, A.; Jindal, T. Persistent organic pollutants in biotic and abiotic components of Antarctic pristine environment. Earth Syst. Environ. 2018, 2, 35–54. [Google Scholar] [CrossRef]
- Fuoco, R.; Giannarelli, S.; Wei, Y.; Ceccarini, A.; Abete, C.; Francesconi, S.; Termine, M. Persistent organic pollutants (POPs) at Ross sea (Antarctica). Microchem. J. 2009, 92, 44–48. [Google Scholar] [CrossRef]
- Montone, R.C.; Taniguchi, S.; Sericano, J.; Weber, R.R.; Lara, W.H. Determination of biphenyls in Antarctic macroalgae Desmaretia sp. Sci. Total Environ. 2001, 277, 181–186. [Google Scholar] [CrossRef]
- Montone, R.C.; Taniguchi, S.; Weber, R.R. Polychlorinated biphenyls in marine sediments of Admiralty Bay, King George Island, Antarctica. Mar. Pollut. Bull. 2001, 42, 611–614. [Google Scholar] [CrossRef]
- Focardi, S.; Lari, L.; Marsili, L. PCB congeners, DDTs and hexachlorobenzene in Antarctic fish from Terra Nova Bay (Ross Sea). Antarct. Sci. 1992, 4, 151–154. [Google Scholar] [CrossRef]
- Focardi, S.; Bargagli, R.; Corsolini, S. Isomer-specific analysis and toxic potential evaluation of polychlorinated biphenyls in Antarctic fish, seabirds and Weddell seals from Terra Nova Bay (Ross Sea). Antarct. Sci. 1995, 7, 31–35. [Google Scholar] [CrossRef]
- Corsolini, S.; Ademollo, N.; Romeo, T.; Olmastroni, S.; Focardi, S. Persistent organic pollutants in some species of a Ross Sea pelagic trophic web. Antarct. Sci. 2003, 15, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Corsolini, S.; Covaci, A.; Ademollo, N.; Focardi, S.; Schepens, P. Occurrence of organochlorine pesticides (OCPs) and their enantiomeric signatures, and concentrations of polybrominated diphenyl ethers (PBDEs) in the Adélie penguin food web, Antarctica. Environ. Pollut. 2006, 140, 371–382. [Google Scholar] [CrossRef]
- Weber, K.; Goerke, H. Persistent organic pollutants (POPs) in Antarctic fish: Levels, patterns, changes. Chemosphere 2003, 53, 667–678. [Google Scholar] [CrossRef]
- Giannarelli, S.; Onor, M.; Abete, C.; Termine, M.; Fuoco, R. Effect of altitude and distance from the sea on fractionation processes of Persistent Organic Pollutants (POPs) associated to atmospheric aerosol from Ross Sea to Dome C, Antarctica. Microchem. J. 2019, 149, 103911. [Google Scholar] [CrossRef]
- Potapowicz, J.; Szumińska, D.; Szopińska, M.; Polkowska, Z. The influence of global climate change on the environmental fate of anthropogenic pollution released from the permafrost: Part, I. Case study of Antarctica. Sci. Total Environ. 2019, 651, 1534–1548. [Google Scholar] [CrossRef] [PubMed]
- Corsolini, S.; Baroni, D.; Martellini, T.; Pala, N.; Cincinelli, A. PBDEs and PCBs in terrestrial ecosystems of the Victoria Land, Antarctica. Chemosphere 2019, 231, 233–239. [Google Scholar] [CrossRef]
- Gillan, D.C.; Danis, B.; Pernet, P.; Joly, G.; Dubois, P. Structure of sediment-associated microbial communities along a heavy-metal contamination gradient in the marine environment. Appl. Environ. Microbiol. 2005, 71, 679–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.; Lee, S.-H.; Kim, M.; Kim, J.-H.; Lim, H.S. Polychlorinated biphenyl congeners in soils and lichens from King George Island, South Shetland Islands, Antarctica. Antarct. Sci. 2010, 22, 31–38. [Google Scholar] [CrossRef]
- Wang, Q.; Shi, Y.; Hu, J.; Yao, Z.; Fang, X.; Dong, Y. Determination of dioxin-like polychlorinated biphenyls in soil and moss from Fildes Peninsula, Antarctica. Chin. Sci. Bull. 2012, 57, 992–996. [Google Scholar] [CrossRef] [Green Version]
- Cabrerizo, A.; Dachs, J.; Barceló, D.; Jones, K.C. Influence of organic matter content and human activities on the occurrence of organic pollutants in antarctic soils, lichens, grass, and mosses. Environ. Sci. Technol. 2012, 46, 1396–1405. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, Q.-H.; Wang, T.; Chen, W.; Ren, D.; Li, Y.-M.; Jiang, G.-B. PCBs and PBDEs in environmental samples from King George Island and Ardley Island, Antarctica. RSC Adv. 2012, 2, 1350–1355. [Google Scholar] [CrossRef]
- Lo Giudice, A.; Casella, P.; Bruni, V.; Michaud, L. Response of bacterial isolates from Antarctic shallow sediments towards heavy metals, antibiotics and polychlorinated biphenyls. Ecotoxicology 2013, 22, 240–250. [Google Scholar] [CrossRef]
- Margesin, R. Alpine microorganisms: Useful tools for low-temperature bioremediation. J. Microbiol. 2007, 45, 281–285. [Google Scholar] [PubMed]
- Margesin, R.; Gander, S.; Zacke, G.; Gounot, A.M.; Schinner, F. Hydrocarbon degradation and enzyme activities of cold-adapted bacteria and yeasts. Extremophiles 2003, 7, 451–458. [Google Scholar] [CrossRef]
- Stibal, M.; Bælum, J.; Holben, W.E.; Sörensen, S.R.; Jensen, A.; Jacobsen, C.S. Microbial degradation of 2, 4-dichlorophenoxyacetic acid on the Greenland ice sheet. Appl. Environ. Microbiol. 2012, 78, 5070–5076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuddus, M.; Joseph, B.; Ramteke, P.W. Production of laccase from newly isolated Pseudomonas putida and its application in bioremediation of synthetic dyes and industrial effluents. Biocatal. Agric. Biotechnol. 2013, 2, 333–338. [Google Scholar] [CrossRef]
- Michaud, L.; Di Marco, G.; Bruni, V.; Lo Giudice, A. Biodegradative potential and characterization of psychrotolerant polychlorinated biphenyl-degrading marine bacteria isolated from a coastal station in the Terra Nova Bay (Ross Sea, Antarctica). Mar. Pollut. Bull. 2007, 54, 1754–1761. [Google Scholar] [CrossRef]
- Poland, J.S.; Riddle, M.J.; Zeeb, B.A. Contaminants in the Arctic and the Antarctic: A comparison of sources, impacts, and remediation options. Polar Rec. 2003, 39, 369–383. [Google Scholar] [CrossRef]
- Grannas, A.M.; Bogdal, C.; Hageman, K.J.; Halsall, C.; Harner, T.; Hung, H.; Kallenborn, R.; Klán, P.; Klánová, J.; Macdonald, R.W.; et al. The role of the global cryosphere in the fate of organic contaminants. Atmos. Chem. Phys. 2013, 13, 3271–3305. [Google Scholar] [CrossRef] [Green Version]
- Casal, P.; Cabrerizo, A.; Vila-Costa, M.; Pizarro, M.; Jiménez, B.; Dachs, J. Pivotal role of snow deposition and melting driving fluxes of polycyclic aromatic hydrocarbons at coastal Livingston Island (Antarctica). Environ. Sci. Technol. 2018, 52, 12327–12337. [Google Scholar] [CrossRef]
- Borghini, F.; Colacevich, A.; Caruso, T.; Bargagli, R. Temporal variation in the water chemistry of northern Victoria Land lakes (Antarctica). Aquat. Sci. 2008, 70, 134–141. [Google Scholar] [CrossRef]
- Malandrino, M.; Abollino, O.; Buoso, S.; Casalino, C.E.; Gasparon, M.; Giacomino, A.; La Gioia, C.; Mentasti, E. Geochemical characterisation of Antarctic soils and lacustrine sediments from Terra Nova Bay. Microchem. J. 2009, 92, 21–31. [Google Scholar] [CrossRef]
- Tanabe, S.; Hideo, H.; Tatsukawa, R. PCBs and chlorinated hydrocarbon pesticides in Antarctic atmosphere and hydrosphere. Chemosphere 1983, 12, 277–288. [Google Scholar] [CrossRef]
- Klánová, J.; Matykiewiczová, N.; Mácka, Z.; Prosek, P.; Láska, K.; Klán, P. Persistent organic pollutants in soils and sediments from James Ross Island, Antarctica. Environ. Pollut. 2008, 152, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Papale, M.; Conte, A.; Mikkonen, A.; Michaud, L.; La Ferla, R.; Azzaro, M.; Caruso, G.; Paranhos, R.; Anderson, S.C.; Maimone, G.; et al. Prokaryotic assemblages within permafrost active layer at Edmonson Point (Northern Victoria Land, Antarctica). Soil Biol. Biochem. 2018, 123, 165–179. [Google Scholar] [CrossRef]
- Bargagli, R.; Monaci, F.; Bucci, C. Environmental biogeochemistry of mercury in Antarctic ecosystems. Soil Biol. Biochem. 2007, 39, 352–360. [Google Scholar] [CrossRef]
- Vecchiato, M.; Argiriadis, E.; Zambon, S.; Barbante, C.; Toscano, G.; Gambaro, A.; Piazza, R. Persistent Organic Pollutants (POPs) in Antarctica: Occurrence in continental and coastal surface snow. Microchem. J. 2015, 119, 75–82. [Google Scholar] [CrossRef]
- Corsolini, S.; Pala, N.; Martellini, T.; Baroni, D.; Cincinelli, A. PCBs and PBDEs in soil, sediment and moss community from ponds across Victoria Land, Antarctica. Organohalogen Compd. 2018, 80, 501–504. [Google Scholar] [CrossRef]
- Cannone, N.; Guglielmin, M. Influence of vegetation on the ground thermal regime in continental Antarctica. Geoderma 2009, 151, 215–223. [Google Scholar] [CrossRef]
- Porcino, N.; Cosenza, A.; Azzaro, M. A review on the geochemistry of lakes in Victoria Land (Antarctica). Chemosphere 2020, 251, 126229. [Google Scholar] [CrossRef]
- Papale, M.; Giannarelli, S.; Francesconi, F.; Di Marco, G.; Mikkonen, A.; Conte, A.; Rizzo, C.; De Domenico, E.; Michaud, L.; Lo Giudice, A. Enrichment, isolation and biodegradation potential of psychrotolerant polychlorinated-biphenyl degrading bacteria from the Kongsfjorden (Svalbard Islands, High Arctic Norway). Mar. Pollut. Bull. 2017, 114, 849–859. [Google Scholar] [CrossRef]
- Frame, G.M.; Cochran, J.W.; Bøwadt, S.S. Complete PCB congener distributions for 17 Aroclor mixtures determined by 3 HRGC systems optimized for comprehensive, quantitative, congener specific analysis. J. High Resolut. Chromatogr. 1996, 19, 657–668. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTALW: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Saitou, M.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tamura, K.; Nei, M. MEGA3: Integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief Bioinform. 2004, 2, 150–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehtinen, T.; Mikkonen, A.; Sigfusson, B.; Ólafsdóttir, K.; Ragnarsdóttir, K.V.; Guicharnaud, R. Bioremediation trial on aged PCB-polluted soils—a bench study in Iceland. Environ. Sci. Pollut. Res. 2013, 21, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Vézina, J.; Barriault, D.; Sylvestre, M. Diversity of the C-terminal portion of the biphenyl dioxygenase large subunit. J. Mol. Microbiol. Biotechnol. 2008, 15, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Witzig, R.; Junca, H.; Hecht, H.-J.; Pieper, D.H. Assessment of toluene/ biphenyl dioxygenase gene diversity in benzene-polluted soils: Link between benzene biodegradation and genes similar to those encoding isopropylbenzene dioxygenases. Appl. Environ. Microbiol. 2006, 72, 3504–3514. [Google Scholar] [CrossRef] [Green Version]
- Hauptmann, A.L.; Sicheritz-Pontén, T.; Cameron, K.A.; Bælum, J.; Plichta, D.R.; Dalgaard, M.; Stibal, M. Contamination of the Arctic reflected in microbial metagenomes from the Greenland ice sheet. Environ. Res. Lett. 2017, 12, 074019. [Google Scholar] [CrossRef]
- Mohn, W.W.; Westerberg, K.; Cullen, W.R.; Reimer, K.J. Aerobic biodegradation of biphenyl and polychlorinated biphenyls by Arctic soil micro-organisms. Appl. Environ. Microbiol. 1997, 63, 3378–3384. [Google Scholar] [CrossRef] [Green Version]
- Master, E.R.; Mohn, W.W. Psychrotolerant bacteria isolated from Arctic soil that degrade polychlorinated biphenyls at low temperatures. Appl. Environ. Microbiol. 1998, 64, 4823–4829. [Google Scholar] [CrossRef] [Green Version]
- Ahn, Y.-B.; Beaudette, L.A.; Lee, H.; Trevors, J.T. Survival of a GFP-labeled polychlorobiphenyl degrading psychrotolerant Pseudomonas spp. in 4 and 22 °C soil microcosms. Microb. Ecol. 2001, 42, 614–623. [Google Scholar] [CrossRef]
- Kuipers, B.; Cullen, W.R.; Mohn, W.W. Reductive dechlorination of weathered Aroclor 1260 during anaerobic biotreatment of Arctic soils. Can. J. Microbiol. 2003, 49, 9–14. [Google Scholar] [CrossRef]
- Margesin, R.; Zacke, G.; Schinner, F. Characterization of heterotrophic microorganisms in alpine glacier cryoconite. Arctic 2002, 34, 88–93. [Google Scholar] [CrossRef]
- Rappazzo, A.C.; Papale, M.; Rizzo, C.; Conte, A.; Giannarelli, S.; Onor, M.; Abete, C.; Cefali, P.; De Domenico, E.; Michaud, L.; et al. Heavy metal tolerance and polychlorinated biphenyl oxidation in bacterial communities inhabiting the Pasvik River and the Varanger Fjord area (Arctic Norway). Mar. Pollut. Bull. 2019, 141, 535–549. [Google Scholar] [CrossRef]
- Yakimov, M.M.; Giuliano, L.; Bruni, V.; Scarfì, S.; Golyshin, P.N. Characterization of Antarctic hydrocarbon-degrading bacteria capable of producing bioemulsifiers. New Microbiol. 1999, 22, 249–256. [Google Scholar]
- De Domenico, M.; Giudice, A.L.; Michaud, L.; Saitta, M.; Bruni, V. Diesel oil and PCB-degrading bacteria isolated from Antarctic seawaters (Terra Nova Bay, Ross Sea). Polar Res. 2004, 23, 141–146. [Google Scholar] [CrossRef]
- Tumeo, M.A.; McGuinn, D.A. Evaluation of bioremediation in cold regions. J. Cold Reg. Engin. 1997, 11, 221–231. [Google Scholar] [CrossRef]
- Luz, A.P.; Pellizari, V.H.; Whyte, L.G.; Greer, C.W. A survey of indigenous microbial hydrocarbon degradation genes in soils from Antarctica and Brazil. Can. J. Microbiol. 2004, 50, 323–333. [Google Scholar] [CrossRef]
- Rizzo, C.; Conte, A.; Azzaro, M.; Papale, M.; Rappazzo, A.C.; Battistel, D.; Roman, M.; Lo Giudice, A.; Guglielmin, M. Cultivable bacterial communities in brines from perennially ice-covered and pristine antarctic lakes: Ecological and biotechnological implications. Microorganisms 2020, 8, 819. [Google Scholar] [CrossRef] [PubMed]
- Wania, F. Assessing the potential of Persistent Organic Chemicals for long-range transport and accumulation in Polar Regions. Environ. Sci. Technol. 2003, 37, 1344–1351. [Google Scholar] [CrossRef]
- Jordahl, J.L.; Foster, L.; Schnoor, J.L.; Alvarez, P.J.J. Effect of hybrid poplar trees on microbial populations important to hazardous waste bioremediation. Environ. Toxicol. Chem. 1997, 16, 1318–1321. [Google Scholar] [CrossRef]
- Ding, N.; Guo, H.; Hayat, T.; Wu, Y.; Xu, J. Microbial community structure changes during Aroclor 1242 degradation in the rhizosphere of ryegrass (Lolium multiflorum L.). FEMS Microbiol. Ecol. 2009, 70, 305–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.K.; Lee, J.; Sul, W.J.; Iwai, S.; Chai, B.; Tiedje, J.M.; Park, J. Novel biphenyl-oxidizing bacteria and dioxygenase genes from a Korean tidal mudflat. Appl. Environ. Microbiol. 2011, 77, 3888–3891. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, K. Oxygenases and dehalogenases: Molecular approaches to efficient degradation of chlorinated environmental pollutants. Biosci. Biotechnol. Biochem. 2006, 70, 2335–2348. [Google Scholar] [CrossRef] [PubMed]
- Hayase, N.; Taira, K.; Furukawa, K. Pseudomonas putida KF715 bphABCD operon encoding biphenyl and polychlorinated biphenyl degradation: Cloning, analysis, and expression in soil bacteria. J. Bacteriol. 1990, 172, 1160–1164. [Google Scholar] [CrossRef] [Green Version]
- Master, E.R.; Mohn, W.W. Induction of bphA, encoding biphenyl dioxygenase, in two polychlorinated biphenyl-degrading bacteria, psychrotolerant Pseudomonas strain Cam-1 and Mesophilic Burkholderia strain LB400. Appl. Environ. Microbiol. 2001, 67, 2669–2676. [Google Scholar] [CrossRef] [Green Version]
- Mouz, S.; Merlin, C.; Springael, D.; Toussaint, A. A GntR-like negative regulator of the biphenyl degradation genes of the transposon Tn4371. Mol. Gen. Genet. 1999, 262, 790–799. [Google Scholar] [CrossRef]
- Watanabe, K.; Futamata, H.; Harayama, S. Understanding the diversity in catabolic potential of microorganisms for the development of biodegradation strategies. Antony Van Leeuwen. 2002, 81, 655–663. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Lake ID | Reported Name | Coordinates | Altitude (m) | Temperature (°C) | Dissolved O2 (ppm) | pH | Conductivity (µm s−1) | Salinity |
|---|---|---|---|---|---|---|---|---|
| EP1 | Edmonson Point 13 | 74°20′ S–165°08′ E | 0 | 4.7 | 9.3 | 6.5 | 1442.0 | 5 |
| EP2 | Edmonson Point 14 | 74°20′ S–165°08′ E | 20 | 1.1 | 10.6 | 9.1 | 5.1 | 8 |
| EP3 | Edmonson Point 15 | 74°20′ S–165°04′ E | 3 | 3.6 | 9.5 | 8.1 | 51.3 | 5 |
| EP4 | Edmonson Point 16 | 74°22′ S–165°06′ E | 10 | 0.5 | 8.5 | 7.4 | 223.0 | 5 |
| Primer Set (n.) | Primer(s) | Sequence (5′ to 3′) | Annealing Temperature | Reference |
|---|---|---|---|---|
| 1 | BPHA1REV11150 | AKW-YYC-SCC-GTC-GTC-CTG-MTC | 60 °C | [45] |
| 2BPHFWD1 | GGC-TGG-GCC-TAC-GAC-ANC-GC | |||
| 2 | 2BPHAREV1 | ADV-CCS-GCB-GCC-GCB-TCH-TCG | 58 °C | [46] |
| 2BPHFWD1 | GGC-TGG-GCC-TAC-GAC-ANC-GC | |||
| 3 | BPHA1REV11150 | AKW-YYC-SCC-GTC-GTC-CTG-MTC | 60 °C | [47] |
| BPHA1FWD1654 | AAY-TGG-GCC-TAC-GAC-ANC-GC |
| PCB-Oxidizing Isolates (n.) | ||
|---|---|---|
| Lake ID | Water | Sediment |
| EP1 | 6 | 2 |
| EP2 | 11 | 6 |
| EP3 | 7 | 13 |
| EP4 | - | 12 |
| Phylum or Class * | Strain | AN | MNA Code | Next Relative by GenBank Alignment (AN **, Organism) | Isolation Matrix | Hom § (%) | bphA Gene |
|---|---|---|---|---|---|---|---|
| BET | EP1-3 | OL470332 | MNA-CIBAN-1523 | NR116103.1, Pusillimonas ginsengisoli strain DCY25 | Water | 98 | + |
| EP1-35 | OL470333 | MNA-CIBAN-1524 | NR116103.1, Pusillimonas ginsengisoli strain DCY25 | Sediment | 98 | + | |
| EP2-12 | OL470334 | MNA-CIBAN-1525 | NR116103.1, Pusillimonas ginsengisoli strain DCY25 | Water | 98 | + | |
| EP3-42 | OL470335 | MNA-CIBAN-1526 | NR116103.1, Pusillimonas ginsengisoli strain DCY25 | Sediment | 96 | + | |
| EP3-46 | OL470336 | MNA-CIBAN-1527 | NR116103.1, Pusillimonas ginsengisoli strain DCY25 | Sediment | 94 | - | |
| EP2-20 | OL470337 | MNA-CIBAN-1528 | NR116103.1, Pusillimonas ginsengisoli strain DCY25 | Water | 97 | + | |
| EP2-15 | OL470338 | MNA-CIBAN-1529 | NR116103.1, Pusillimonas ginsengisoli strain DCY25 | Water | 98 | + | |
| EP3-34 | OL470339 | MNA-CIBAN-1530 | NR116103.1, Pusillimonas ginsengisoli strain DCY25 | Sediment | 98 | - | |
| EP1-31 | OL470340 | MNA-CIBAN-1531 | NR116103.1, Pusillimonas ginsengisoli strain DCY25 | Sediment | 98 | - | |
| GAM | EP4-4 | OL470341 | MNA-CIBAN-1532 | KY476459, Lysobacter sp. strain A5-17 | Sediment | 99 | + |
| FIR | EP4-7 | OL470342 | MNA-CIBAN-1533 | CP013661, Planococcus kocurii ATCC 43650 | Sediment | 100 | - |
| ACT | EP3-13 | OL470343 | MNA-CIBAN-1534 | JX428856, Salinibacterium sp. ZS217 | Water | 99 | - |
| EP3-15 | OL470344 | MNA-CIBAN-1535 | JX428856, Salinibacterium sp. ZS217 | Water | 100 | - | |
| EP3-3 | OL470345 | MNA-CIBAN-1536 | JX428856, Salinibacterium sp. ZS217 | Water | 98 | - | |
| EP1-8 | OL470346 | MNA-CIBAN-1537 | JX428856, Salinibacterium sp. ZS217 | Water | 99 | - | |
| EP1-15 | OL470347 | MNA-CIBAN-1538 | JX428856, Salinibacterium sp. ZS217 | Water | 99 | - | |
| EP1-16 | OL470348 | MNA-CIBAN-1539 | JX428856, Salinibacterium sp. ZS217 | Water | 100 | + | |
| EP1-17 | OL470349 | MNA-CIBAN-1540 | JX428856, Salinibacterium sp. ZS217 | Water | 100 | + | |
| EP3-16 | OL470350 | MNA-CIBAN-1541 | JX428856, Salinibacterium sp. ZS217 | Water | 99 | + | |
| EP3-18 | OL470351 | MNA-CIBAN-1542 | JX428856, Salinibacterium sp. ZS217 | Water | 99 | + | |
| EP3-21 | OL470352 | MNA-CIBAN-1543 | JX428856, Salinibacterium sp. ZS217 | Water | 99 | + | |
| EP2-1 | OL470353 | MNA-CIBAN-1549 | KM891542, S. amurskyense AP9-24B | Water | 99 | - | |
| EP2-43 | OL470354 | MNA-CIBAN-1544 | MT890162, Leifsonia rubra strain G.S.5 | Sediment | 96 | + | |
| EP3-28 | OL470355 | MNA-CIBAN-1546 | MT890162, Leifsonia rubra strain G.S.5 | Sediment | 99 | + | |
| EP3-17 | OL470356 | MNA-CIBAN-1545 | MH482237, Leifsonia rubra strain 4ABZ17 | Water | 99 | + | |
| EP2-2 | OL470357 | MNA-CIBAN-1547 | MK660300, Leifsonia sp. strain SER12 | Water | 99 | - | |
| EP2-8 | OL470358 | MNA-CIBAN-1548 | KY476591, Leifsonia sp. strain F3-8 | Water | 99 | + | |
| EP2-32 | OL470359 | MNA-CIBAN-1550 | KR023886, Arthrobacter antarcticus strain TBQH1 | Sediment | 99 | + | |
| EP2-46 | OL470360 | MNA-CIBAN-1551 | KR023886, Arthrobacter antarcticus strain TBQH1 | Sediment | 99 | + | |
| EP2-31 | OL470361 | MNA-CIBAN-1552 | KR023886, Arthrobacter antarcticus strain TBQH1 | Sediment | 99 | + | |
| EP3-44 | OL470362 | MNA-CIBAN-1553 | MW295493, Arthrobacter sp. strain Y36 | Sediment | 99 | + | |
| EP3-29 | OL470363 | MNA-CIBAN-1556 | MW295493, Arthrobacter sp. strain Y36 | Sediment | 99 | + | |
| EP1-13 | OL470364 | MNA-CIBAN-1554 | MH018889, Arthrobacter sp. strain Kopri-28 | Water | 99 | + | |
| EP4-11 | OL470365 | MNA-CIBAN-1555 | AB920568, Arthrobacter alpinus strain S6-3 | Sediment | 99 | - | |
| EP2-42 | OL470366 | MNA-CIBAN-1557 | MW960234, A. livingstonensis NJ-QEDSD-9-C | Sediment | 100 | + | |
| EP3-27 | OL470367 | MNA-CIBAN-1558 | MT622202, Arthrobacter sp. strain R16 | Sediment | 99 | + | |
| EP3-43 | OL470368 | MNA-CIBAN-1559 | MT622202, Arthrobacter sp. strain R16 | Sediment | 100 | + | |
| EP3-26, EP3-30, EP3-41 | Unidentified | Sediment | -- | + | |||
| EP2-10, EP1-11, EP2-11, EP2-14 | Unidentified | Water | -- | + | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papale, M.; Lo Giudice, A.; Rappazzo, A.C.; Azzaro, M.; Rizzo, C. A First Glimpse on Cold-Adapted PCB-Oxidizing Bacteria in Edmonson Point Lakes (Northern Victoria Land, Antarctica). Water 2022, 14, 109. https://doi.org/10.3390/w14010109
Papale M, Lo Giudice A, Rappazzo AC, Azzaro M, Rizzo C. A First Glimpse on Cold-Adapted PCB-Oxidizing Bacteria in Edmonson Point Lakes (Northern Victoria Land, Antarctica). Water. 2022; 14(1):109. https://doi.org/10.3390/w14010109
Chicago/Turabian StylePapale, Maria, Angelina Lo Giudice, Alessandro Ciro Rappazzo, Maurizio Azzaro, and Carmen Rizzo. 2022. "A First Glimpse on Cold-Adapted PCB-Oxidizing Bacteria in Edmonson Point Lakes (Northern Victoria Land, Antarctica)" Water 14, no. 1: 109. https://doi.org/10.3390/w14010109
APA StylePapale, M., Lo Giudice, A., Rappazzo, A. C., Azzaro, M., & Rizzo, C. (2022). A First Glimpse on Cold-Adapted PCB-Oxidizing Bacteria in Edmonson Point Lakes (Northern Victoria Land, Antarctica). Water, 14(1), 109. https://doi.org/10.3390/w14010109