
National-Scale Assessment of Climate Change Impacts on Two Native Freshwater Fish Using a Habitat Suitability Model


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Data Collection
2.2. Regression Models for Hydraulic and Thermal Variables
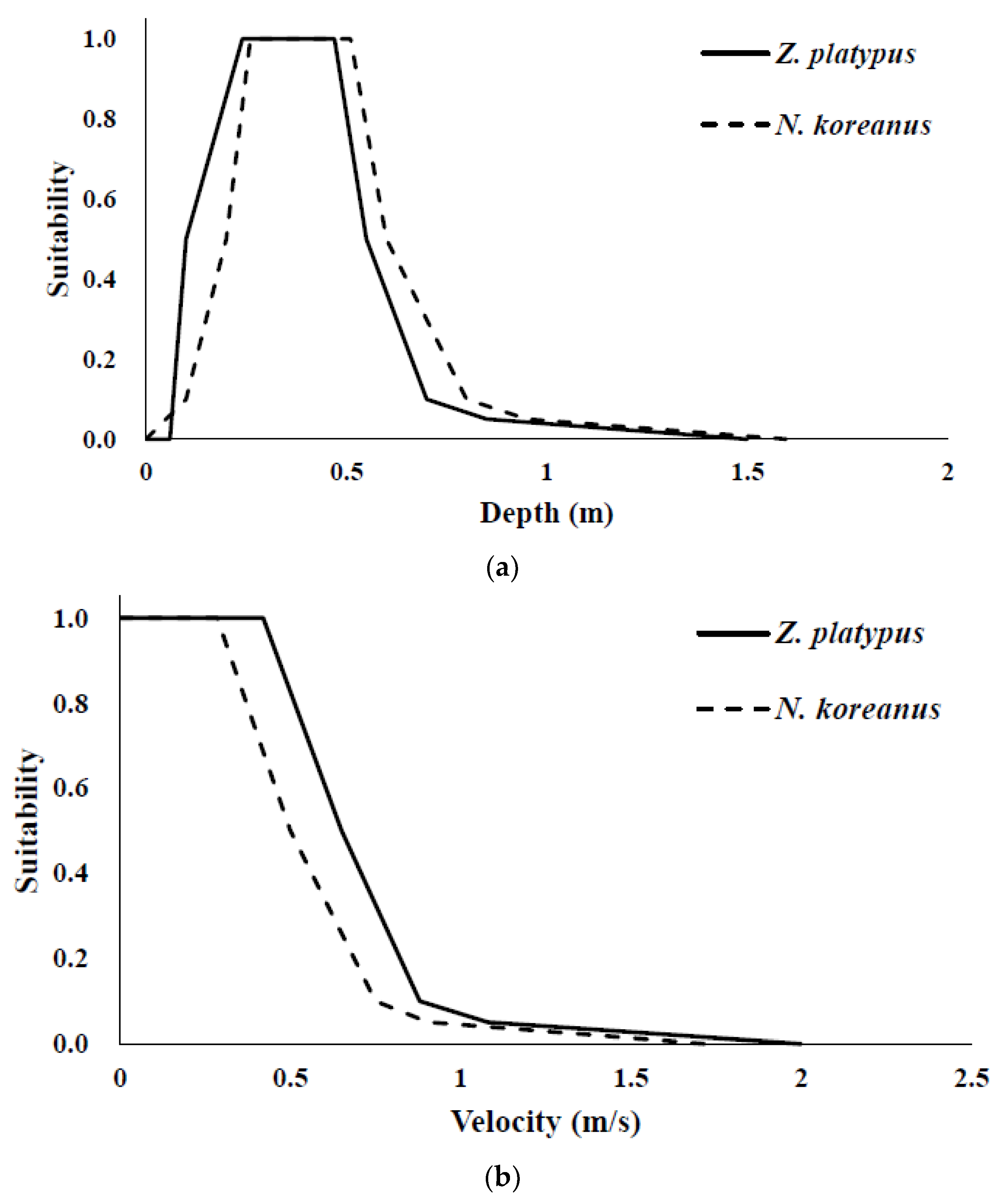
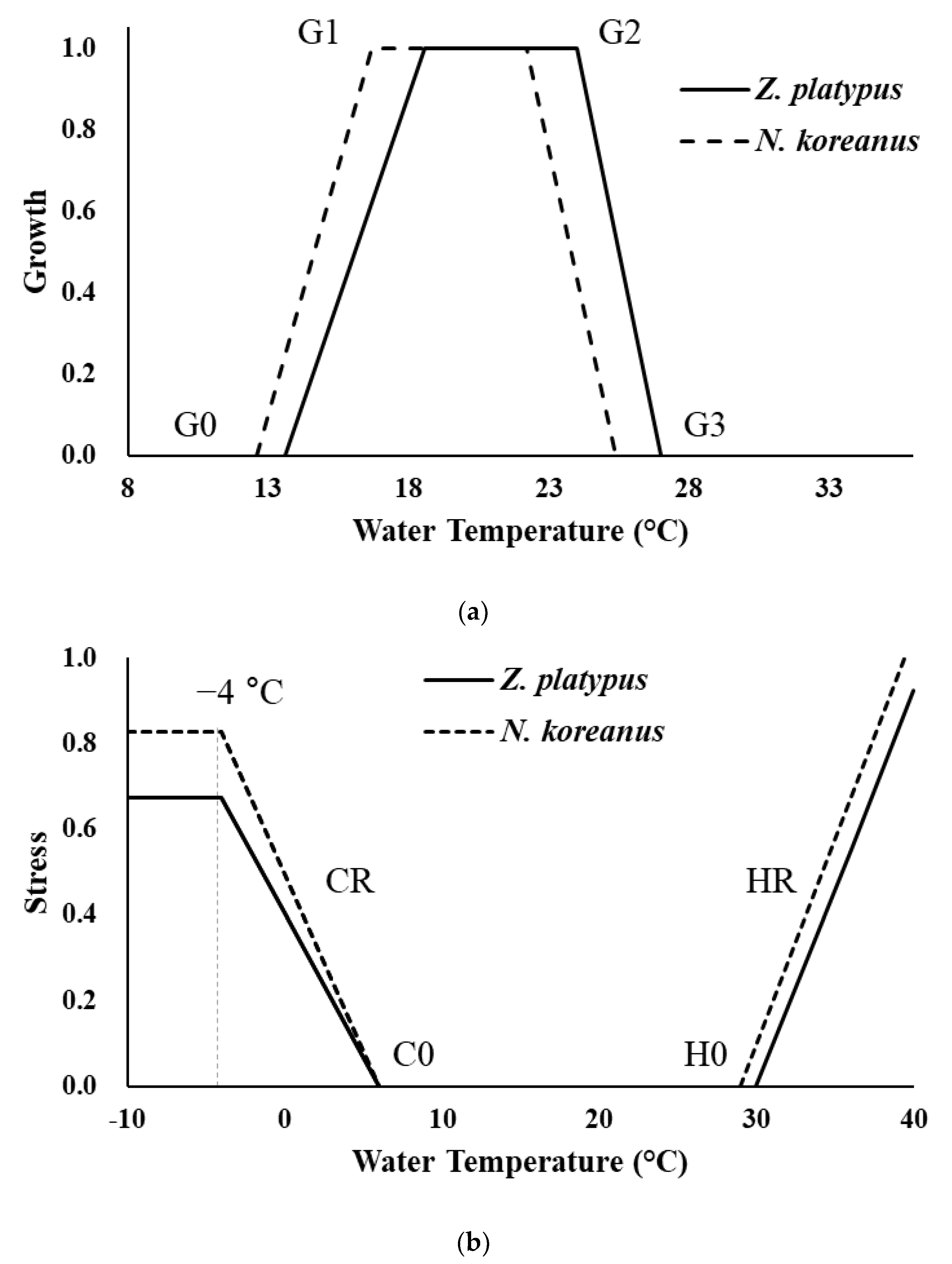
2.3. Response Curves for Hydraulic and Physiologic Habitat Suitability
2.4. Habitat Suitability Modeling
2.5. Habitat Suitability Model Validation
2.6. Statistical Analysis
3. Results and Discussion
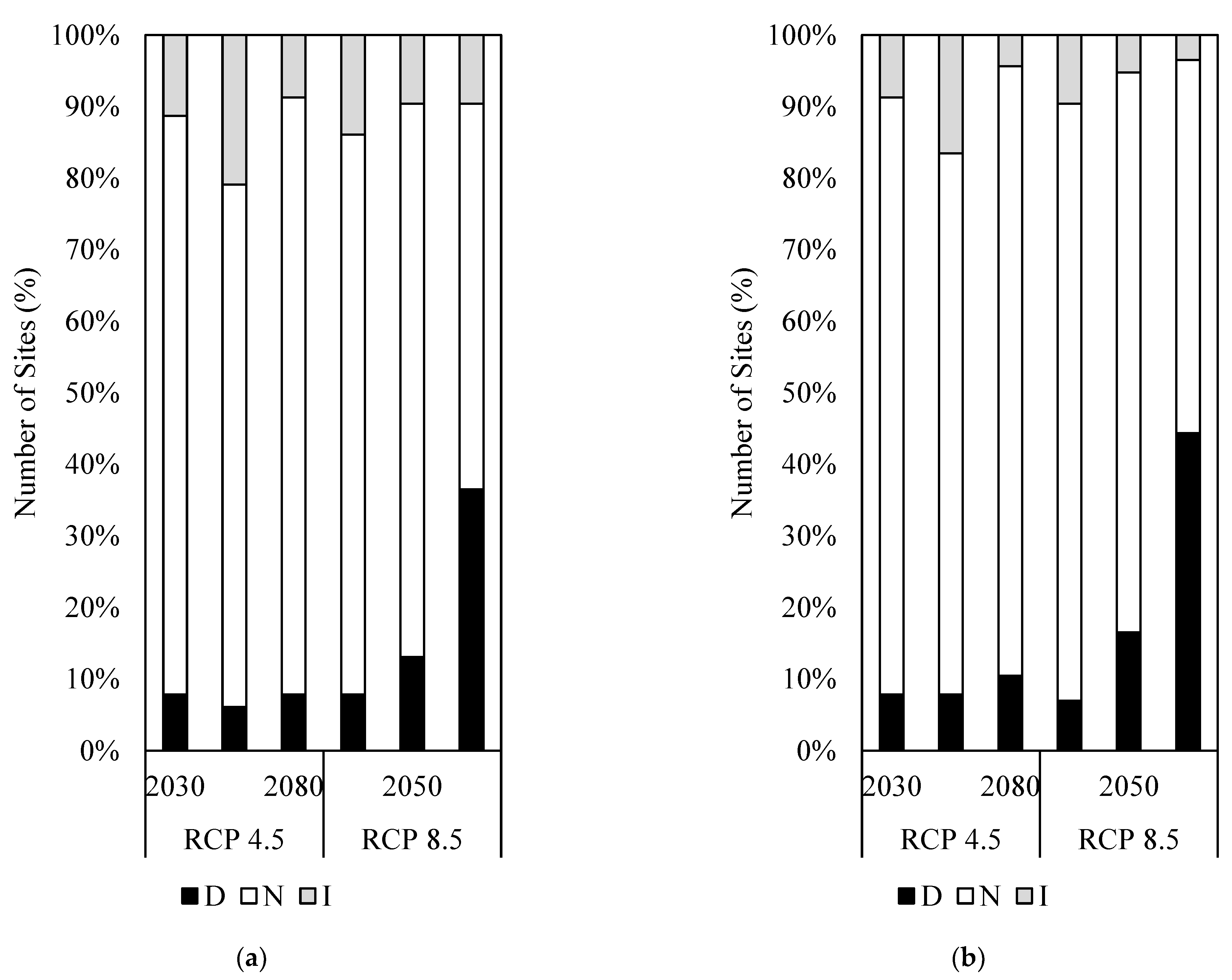
3.1. Impact of Climate Change on Habitat Suitability
3.2. Identification of Major Environmental Factors
3.3. Implications and Limitations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Isaak, D.J.; Wollrab, S.; Horan, D.; Chandler, G. Climate change effects on stream and river temperatures across the northwest US from 1980–2009 and implications for salmonid fishes. Clim. Chang. 2012, 113, 499–524. [Google Scholar] [CrossRef] [Green Version]
- Webb, B.W.; Nobilis, F. Long-term changes in river temperature and the influence of climatic and hydrological factors. Hydol. Sci. 2007, 52, 74–85. [Google Scholar] [CrossRef]
- Fraser, G.S.; Bestgen, K.R.; Winkelman, D.L.; Thompson, K.G. Temperature—Not flow—Predicts native fish reproduction with implications for climate change. Trans. Am. Fish. Soc. 2019, 148, 509–527. [Google Scholar] [CrossRef]
- Olden, J.D.; Naiman, R.J. Incorporating thermal regimes into environmental flows assessments: Modifying dam operations to restore freshwater ecosystem integrity. Freshw. Biol. 2010, 55, 86–107. [Google Scholar] [CrossRef]
- Wenger, S.J.; Isaak, D.J.; Luce, C.H.; Neville, H.M.; Fausch, K.D.; Dunham, J.B.; Dauwalter, D.C.; Young, M.K.; Elsner, M.M.; Rieman, B.E.; et al. Flow regime, temperature, and biotic interactions drive differential declines of trout species under climate change. Proc. Natl. Acad. Sci. USA 2011, 108, 14175–14180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, A.K.; Taylor, W.W.; Schlee, K.M.; Zorn, T.G.; Infante, D.M. Projected impacts of climate change on stream salmonids with implications for resilience-based management. Ecol. Freshw. Fish 2017, 26, 190–204. [Google Scholar] [CrossRef]
- Mantua, N.; Tohver, I.; Hamlet, A. Climate change impacts on streamflow extremes and summertime stream temperature and their possible consequences for freshwater salmon habitat in Washington State. Clim. Chang. 2010, 102, 187–223. [Google Scholar] [CrossRef]
- Poesch, M.S.; Chavarie, L.; Chu, C.; Pandit, S.N.; Tonn, W. Climate change impacts on freshwater fishes: A Canadian perspective. Fisheries 2016, 41, 385–391. [Google Scholar] [CrossRef]
- Bovee, K.D. A Guide to Stream Habitat Analysis Using the Instream Flow Incremental Methodology. Instream Flow Information Paper, No. 12; FWS/OBS–82/26; U.S. Fish and Wildlife Service: Fort Collins, CO, USA, 1982. [Google Scholar]
- Gore, J.A. The Restoration of Rivers and Streams: Theories and Experience; Butterworth Publishers: Boston, MA, USA, 1985. [Google Scholar]
- Bush, A.A.; Nipperess, D.A.; Duursma, D.E.; Theischinger, G.; Turak, E.; Hughes, L. Continental-scale assessment of risk to the Australian Odonata from climate change. PLoS ONE 2014, 9, e88958. [Google Scholar] [CrossRef] [Green Version]
- Kim, Z.; Shim, T.; Koo, Y.-M.; Seo, D.; Kim, Y.-O.; Hwang, S.-J.; Jung, J. Predicting the impact of climate change on freshwater fish distribution by incorporating water flow rate and quality variables. Sustainability 2020, 12, 10001. [Google Scholar] [CrossRef]
- Markovic, D.; Freyhof, J.; Wolter, C. Where are all the fish: Potential of biogeographical maps to project current and future distribution patterns of freshwater species. PLoS ONE 2012, 7, e40530. [Google Scholar] [CrossRef] [PubMed]
- Bovee, K.D.; Lamb, B.L.; Bartholow, J.M.; Stalnaker, C.B.; Taylor, J.; Henriksen, J. Stream Habitat Analysis Using the Instream Flow Incremental Methodology; Information and Technology Report USGS/BRD-1998-0004; U.S. Geological Survey, Biological Resources Division: Fort Collin, CO, USA, 1998; p. 131. [Google Scholar]
- Choi, S.-U.; Kim, S.K.; Choi, B.; Kim, Y. Impact of hydropeaking on downstream fish habitat at the Goesan Dam in Korea. Ecohydrology 2017, 10, e1861. [Google Scholar] [CrossRef]
- Choi, B.; Choi, S.-U. Impacts of hydropeaking and thermopeaking on the downstream habitat in the Dal River, Korea. Ecol. Inf. 2018, 43, 1–11. [Google Scholar] [CrossRef]
- Im, D.; Kang, H.; Kim, K.-H.; Choi, S.-U. Changes of river morphology and physical fish habitat following weir removal. Ecol. Eng. 2011, 37, 883–892. [Google Scholar]
- Kim, S.K.; Choi, S.-U. Evaluation of the impact of abandoned channel restoration on Zaco platypus habitat using the physical habitat simulation: A case study of the Cheongmi-cheon stream in Korea. Ecol. Resil. Infrastruct. 2019, 6, 101–108. [Google Scholar]
- Yi, Y.; Cheng, X.; Wieprecht, S.; Tang, C. Comparison of habitat suitability models using different habitat suitability evaluation methods. Ecol. Eng. 2014, 71, 335–345. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, Z.; Cai, L.; Qiao, Y.; Chen, X.; Chang, J. Effects of upstream and downstream dam operation on spawning habitat suitability of Coreius guichenoti in the middle reach of the Jinsha River. Ecol. Eng. 2018, 120, 198–208. [Google Scholar] [CrossRef]
- Macura, V.; Štefunková, Z.; Škrinár, A. Determination of the effect of water depth and flow velocity on the quality of an in-stream habitat in terms of climate change. Adv. Meteorol. 2016, 2016, 4560378. [Google Scholar] [CrossRef] [Green Version]
- Papadaki, C.; Soulis, K.; Muñoz–Mas, R.; Martinez–Capel, F.; Zogaris, S.; Ntoanidis, L.; Dimitriou, E. Potential impacts of climate change on flow regime and fish habitat in mountain rivers of the south-western Balkans. Sci. Total Environ. 2016, 540, 418–428. [Google Scholar] [CrossRef] [Green Version]
- Walsh, C.L.; Kilsby, C.G. Implications of climate change on flow regime affecting Atlantic salmon. Hydrol. Earth Syst. Sci. 2007, 11, 1127–1143. [Google Scholar] [CrossRef]
- Beaupré, J.; Boudreault, J.; Bergeron, N.E.; St–Hilaire, A. Inclusion of water temperature in a fuzzy logic Atlantic salmon (Salmo salar) parr habitat model. J. Therm. Biol. 2020, 87, 102471. [Google Scholar] [CrossRef]
- Morid, R.; Shimatani, Y.; Sato, T. An integrated framework for prediction of climate change impact on habitat suitability of a river in terms of water temperature, hydrological and hydraulic parameters. J. Hydrol. 2020, 587, 124936. [Google Scholar] [CrossRef]
- Muñoz–Mas, R.; Marcos–Garcia, P.; Lopez–Nicolas, A.; Martínez–García, F.J.; Pulido–Velazquez, M.; Martínez–Capel, F. Combining literature–based and data–driven fuzzy models to predict brown trout (Salmo trutta L.) spawning habitat degradation induced by climate change. Ecol. Model. 2018, 386, 98–114. [Google Scholar] [CrossRef]
- Zhang, P.; Qiao, Y.; Schneider, M.; Chang, J.; Mutzner, R.; Fluixá–Sanmartín, J.; Yang, Z.; Fu, R.; Chen, X.; Cai, L.; et al. Using a hierarchical model framework to assess climate change and hydropower operation impacts on the habitat of an imperiled fish in the Jinsha river, China. Sci. Total Environ. 2019, 646, 1624–1638. [Google Scholar] [CrossRef]
- Beechie, T.; Imaki, H.; Greene, J.; Wade, A.; Wu, H.; Roni, P.; Kimball, J.; Stanford, J.; Kiffney, P.; Mantua, N. Restoring salmon habitat for a changing climate. River Res. Appl. 2013, 29, 939–960. [Google Scholar] [CrossRef]
- Justice, C.; White, S.M.; McCullough, D.A.; Graves, D.S.; Blanchard, M.R. Can stream and riparian restoration offset climate change impacts to salmon populations? J. Environ. Manag. 2017, 188, 212–227. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, S.S.; Likens, G.E.; Jaworski, N.A.; Pace, M.L.; Sides, A.M.; Seekell, D.; Belt, K.T.; Secor, D.H.; Wingate, R.L. Rising stream and river temperatures in the United States. Front. Ecol. Environ. 2010, 8, 461–466. [Google Scholar] [CrossRef]
- Wondzell, S.M.; Diabat, M.; Haggerty, R. What matters most: Are future stream temperatures more sensitive to changing air temperatures, discharge, or riparian vegetation? J. Am. Water Resour. Assoc. 2019, 55, 116–132. [Google Scholar] [CrossRef] [Green Version]
- Mathur, D.; Bason, W.; Purdy, E.J., Jr.; Silver, C.A. A critique of the instream flow incremental methodology. Can. J. Fish Aquat. Sci. 1984, 42, 825–831. [Google Scholar] [CrossRef]
- Shim, T.; Kim, Z.; Seo, D.; Kim, Y.-O.; Hwang, S.-J.; Jung, J. Integrating hydraulic and physiologic factors to develop an ecological habitat suitability model. Environ. Model. Softw. 2020, 131, 104760. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Maywald, G.F.; Yonow, T.; Zurcher, E.J.; Herrmann, N.I.; Sutherst, R.W. CLIMEX Version 4: Exploring the Effects of Climate on Plants, Animals and Diseases; Commonwealth Scientific and Industrial Research Organisation: Canberra, Australia, 2015. [Google Scholar]
- Yoon, J.-D.; Kim, J.-H.; Park, S.-H.; Jang, M.-H. The distribution and diversity of freshwater fishes in the Korean peninsula. Korean J. Ecol. Environ. 2018, 51, 71–85. [Google Scholar] [CrossRef]
- Lee, W.-O.; Noh, S.-Y. Freshwater Fishes in Korean Peninsula with Looking Characteristics; Jisung Publishing: Seoul, Korea, 2007; p. 432. [Google Scholar]
- Lee, S.-H.; Jeong, H.-G.; Shin, H.-S.; Shin, Y.; Lee, S.-W.; Lee, J.-K. Comparison on ecological index characteristics between Zacco platypus and Zacco koreanus by stream order in Korea. Korean J. Ecol. Environ. 2017, 50, 403–410. [Google Scholar] [CrossRef]
- Lee, S.-W.; Hwang, S.J.; Lee, J.-K.; Jung, D.-I.; Park, Y.-J.; Kim, J.-T. Overview and application of the National Aquatic Ecological Monitoring Program (NAEMP) in Korea Ann. Limnol. Int. J. Limnol. 2011, 47, S3–S14. [Google Scholar] [CrossRef]
- NIER. Survey and Assessment of Stream/River Ecosystem Health (VII); Publication Number: 11-1480523-002181-01; NIER: Incheon, Korea, 2014. [Google Scholar]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Jang, Y.; Park, J.; Seo, D. Estimations of flow rate and pollutant loading changes of the Yo-Cheon basin under AR5 climate change scenarios using SWAT. J. Korean Soc. Water Wastewater 2018, 32, 221–233. [Google Scholar] [CrossRef]
- Park, J.; Jang, Y.; Seo, D. Water quality prediction of inflow of the Yongdam Dam basin and its reservoir using SWAT and CE-QUAL-W2 models in series to climate change scenarios. J. Korea Water Resourc. Assoc. 2017, 50, 703–714. [Google Scholar]
- Moriasi, D.N.; Arnold, J.G.; Van Liew, M.W.; Bingner, R.L.; Harmel, R.D.; Veith, T.L. Model evaluation guidelines for systematic quantification of accuracy in watershed simulations. Trans. ASABE 2007, 50, 885–900. [Google Scholar] [CrossRef]
- Santhi, C.; Arnold, J.G.; Williams, J.R.; Dugas, W.A.; Srinivasan, R.; Hauk, L.M. Validation of the SWAT model on a large river basin with point and nonpoint sources. J. Am. Water Resour. Assoc. 2001, 37, 1169–1188. [Google Scholar] [CrossRef]
- Waddle, T.J. (Ed.) PHABSIM for Windows: User’s Manual and Exercises. Open–File Report 01–340; US Geological Survey: Fort Collins, CO, USA, 2001. [Google Scholar]
- Neitsch, S.L.; Arnold, J.G.; Kiniry, J.R.; Williams, J.R. Soil and Water Assessment Tool Theoretical Documentation Version 2009; Texas Water Resources Institute, Texas A&M University: College Station, TX, USA, 2011; p. 60. [Google Scholar]
- Kang, H. Development of Physical Fish Habitat Suitability Index; Korea Environment Institute: Seoul, Korea, 2010; p. 60. [Google Scholar]
- Gosse, J.C. Microhabitat of Rainbow and Cutthroat Trout in the Green River Below Flaming Gorge Dam; Final Report, Contract 81-5049; Utah Division of Wildlife Resources: Salt Lake City, UT, USA, 1982; p. 114. [Google Scholar]
- IFASG (Instream Flow and Aquatic Systems Group). Development and Evaluation of Habitat Suitability Criteria for Use in the Instream Flow Incremental Methodology: Biologic Report. Instream Flow Information Paper No. 21; U.S. Fish & Wildlife Service: Fort Collins, CO, USA, 1986. [Google Scholar]
- Chung, N.; Park, B.; Kim, K. Potential effect of increased water temperature on fish habitats in Han river watershed. J. Korean Soc. Water Environ. 2011, 24, 314–321. [Google Scholar]
- Kang, H.; Park, M.-Y.; Jang, J.-H. Effect of climate change on fish habitat in the Nakdong river watershed. J. Korea Water Resour. Assoc. 2013, 46, 1–12. [Google Scholar] [CrossRef] [Green Version]
- NIER (National Institute of Environmental Research). Distribution Patterns of Aquatic Animals with Water Environment; Publication Number: 11-1480523-000094-01; NIER: Incheon, Korea, 2006. [Google Scholar]
- Ahmadi–Nedushan, B.; St–Hilaire, A.; Bérubé, M.; Robichaud, É.; Thiémonge, N.; Bobée, B. A review of statistical methods for the evaluation of aquatic habitat suitability for instream flow assessment. River Res. Appl. 2006, 22, 503–523. [Google Scholar] [CrossRef]
- Fukuda, S.; De Baets, B.; Mouton, A.M.; Waegeman, W.; Nakajima, J.; Mukai, T.; Hiramatsu, K.; Onikura, N. Effect of model formulation on the optimization of a Takagi-Sugeno fuzzy system for fish habitat suitability evaluation. Ecol. Model. 2011, 222, 1401–1413. [Google Scholar] [CrossRef]
- Jenks, G.G. The data model concept in statistical mapping. Int. J. Cart. 1967, 7, 186–190. [Google Scholar]
- Alemany, D.; Iribarne, O.O.; Acha, E.M. Effects of a large-scale and offshore marine protected area on the demersal fish assemblage in the Southwest Atlantic. ICES J. Mar. Sci. 2013, 70, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Van Der Wal, J.; Shoo, L.P.; Johnson, C.N.; Williams, S.E. Abundance and the environmental niche: Environmental suitability estimated from niche models predicts the upper limit of local abundance. Am. Nat. 2009, 174, 282–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayllón, D.; Railsback, S.F.; Harvey, B.C.; Quirós, I.G.; Nicola, G.G.; Elvira, B.; Almodóvar, A. Mechanistic simulations predict that thermal and hydrological effects of climate change on Mediterranean trout cannot be offset by adaptive behavior, evolution, and increased food production. Sci. Total Environ. 2019, 693, 33648. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, A.L. Species diversity and longitudinal succession in stream fishes. Ecology 1968, 49, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Vismara, R.; Azzellino, A.; Bosi, R.; Crosa, G.; Gentili, G. Habitat suitability curves for brown trout (Salmo trutta fario L.) in the River Adda, Northern Italy: Comparing univariate and multivariate approaches. Regul. Rivers Res. Manag. 2001, 17, 37–50. [Google Scholar] [CrossRef]
- Xu, C.; Gertner, G.Z. Uncertainty and sensitivity analysis for models with correlated parameters. Reliab. Eng. Syst. Saf. 2008, 93, 563–1573. [Google Scholar] [CrossRef]
- Morales-Marín, L.A.; Rokay, P.; Sanyal, P.R.; Sereda, J.; Lindenschmidt, K.E. Changes in streamflow and water temperature affect fish habitat in the Athabasca River basin in the context of climate change. Ecol. Model. 2019, 407, 08718. [Google Scholar] [CrossRef]
- Chadwick, J.G.; McCormick, S.D. Upper thermal limits of growth in brook trout and their relationship to stress physiology. J. Exp. Biol. 2017, 220, 976–3987. [Google Scholar] [CrossRef] [Green Version]
- Butryn, R.S.; Parrish, D.L.; Rizzo, D.M. Summer stream temperature metrics for predicting brook trout (Salvelinus fontinalis) distribution in streams. Hydrobiologia 2013, 703, 47–57. [Google Scholar] [CrossRef]
- Kangur, A.; Kangur, P.; Kangur, K.; Möls, T. The role of temperature in the population dynamics of smelt Osmerus eperlanus eperlanus m. spirinchus Pallas in Lake Peipsi (Estonia/Russia). Hydrobiologia 2007, 584, 433–441. [Google Scholar] [CrossRef]
- Elliott, J.M.; Elliott, J.A. Temperature requirements of Atlantic salmon Salmo salar, brown trout Salmo trutta and Arctic charr Salvelinus alpinus: Predicting the effects of climate change. J. Fish. Biol. 2010, 77, 1793–1817. [Google Scholar] [CrossRef] [PubMed]
- Ficke, A.D.; Myrick, C.A.; Hansen, L.J. Potential impacts of global climate change on freshwater fisheries. Rev. Fish Biol. Fish. 2007, 17, 581–613. [Google Scholar] [CrossRef]
- Hari, R.; Livingstone, D.M.; Siber, R.; Burkhardt–Holm, P.; Güttinger, H. Consequences of climatic change for water temperature and brown trout populations in Alpine rivers and streams. Glob. Chang. Biol. 2006, 12, 20–26. [Google Scholar] [CrossRef]
- Hasnain, S.S.; Minns, C.K.; Shuter, B.J. Key Ecological Temperature Metrics for Canadian Freshwater Fishes; Ontario Forest Research Institute: Sault Ste. Marie, ON, Canada, 2010. [Google Scholar]
- Kling, G.W.; Hayhoe, K.; Johnson, L.B.; Magnuson, J.J.; Polasky, S.; Robinson, S.K.; Shuter, B.J.; Wander, M.M.; Wuebbles, D.J.; Zack, D.R. Confronting Climate Change in the Great Lakes Region: Impacts on Our Communities and Ecosystems; Union of Concerned Scientists: Cambridge, MA, USA; Ecological Society of America: Washington, DC, USA, 2003; p. 92. [Google Scholar]
- Sutherst, R.W. Implications of global change and climate variability for vector–borne diseases: Genertic approaches to impact assessments. Int. J. Parasitol. 1998, 28, 935–945. [Google Scholar] [CrossRef]
- Kearney, M.; Porter, W. Mechanistic niche modelling: Combining physiological and spatial data to predict species’ ranges. Ecol. Lett. 2009, 12, 334–350. [Google Scholar] [CrossRef]
- Li, Y.; Blazer, V.S.; Wagner, T. Quantifying population-level effects of water temperature, flow velocity and chemical–induced reproduction depression: A simulation study with smallmouth bass. Ecol. Model. 2018, 384, 63–74. [Google Scholar] [CrossRef]
- Selong, J.H.; McMahon, T.E.; Zale, A.V.; Barrows, F.T. Effect of temperature on growth and survival of bull trout with application of an improved method for determining thermal tolerance in fishes. Trans. Am. Fish. Soc. 2001, 130, 1026–1037. [Google Scholar] [CrossRef]
- Widmer, A.M.; Carveth, C.J.; Bonar, S.A.; Simms, J.R. Upper temperature tolerance of loach minnow under acute, chronic, and fluctuating thermal regimes. Trans. Am. Fish. Soc. 2006, 135, 755–762. [Google Scholar] [CrossRef]
- Muñoz–Mas, R.; Lopez–Nicolas, A.; Martínez–Capel, F.; Pulido–Velazquez, M. Shifts in the suitable habitat available for brown trout (Salmo trutta L.) under short-term climate change scenarios. Sci. Total Environ. 2016, 544, 686–700. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scenario | Period | Z. platypus | N. koreanus |
|---|---|---|---|
| RCP 4.5 | Present | 0.313 ± 0.149 | 0.260 ± 0.129 |
| 2030 | 0.319 ± 0.159 (+1.92%) | 0.259 ± 0.136 (−0.385%) | |
| 2050 | 0.333 ± 0.163 (+6.39%) | 0.275 ± 0.139 (+5.77%) | |
| 2080 | 0.322 ± 0.161 (+2.88%) | 0.261 ± 0.136 (+0.385%) | |
| RCP 8.5 | Present | 0.314 ± 0.150 | 0.261 ± 0.131 |
| 2030 | 0.326 ± 0.161 (+3.82%) | 0.267 ± 0.137 (+2.30%) | |
| 2050 | 0.308 ± 0.159 (−1.91%) | 0.245 ± 0.138 (−6.13%) | |
| 2080 | 0.283 ± 0.163 (−9.87%) | 0.211 ± 0.144 (−19.2%) |
| Total | ΔFlowavg | ΔFlowmin | ΔFlowmax | ΔDepth | ΔVelocity | ΔWTavg | ΔWTmin | ΔWTmax | ΔCold | ΔHot |
|---|---|---|---|---|---|---|---|---|---|---|
| Z. platypus | −0.009 | 0.011 | −0.045 | −0.129 * | 0.043 | −0.095 * | 0.110 * | −0.054 | −0.069 | −0.314 * |
| N. koreanus | 0.038 | 0.047 | −0.033 | −0.085 * | 0.035 | −0.101 * | 0.117 * | −0.065 | −0.105 * | −0.326 * |
| Hydraulic | ΔFlowavg (cms) | ΔFlowmin (cms) | ΔFlowmax (cms) | ΔDepth (m) | ΔVelocity (m/s) | ||
|---|---|---|---|---|---|---|---|
| RCP4.5 | 2030 | 3.94 ± 13.5 | −0.022 ± 0.131 | 146 ± 472 | 0.406 ± 0.823 | 0.007 ± 0.069 | |
| 2050 | 6.48 ± 11.5 | −0.010 ± 0.085 | 251 ± 478 | 0.432 ± 0.830 | 0.023 ± 0.035 | ||
| 2080 | 9.86 ± 18.8 | −0.019 ± 0.111 | 585 ± 1330 | 0.434 ± 0.833 | 0.025 ± 0.037 | ||
| RCP8.5 | 2030 | 7.25 ± 14.5 | −0.016 ± 0.106 | 281 ± 514 | 0.059 ± 0.307 | 0.012 ± 0.040 | |
| 2050 | 4.87 ± 9.69 | −0.022 ± 0.210 | 121 ± 494 | 0.061 ± 0.309 | 0.013 ± 0.036 | ||
| 2080 | 7.04 ± 13.5 | −0.028 ± 0.190 | 101 ± 316 | 0.073 ± 0.312 | 0.022 ± 0.037 | ||
| Physiologic | ΔTavg (°C) | ΔTmin (°C) | ΔTmax (°C) | ΔCold (d) | ΔHot (d) * | ΔHot (d) ** | |
| RCP 4.5 | 2030 | 0.304 ± 0.305 | 0.163 ± 0.391 | 0.485 ± 0.318 | 0.248 ± 9.02 | −13.3 ± 14.2 | −12.7 ± 13.4 |
| 2050 | 0.785 ± 0.339 | 0.744 ± 0.407 | 0.867 ± 0.345 | −5.89 ± 9.52 | −8.47 ± 14.7 | −7.68 ± 15.2 | |
| 2080 | 1.21 ± 0.359 | 1.21 ± 0.430 | 1.28 ± 0.342 | −11.4 ± 9.98 | 0.650 ± 12.9 | 3.21 ± 14.4 | |
| RCP 8.5 | 2030 | 0.378 ± 0.331 | 0.244 ± 0.468 | 0.474 ± 0.567 | −2.81 ± 11.1 | −13.5 ± 14.5 | −11.9 ± 14.4 |
| 2050 | 1.16 ± 0.376 | 1.04 ± 0.522 | 1.23 ± 0.596 | −8.61 ± 11.2 | 6.07 ± 12.8 | 8.94 ± 14.7 | |
| 2080 | 2.37 ± 0.473 | 2.38 ± 0.575 | 2.43 ± 0.462 | −24.2 ± 13.9 | 33.0 ± 17.2 | 36.7 ± 17.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shim, T.; Kim, Z.; Seo, D.; Jung, J. National-Scale Assessment of Climate Change Impacts on Two Native Freshwater Fish Using a Habitat Suitability Model. Water 2022, 14, 1825. https://doi.org/10.3390/w14111825
Shim T, Kim Z, Seo D, Jung J. National-Scale Assessment of Climate Change Impacts on Two Native Freshwater Fish Using a Habitat Suitability Model. Water. 2022; 14(11):1825. https://doi.org/10.3390/w14111825
Chicago/Turabian StyleShim, Taeyong, Zhonghyun Kim, Dongil Seo, and Jinho Jung. 2022. "National-Scale Assessment of Climate Change Impacts on Two Native Freshwater Fish Using a Habitat Suitability Model" Water 14, no. 11: 1825. https://doi.org/10.3390/w14111825
APA StyleShim, T., Kim, Z., Seo, D., & Jung, J. (2022). National-Scale Assessment of Climate Change Impacts on Two Native Freshwater Fish Using a Habitat Suitability Model. Water, 14(11), 1825. https://doi.org/10.3390/w14111825