
Aquatic Vegetation Loss and Its Implication on Climate Regulation in a Protected Freshwater Wetland of Po River Delta Park (Italy)

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
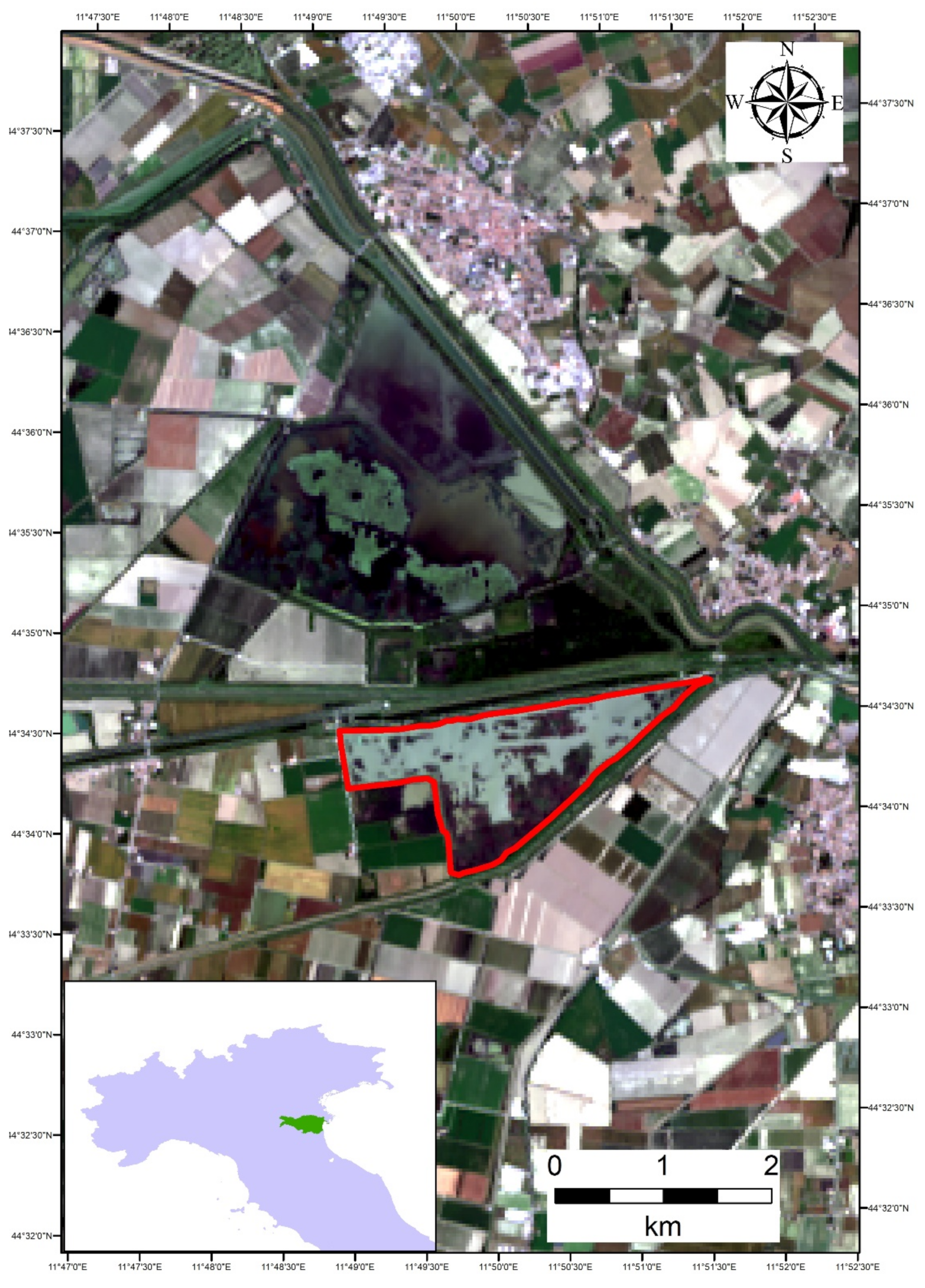
2.1. Study Area
2.2. Loss of Aquatic Vegetation and Aboveground Biomass
2.3. Breakdown Rate and Climate Regulation
3. Results
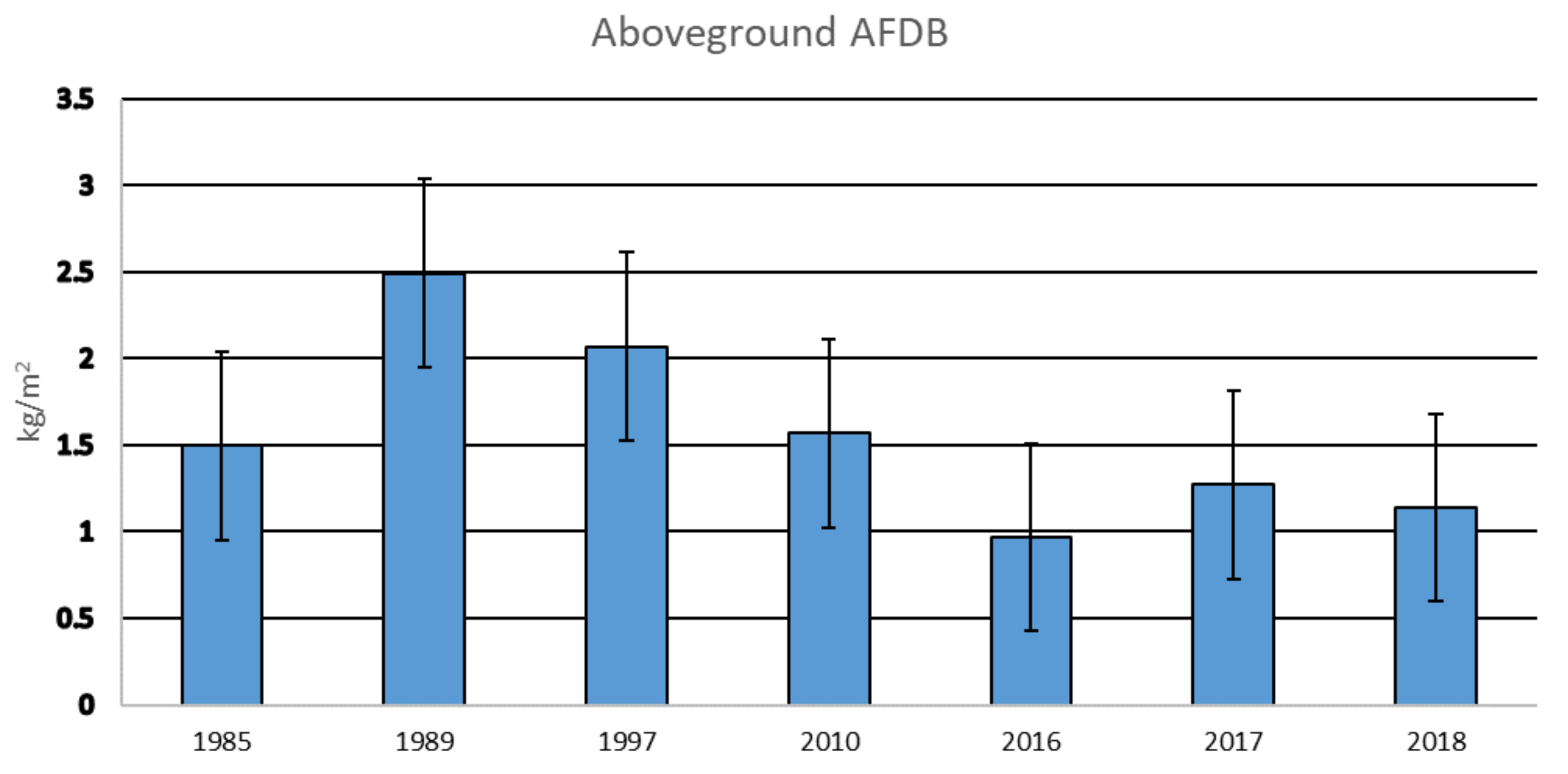
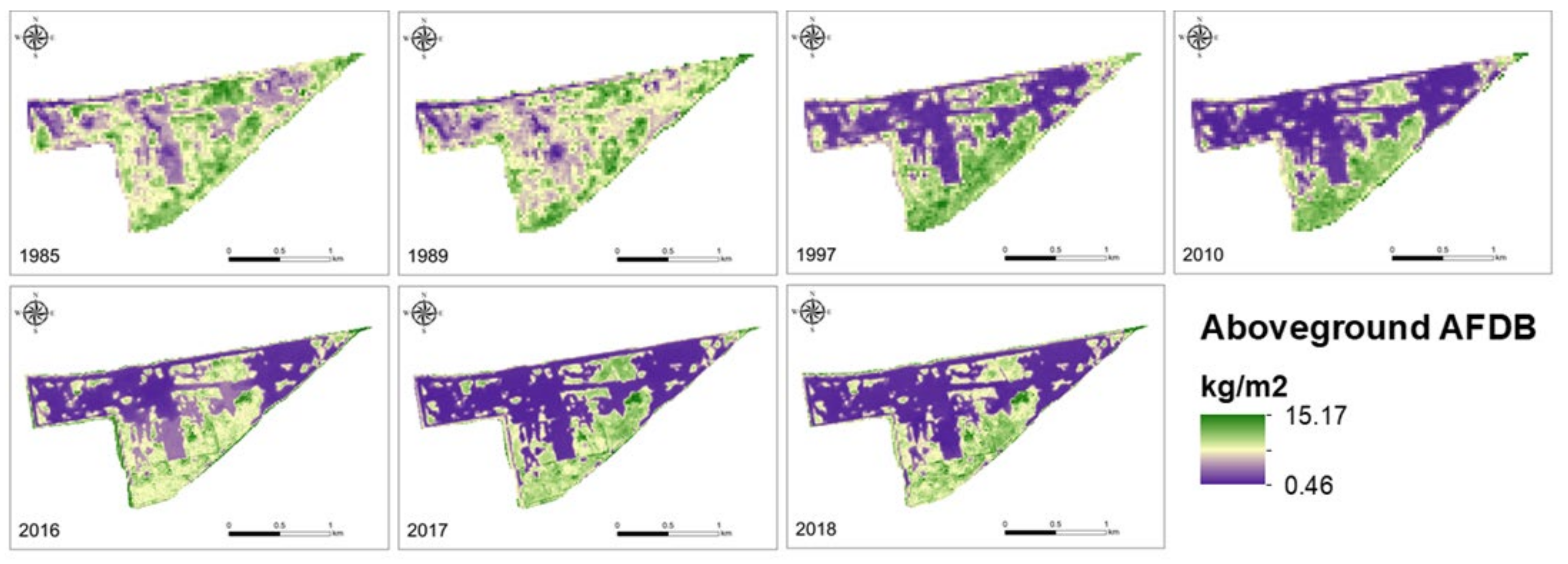
3.1. Aboveground Biomass over Time
3.2. Biomass Decomposition
3.3. Carbon Storage and Sequestration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitsch, W.J.; Bernal, B.; Hernandez, M.E. Ecosystem services of wetlands. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2015, 11, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chen, M.; Yang, G.; Jiang, B.; Zhang, J. Wetland ecosystem services research: A critical review. Glob. Ecol. Conserv. 2020, 22, e01027. [Google Scholar] [CrossRef]
- Kingsford, R.T.; Basset, A.; Jackson, L. Wetlands: Conservation’s poor cousins. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 892–916. [Google Scholar] [CrossRef] [Green Version]
- Xi, Y.; Peng, S.; Ciais, P.; Chen, Y. Future impacts of climate change on inland Ramsar wetlands. Nat. Clim. Chang. 2021, 11, 45–51. [Google Scholar] [CrossRef]
- Millennium Ecosystem Assessment. Ecosystems and Human Well-being. In Synthesis; Island Press: Washington, DC, USA, 2005; Volume 5, ISBN 1597260401.
- Hettiarachchi, M.; Morrison, T.H.; McAlpine, C. Forty-three years of ramsar and urban wetlands. Glob. Environ. Chang. 2015, 32, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Weng, B.; Yan, D.; Wang, K.; Li, X.; Bi, W.; Li, M.; Cheng, X.; Liu, Y. Wetlands of international importance: Status, threats, and future protection. Int. J. Environ. Res. Public Health 2019, 16, 1818. [Google Scholar] [CrossRef] [Green Version]
- Gaglio, M.; Aschonitis, V.G.; Gissi, E.; Castaldelli, G.; Fano, E.A. Land use change effects on ecosystem services of river deltas and coastal wetlands: Case study in Volano–Mesola–Goro in Po river delta (Italy). Wetl. Ecol. Manag. 2017, 25, 67–86. [Google Scholar] [CrossRef]
- Fogli, S.; Marchesini, R.; Gerdol, R. Reed (Phragmites australis) decline in a brackish wetland in Italy. Mar. Environ. Res. 2002, 53, 465–479. [Google Scholar] [CrossRef]
- Lastrucci, L.; Lazzaro, L.; Coppi, A.; Foggi, B.; Ferranti, F.; Venanzoni, R.; Cerri, M.; Ferri, V.; Gigante, D.; Reale, L. Demographic and macro-morphological evidence for common reed dieback in central Italy. Plant Ecol. Divers. 2017, 10, 241–251. [Google Scholar] [CrossRef]
- Zhang, Y.; Jeppesen, E.; Liu, X.; Qin, B.; Shi, K.; Zhou, Y.; Thomaz, S.M.; Deng, J. Global loss of aquatic vegetation in lakes. Earth-Sci. Rev. 2017, 173, 259–265. [Google Scholar] [CrossRef]
- Martínez, A.; Larrañaga, A.; Pérez, J.; Basaguren, A.; Pozo, J. Leaf-litter quality effects on stream ecosystem functioning: A comparison among five species. Fundam. Appl. Limnol. 2013, 183, 239–248. [Google Scholar] [CrossRef]
- Sauer, F.G.; Bundschuh, M.; Zubrod, J.P.; Schäfer, R.B.; Thompson, K.; Kefford, B.J. Effects of salinity on leaf breakdown: Dryland salinity versus salinity from a coalmine. Aquat. Toxicol. 2016, 177, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Paul, M.J.; Meyer, J.L.; Couch, C.A. Leaf breakdown in streams differing in catchment land use. Freshw. Biol. 2006, 51, 1684–1695. [Google Scholar] [CrossRef]
- Arroita, M.; Aristi, I.; Díez, J.; Martinez, M.; Oyarzun, G.; Elosegi, A. Impact of water abstraction on storage and breakdown of coarse organic matter in mountain streams. Sci. Total Environ. 2015, 503, 233–240. [Google Scholar] [CrossRef]
- De Mello Cionek, V.; Fogaça, F.N.O.; Moulton, T.P.; Pazianoto, L.H.R.; Landgraf, G.O.; Benedito, E. Influence of leaf miners and environmental quality on litter breakdown in tropical headwater streams. Hydrobiologia 2021, 848, 1311–1331. [Google Scholar] [CrossRef]
- Cunha, E.R.; Winemiller, K.O.; da Silva, J.C.B.; Lopes, T.M.; Gomes, L.C.; Thomaz, S.M.; Agostinho, A.A. α and β diversity of fishes in relation to a gradient of habitat structural complexity supports the role of environmental filtering in community assembly. Aquat. Sci. 2019, 81, 1–12. [Google Scholar] [CrossRef]
- Gaglio, M.; Lanzoni, M.; Nobili, G.; Viviani, D.; Castaldelli, G.; Fano, E.A. Ecosystem Services approach for sustainable governance in a brackish water lagoon used for aquaculture. J. Environ. Plan. Manag. 2019, 62, 1501–1524. [Google Scholar] [CrossRef]
- Caraco, N.; Cole, J.; Findlay, S.; Wigand, C. Vascular plants as engineers of oxygen in aquatic systems. Bioscience 2006, 56, 219–225. [Google Scholar] [CrossRef]
- Soana, E.; Gavioli, A.; Tamburini, E.; Fano, E.A.; Castaldelli, G. To mow or not to mow: Reed biofilms as denitrification hotspots in drainage canals. Ecol. Eng. 2018, 113, 1–10. [Google Scholar] [CrossRef]
- Klemas, V. Remote sensing of emergent and submerged wetlands: An overview. Int. J. Remote Sens. 2013, 34, 6286–6320. [Google Scholar] [CrossRef]
- Guo, M.; Li, J.; Sheng, C.; Xu, J.; Wu, L. A review of wetland remote sensing. Sensors 2017, 17, 777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdavi, S.; Salehi, B.; Granger, J.; Amani, M.; Brisco, B.; Huang, W. Remote sensing for wetland classification: A comprehensive review. GIScience Remote Sens. 2018, 55, 623–658. [Google Scholar] [CrossRef]
- Mishra, N.B. Wetlands: Remote Sensing. In Wetlands and Habitats; Wang, Y., Ed.; CRC Press: Boca Raton, CA, USA, 2020; pp. 201–212. ISBN 9780429445507. [Google Scholar]
- Liira, J.; Feldmann, T.; Mäemets, H.; Peterson, U. Two decades of macrophyte expansion on the shores of a large shallow northern temperate lake-A retrospective series of satellite images. Aquat. Bot. 2010, 93, 207–215. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Shi, K.; Zhou, Y.; Tang, X.; Zhu, G.; Qin, B. Mapping aquatic vegetation in a large, shallow eutrophic lake: A frequency-based approach using multiple years of MODIS data. Remote Sens. 2015, 7, 10295–10320. [Google Scholar] [CrossRef] [Green Version]
- Villa, P.; Bresciani, M.; Bolpagni, R.; Pinardi, M.; Giardino, C. A rule-based approach for mapping macrophyte communities using multi-temporal aquatic vegetation indices. Remote Sens. Environ. 2015, 171, 218–233. [Google Scholar] [CrossRef] [Green Version]
- Berhane, T.M.; Lane, C.R.; Wu, Q.; Autrey, B.C.; Anenkhonov, O.A.; Chepinoga, V.V.; Liu, H. Decision-tree, rule-based, and random forest classification of high-resolution multispectral imagery for wetland mapping and inventory. Remote Sens. 2018, 10, 580. [Google Scholar] [CrossRef] [Green Version]
- Wilson, N.R.; Norman, L.M. Analysis of vegetation recovery surrounding a restored wetland using the normalized difference infrared index (NDII) and normalized difference vegetation index (NDVI). Int. J. Remote Sens. 2018, 39, 3243–3274. [Google Scholar] [CrossRef] [Green Version]
- Wulder, M.A.; White, J.C.; Loveland, T.R.; Woodcock, C.E.; Belward, A.S.; Cohen, W.B.; Fosnight, E.A.; Shaw, J.; Masek, J.G.; Roy, D.P. The global Landsat archive: Status, consolidation, and direction. Remote Sens. Environ. 2016, 185, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Woodcock, C.E.; Allen, R.; Anderson, M.; Belward, A.; Bindschadler, R.; Cohen, W.; Gao, F.; Goward, S.N.; Helder, D.; Helmer, E.; et al. Free access to landsat imagery. Science 2008, 320, 1011. [Google Scholar] [CrossRef]
- Fickas, K.C.; Cohen, W.B.; Yang, Z. Landsat-based monitoring of annual wetland change in the Willamette Valley of Oregon, USA from 1972 to 2012. Wetl. Ecol. Manag. 2016, 24, 73–92. [Google Scholar] [CrossRef]
- Mahdianpari, M.; Jafarzadeh, H.; Granger, J.E.; Mohammadimanesh, F.; Brisco, B.; Salehi, B.; Homayouni, S.; Weng, Q. A large-scale change monitoring of wetlands using time series Landsat imagery on Google Earth Engine: A case study in Newfoundland. GIScience Remote Sens. 2020, 57, 1102–1124. [Google Scholar] [CrossRef]
- Pinardi, M.; Bresciani, M.; Villa, P.; Cazzaniga, I.; Laini, A.; Tóth, V.; Fadel, A.; Austoni, M.; Lami, A.; Giardino, C. Spatial and temporal dynamics of primary producers in shallow lakes as seen from space: Intra-annual observations from Sentinel-2A. Limnologica 2018, 72, 32–43. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Gill, L.; Regan, S.; Naughton, O.; Johnston, P.; Waldren, S.; Ghosh, B. Mapping Vegetation Communities Inside Wetlands Using Sentinel-2 Imagery in Ireland. Int. J. Appl. Earth Obs. Geoinf. 2020, 88, 102083. [Google Scholar] [CrossRef]
- Piccoli, F. Flora e vegetazione delle casse di espansione di Campotto e Valle Santa (Ferrara). Biogeogr. J. Integr. Biogeogr. 1979, 6, 231–262. [Google Scholar] [CrossRef]
- Bower, C.E.; Holm-Hansen, T. A Salicylate–Hypochlorite Method for Determining Ammonia in Seawater. Can. J. Fish. Aquat. Sci. 1980, 37, 794–798. [Google Scholar] [CrossRef]
- Armstrong, F.A.J.; Stearns, C.R.; Strickland, J.D.H. The measurement of upwelling and subsequent biological process by means of the Technicon Autoanalyzer® and associated equipment. Deep. Res. Oceanogr. Abstr. 1967, 14, 381–389. [Google Scholar] [CrossRef]
- APHA; AWWA; WEF. Standard Methods, For the Examination of Water and Freshwater, 18th ed.; Greenberg, A.E., Clesceri, L.S., Eaton, A.D., Eds.; American Public Health Association: Washington, DC, USA, 1992. [Google Scholar]
- Vermote, E.F.; El Saleous, N.; Justice, C.O.; Kaufman, Y.J.; Privette, J.L.; Remer, L.; Roger, J.C.; Tanré, D. Atmospheric correction of visible to middle-infrared EOS-MODIS data over land surfaces: Background, operational algorithm and validation. J. Geophys. Res. Atmos. 1997, 102, 17131–17141. [Google Scholar] [CrossRef] [Green Version]
- Acker, J.G.; Leptoukh, G. Online analysis enhances use of NASA Earth Science Data. Eos 2007, 88, 14–17. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Vina, A.; Arkebauer, T.J.; Rundquist, D.C.; Keydan, G.; Leavitt, B. Remote estimation of leaf area index and green leaf biomass in maize canopies. Geophys. Res. Lett. 2003, 30, 1248. [Google Scholar] [CrossRef] [Green Version]
- Dash, J.; Curran, P.J. The MERIS terrestrial chlorophyll index. Int. J. Remote Sens. 2004, 25, 5403–5413. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Pattey, E.; Zarco-Tejada, P.J.; Strachan, I.B. Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies: Modeling and validation in the context of precision agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deeering, D. Monitoring vegetation systems in the Great Plains with ERTS (Earth Resources Technology Satellite). In Proceedings of the Third Earth Resources Technology Satellite-1 Symposium, Washington, DC, USA, 10–14 December 1973; pp. 309–317. [Google Scholar]
- Villa, P.; Laini, A.; Bresciani, M.; Bolpagni, R. A remote sensing approach to monitor the conservation status of lacustrine Phragmites australis beds. Wetl. Ecol. Manag. 2013, 21, 399–416. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Petersen, R.C.; Cummins, K.W. Leaf processing in a woodland stream. Freshw. Biol. 1974, 4, 343–368. [Google Scholar] [CrossRef]
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Duke, S.T.; Francoeur, S.N.; Judd, K.E. Effects of Phragmites australis Invasion on Carbon Dynamics in a Freshwater Marsh. Wetlands 2015, 35, 311–321. [Google Scholar] [CrossRef]
- Ricke, K.; Drouet, L.; Caldeira, K.; Tavoni, M. Country-level social cost of carbon. Nat. Clim. Chang. 2018, 8, 895–900. [Google Scholar] [CrossRef]
- Van Der Putten, W.H. Die-back of Phragmites australis in European wetlands: An overview of the European Research Programme on Reed Die-back and Progression (1993–1994). Aquat. Bot. 1997, 59, 263–275. [Google Scholar] [CrossRef]
- Cerri, M.; Ferranti, F.; Coppi, A.; Foggi, B.; Gigante, D.; Lastrucci, L.; Onofri, A.; Venanzoni, R.; Viciani, D.; Reale, L. Influence of die-back syndrome on reproductive strategies within Phragmites australis populations. Plant Biosyst. 2019, 153, 250–256. [Google Scholar] [CrossRef]
- Lastrucci, L.; Cerri, M.; Coppi, A.; Ferranti, F.; Ferri, V.; Foggi, B.; Lazzaro, L.; Reale, L.; Venanzoni, R.; Viciani, D.; et al. Understanding common reed die-back: A phytocoenotic approach to explore the decline of palustrine ecosystems. Plant Sociol. 2017, 54, 15–28. [Google Scholar] [CrossRef]
- Scarton, F.; Day, J.W.; Rismondo, A. Primary production and decomposition of Sarcocornia fruticosa (L.) Scott and Phragmites australis Trin. Ex Steudel in the Po Delta, Italy. Estuaries 2002, 25, 325–336. [Google Scholar] [CrossRef]
- Brix, H.; Sorrell, B.K.; Lorenzen, B. Are phragmites-dominated wetlands a net source or net sink of greenhouse gases? Aquat. Bot. 2001, 69, 313–324. [Google Scholar] [CrossRef]
- Longhi, D.; Bartoli, M.; Viaroli, P. Decomposition of four macrophytes in wetland sediments: Organic matter and nutrient decay and associated benthic processes. Aquat. Bot. 2008, 89, 303–310. [Google Scholar] [CrossRef]
- Gessner, M.O. Breakdown and nutrient dynamics of submerged phragmites shoots in the littoral zone of a temperate hardwater lake. Aquat. Bot. 2000, 66, 9–20. [Google Scholar] [CrossRef]
- Bertoli, M.; Brichese, G.; Michielin, D.; Ruzič, M.; Vignes, F.; Basset, A.; Pizzul, E. Seasonal and multi-annual patterns of Phragmites australis decomposition in a wetland of the Adriatic area (Northeast Italy): A three-years analysis. Knowl. Manag. Aquat. Ecosyst. 2016, 417, 14. [Google Scholar] [CrossRef] [Green Version]
- Gaberščik, A.; Grašič, M.; Abram, D.; Zelnik, I. Water level fluctuations and air temperatures affect common reed habitus and productivity in an intermittent wetland ecosystem. Water 2020, 12, 2806. [Google Scholar] [CrossRef]
- Dolinar, N.; Regvar, M.; Abram, D.; Gaberščik, A. Water-level fluctuations as a driver of Phragmites australis primary productivity, litter decomposition, and fungal root colonisation in an intermittent wetland. Hydrobiologia 2016, 774, 69–80. [Google Scholar] [CrossRef]
- Lanzoni, M.; Milardi, M.; Aschonitis, V.; Fano, E.A.; Castaldelli, G. A regional fish inventory of inland waters in Northern Italy reveals the presence of fully exotic fish communities. Eur. Zool. J. 2018, 85, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Mitsch, W.J.; Mander, Ü. Wetlands and carbon revisited. Ecol. Eng. 2018, 114, 1–6. [Google Scholar] [CrossRef]
- Grünfeld, S.; Brix, H. Methanogenesis and methane emissions: Effects of water table, substrate type and presence of Phragmites australis. Aquat. Bot. 1999, 64, 63–75. [Google Scholar] [CrossRef]
- Soana, E.; Gavioli, A.; Vincenzi, F.; Fano, E.A.; Castaldelli, G. Nitrate availability affects denitrification in Phragmites australis sediments. J. Environ. Qual. 2020, 49, 194–209. [Google Scholar] [CrossRef]
- Castaldelli, G.; Aschonitis, V.; Vincenzi, F.; Fano, E.A.; Soana, E. The effect of water velocity on nitrate removal in vegetated waterways. J. Environ. Manage 2018, 215, 230–238. [Google Scholar] [CrossRef]
- Gavioli, A.; Milardi, M.; Lanzoni, M.; Mantovani, S.; Aschonitis, V.; Soana, E.; Fano, E.A.; Castaldelli, G. Managing the environment in a pinch: Red swamp crayfish tells a cautionary tale of ecosystem based management in northeastern Italy. Ecol. Eng. 2018, 120, 546–553. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Date | Satellite |
|---|---|
| 9 October 1985 | Landsat 5-TM |
| 4 October 1989 | Landsat 5-TM |
| 24 September 1997 | Landsat 5-TM |
| 12 September 2010 | Landsat 5-TM |
| 15 November 2016 | Sentinel 2-MSI |
| 21 September 2017 | Sentinel 2-MSI |
| 31 October 2018 | Sentinel 2-MSI |
| Vegetation Index | Reference | Fittest Model | |||
|---|---|---|---|---|---|
| R2 | p-Value | Model Type | Equation | ||
| Chlorophyll Index Green (CIGreen) | [42] | 0.837 | <0.01 | Exponential | y = e(6.29595+0.660924x) |
| MERIS Terrestrial Chlorophyll Index (MTCI) | [43] | 0.803 | <0.01 | Squared | y = 595.21 + 4747.09x2 |
| Modified Chlorophyll Absorption in Reflectance Index (MCARI) | [44] | 0.792 | <0.01 | Exponential | y = e(6.32046+9.87445x) |
| Normalized Difference Vegetation Index (NDVI) | [45] | 0.731 | 0.014 | Exponential | y = e(5.82568+2.94848x) |
| Normalized Difference Aquatic Vegetation Index (NDAVI) | [46] | 0.709 | <0.01 | Exponential | y = e(5.8476+2.85724x) |
| Enhanced Vegetation Index (EVI) | [47] | 0.706 | 0.018 | Exponential | y = e(5.89928+5.66156x) |
| Sampling Site | Aboveground Biomass | Coordinates | ||
|---|---|---|---|---|
| Fresh Matter (g m−2) | AFDB (g m−2) | N | E | |
| 1 | 3219.6 | 2289.1 | 44.56938 | 11.83117 |
| 2 | 2649.6 | 1739.5 | 44.57368 | 11.81999 |
| 3 | 1639.8 | 1056.1 | 44.57347 | 11.82222 |
| 4 | 1178.1 | 715.1 | 44.57371 | 11.82299 |
| 5 | 2166.4 | 1337.3 | 44.57289 | 11.81776 |
| 6 | 1112.4 | 667.7 | 44.57372 | 11.82539 |
| 7 | 3744.5 | 2429.7 | 44.57388 | 11.82720 |
| C Sequestration | C Storage | |||||||
|---|---|---|---|---|---|---|---|---|
| Year | g AFDB m−2 | g C m−2 | g C m−2 yr−1 | t C yr−1 | € 103 yr−1 | t AFDB | t C | € 103 |
| 1985 | 1495.9 | 925.7 | 327.0 | 814.3 | 78.9 | 3724.9 | 2305.0 | 223.4 |
| 1989 | 2492.5 | 1542.3 | 544.9 | 1356.8 | 131.5 | 6206.3 | 3840.4 | 372.3 |
| 1997 | 2069.0 | 1280.3 | 452.3 | 1126.3 | 109.2 | 5151.7 | 3187.9 | 309.0 |
| 2010 | 1570.5 | 971.8 | 343.3 | 854.9 | 82.9 | 3910.5 | 2419.8 | 234.6 |
| 2016 | 967.7 | 598.8 | 211.5 | 526.8 | 51.0 | 2409.5 | 1491.0 | 144.5 |
| 2017 | 1270.4 | 786.1 | 277.7 | 691.5 | 67.0 | 3163.3 | 1957.4 | 189.7 |
| 2018 | 1139.1 | 704.9 | 249.0 | 620.1 | 60.1 | 2836.4 | 1755.2 | 170.1 |
| Unit | E | N-V | V | |
|---|---|---|---|---|
| NH4+ | mg/L | 0.14 (±0.03) | 0.07 (±0.01) | 0.13 (±0.01) |
| NO2− | mg/L | 0.12 (±0.03) | 0.09 (±0.03) | 0.09 (±0.03) |
| NO3− | mg/L | 4.27 (±1.11) | 2.45 (±1.00) | 2.41 (±0.93) |
| PO43− | mg/L | 0.01 (±0.001) | 0.01 (±0.001) | 0.01 (±0.001) |
| TSS | mg/L | 69.15 (±8.75) | 71.44 (±9.66) | 147.27 (±17.06) |
| ISS | mg/L | 54.18 (±7.49) | 52.70 (±7.03) | 117.18 (±14.35) |
| OSS | mg/L | 14.97 (±1.52) | 18.75 (±2.90) | 30.08 (±2.93) |
| O2 | mg/L | 8.63 (±0.70) | 9.70 (±0.75) | 8.54 (±0.47) |
| pH | - | 7.30 (±0.09) | 7.60 (±0.05) | 7.60 (±0.04) |
| Temp | °C | 11.30 (±1.53) | 11.60 (±1.54) | 11.70 (±1.38) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaglio, M.; Bresciani, M.; Ghirardi, N.; Muresan, A.N.; Lanzoni, M.; Vincenzi, F.; Castaldelli, G.; Fano, E.A. Aquatic Vegetation Loss and Its Implication on Climate Regulation in a Protected Freshwater Wetland of Po River Delta Park (Italy). Water 2022, 14, 117. https://doi.org/10.3390/w14010117
Gaglio M, Bresciani M, Ghirardi N, Muresan AN, Lanzoni M, Vincenzi F, Castaldelli G, Fano EA. Aquatic Vegetation Loss and Its Implication on Climate Regulation in a Protected Freshwater Wetland of Po River Delta Park (Italy). Water. 2022; 14(1):117. https://doi.org/10.3390/w14010117
Chicago/Turabian StyleGaglio, Mattias, Mariano Bresciani, Nicola Ghirardi, Alexandra Nicoleta Muresan, Mattia Lanzoni, Fabio Vincenzi, Giuseppe Castaldelli, and Elisa Anna Fano. 2022. "Aquatic Vegetation Loss and Its Implication on Climate Regulation in a Protected Freshwater Wetland of Po River Delta Park (Italy)" Water 14, no. 1: 117. https://doi.org/10.3390/w14010117
APA StyleGaglio, M., Bresciani, M., Ghirardi, N., Muresan, A. N., Lanzoni, M., Vincenzi, F., Castaldelli, G., & Fano, E. A. (2022). Aquatic Vegetation Loss and Its Implication on Climate Regulation in a Protected Freshwater Wetland of Po River Delta Park (Italy). Water, 14(1), 117. https://doi.org/10.3390/w14010117