
The Geochemical Drivers of Bacterial Community Diversity in the Watershed Sediments of the Heihe River (Northern China)

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Method
2.2. Chemical and Physical Analysis of Samples
2.3. DNA Extraction and 454 Pyrotag Sequencing
2.4. Statistical Analysis
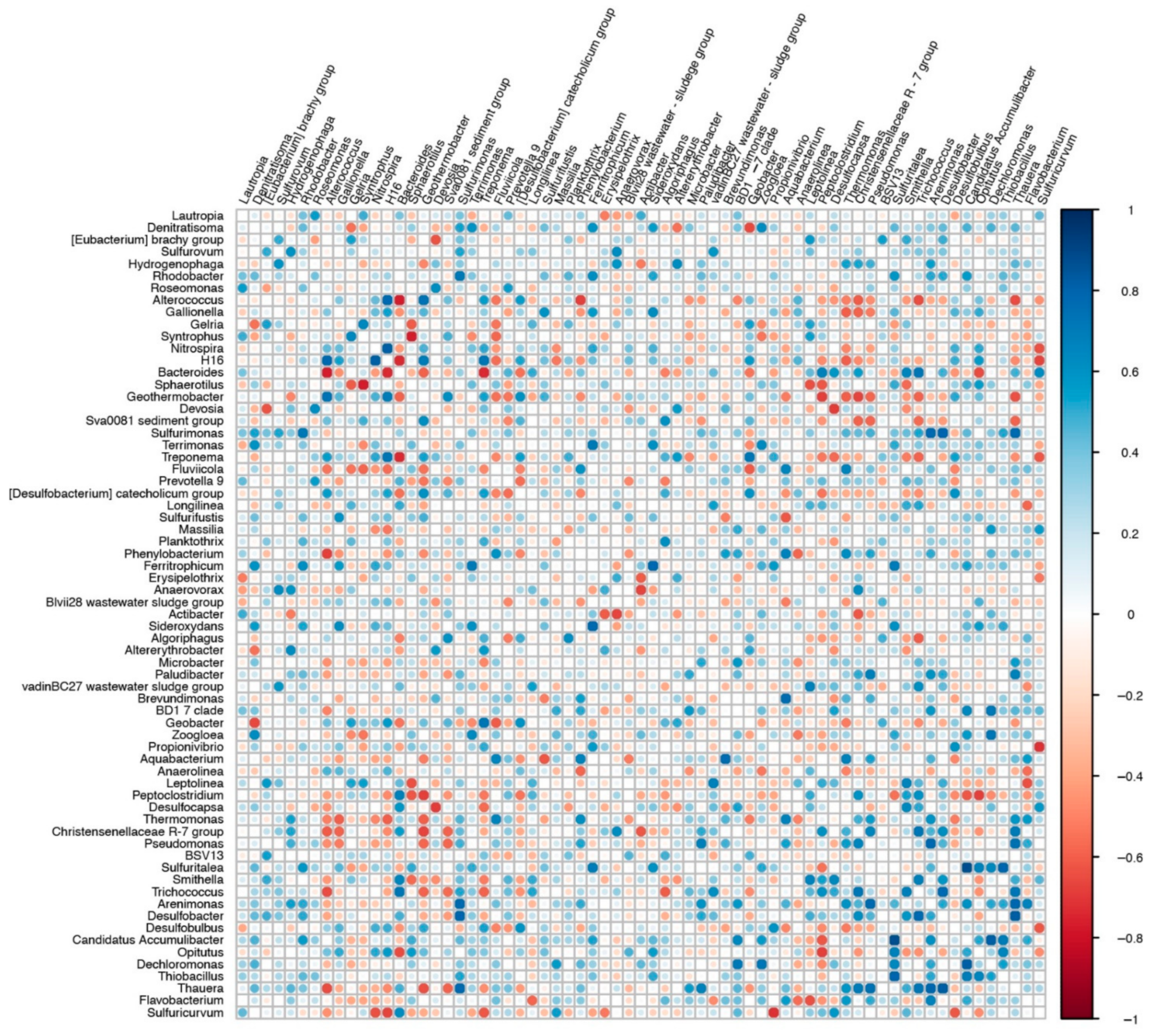
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vörösmarty, C.J.; Green, P.; Salisbury, J.; Lammers, R.B. Global Water Resources: Vulnerability from Climate Change and Population Growth. Science 2000, 289, 284–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, R. Water: The International Crisis; Routledge: Abingdon, UK, 2013; ISBN 978-1-134-15934. [Google Scholar]
- Mekonnen, M.M.; Hoekstra, A.Y. Four Billion People Facing Severe Water Scarcity. Sci. Adv. 2016, 2, e1500323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UN. Transforming Our World: The 2030 Agenda for Sustainable Development; Springer: New York, NY, USA, 2015. [Google Scholar]
- Siddig, A.A.; Ellison, A.M.; Ochs, A.; Villar-Leeman, C.; Lau, M.K. How do ecologists select and use indicator species to monitor ecological change? Insights from 14 years of publication in Ecological Indicators. Ecol. Indic. 2016, 60, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Borruso, L.; Zerbe, S.; Brusetti, L. Bacterial community structures as a diagnostic tool for watershed quality assessment. Res. Microbiol. 2015, 166, 38–44. [Google Scholar] [CrossRef]
- Garcia, S.L.; Stevens, S.L.R.; Crary, B.; Martinez-Garcia, M.; Stepanauskas, R.; Woyke, T.; Tringe, S.G.; Andersson, S.G.E.; Bertilsson, S.; Malmstrom, R.R.; et al. Contrasting patterns of genome-level diversity across distinct co-occurring bacterial populations. ISME J. 2018, 12, 742–755. [Google Scholar] [CrossRef] [Green Version]
- Jacquiod, S.; Cyriaque, V.; Riber, L.; Alsoud, W.A.; Gillan, D.C.; Wattiez, R.; Sørensen, S.J. Long-term industrial metal contamination unexpectedly shaped diversity and activity response of sediment microbiome. J. Hazard. Mater. 2018, 344, 299–307. [Google Scholar] [CrossRef]
- Burger, J. Bioindicators: A review of their use in the environmental literature 1970–2005. Environ. Bioindic. 2006, 1, 136–144. [Google Scholar] [CrossRef]
- Dickinson, A.W.; Power, A.; Hansen, M.G.; Brandt, K.K.; Piliposian, G.; Appleby, P.; O’Neill, P.A.; Jones, R.T.; Sierocinski, P.; Koskella, B.; et al. Heavy metal pollution and co-selection for antibiotic resistance: A microbial palaeontology approach. Environ. Int. 2019, 132, 105117. [Google Scholar] [CrossRef]
- Griebler, C.; Avramov, M. Groundwater ecosystem services: A review. Freshw. Sci. 2015, 34, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Achermann, S.; Mansfeldt, C.B.; Müller, M.; Johnson, D.R.; Fenner, K. Relating meta transcriptomic profiles to the micropollutant biotransformation potential of complex microbial communities. Environ. Sci. Technol. 2020, 54, 235–244. [Google Scholar] [CrossRef]
- Fakruddin, M.; Mannan, K. Methods for analyzing diversity of microbial communities in natural environments. Ceylon. J. Sci. 2013, 42, 19–33. [Google Scholar] [CrossRef]
- Lindström, E.S.; Langenheder, S. Local and regional factors influencing bacterial community assembly. Environ. Microbiol. Rep. 2012, 4, 1–9. [Google Scholar] [CrossRef]
- Cao, X.; Zhao, D.; Zheng, J.; Huang, R.; Fe, H. Biogeographic patterns of abundant and rare bacterial and microeukaryotic subcommunities in connected freshwater lake zones subjected to different levels of nutrient loading. J. Appl. Microbiol. 2020, 130, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tu, T.; Gao, G.; Bartlam, M.; Wang, Y. Biogeography and diversity of freshwater bacteria on a river catchment scale. Microb. Ecol. 2019, 78, 324–335. [Google Scholar] [CrossRef]
- Yin, X.; Chen, L.; Tang, D.; Zhang, Y.; Liu, G.; Hua, Y.; Wan, X.; Zhou, W.; Zhao, J.; Zhu, D. Seasonal and vertical variations in the characteristics of the nitrogen-related functional genes in sediments from urban eutrophic lakes. Appl. Soil. Ecol. 2019, 143, 80–88. [Google Scholar] [CrossRef]
- Ruuskanen, M.O.; St Pierre, K.A.; St Louis, V.L.; Aris-Brosou, S.; Poulain, A.J. Physicochemical drivers of microbial community structure in sediments of Lake Hazen, Nunavut, Canada. Front. Microbiol. 2018, 9, 1138. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kojima, H.; Fukui, M. Identity of major sulfur-cycle prokaryotes in freshwater lake ecosystems revealed by a comprehensive phylogenetic study of the dissimilatory adenyly lsulfate reductase. Sci. Rep. 2016, 6, 36262. [Google Scholar] [CrossRef]
- Merkley, M.; Rader, R.B.; McArthur, J.V.; Eggett, D. Bacteria as bioindicators in wetlands: Bioassessment in the Bonneville Basin of Utah, USA. Wetlands 2004, 24, 600–607. [Google Scholar] [CrossRef]
- McArthur, J.V. Bacteria as Biomonitors.Bioassessment and Management of North American Freshwater Wetlands; John Wiley & Sons. Inc.: New York, NY, USA, 2001; Volume 249. [Google Scholar]
- Zinger, L.; Gobet, A.; Pommier, T. Two decades of describing the unseen majority of aquatic microbial diversity. Mol. Ecol. 2012, 21, 1878–1896. [Google Scholar] [CrossRef]
- Zheng, B.; Wang, L.; Liu, L. Bacterial community structure and its regulating factors in the intertidal sediment along the Liaodong Bay of Bohai Sea, China. Microbiol. Res. 2014, 169, 585–592. [Google Scholar] [CrossRef]
- Magurran, A.E. Biological diversity. Curr. Biol. 2005, 15, R116–R118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClary-Gutierrez, J.S.; Driscoll, Z.; Nenn, C.; Newton, R.J. Human Fecal Contamination Corresponds to Changes in the Freshwater Bacterial Communities of a Large River Basin. Microbiol. Spectr. 2021, 9, e01200-21. [Google Scholar] [CrossRef] [PubMed]
- Mohiuddin, M.M.; Botts, S.R.; Paschos, A.; Schellhorn, H.E. Temporal and spatial changes in bacterial diversity in mixed use watersheds of the Great Lakes region. J. Gt. Lakes Res. 2019, 45, 109–118. [Google Scholar] [CrossRef]
- Liu, B.-R.; Jia, G.-M.; Chen, J.; Wang, G. A review of methods for studying microbial diversity in soils. Pedosphere 2006, 16, 18–24. [Google Scholar] [CrossRef]
- Zhou, X.; Ren, L.; Meng, Q.; Li, Y.; Yu, Y.; Yu, J. The next-generation sequencing technology and application. Protein. Cell. 2010, 1, 520–536. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Yang, J.; Yu, Z.; Wilkinson, D.M. The biogeography of abundant and rare bacterioplankton in the lakes and reservoirs of China. ISME J. 2015, 9, 2068–2077. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, C.; Zhang, W.; Di, P.; Yi, N.; Chen, C. Vertical and horizontal assemblage patterns of bacterial communities in a eutrophic river receiving domestic wastewater in southeast China. Environ. Pollut. 2017, 230, 469–478. [Google Scholar] [CrossRef]
- Lu, X.M.; Chen, C.; Zheng, T.L. Metagenomic insights into effects of chemical pollutants on microbial community composition and function in estuarine sediments receiving polluted river water. Microbiol. Ecol. 2017, 73, 791–800. [Google Scholar] [CrossRef]
- Hu, A.; Yang, X.; Chen, N.; Hou, L.; Ma, Y.; Yu, C.P. Response of bacterial communities to environmental changes in a363mesoscale subtropical watershed, Southeast China. Sci. Total Environ. 2014, 472, 746–756. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Wang, P.; Niu, L.; Zhang, W.; Wang, C. Response of bacterial community compositions to different sources of pollutants in sediments of a tributary of Taihu Lake, China. Environ. Sci. Pollut. Res. 2016, 23, 13886–13894. [Google Scholar] [CrossRef]
- Liu, J.; Chen, X.; Shu, H.Y.; Lin, X.R.; Zhou, Q.X.; Bramryd, T.; Shu WHHuang, L.N. Microbial community structure and function in sediments from e-waste contaminated rivers at Guiyu area of China. Environ. Pollut. 2018, 235, 171–179. [Google Scholar] [CrossRef]
- Cirella, G.T.; Zerbe, S. Sustainable Water Management and Wetland Restoration Strategies in Northern China; Bozen-Bolzano University Press, Free University of Bozen-Bolzano: Italy, France, 2014. [Google Scholar]
- MacDonald, D.D.; Ingersoll, C.G.; Berger, T.A. Development and evaluation of consensus-based sediment quality guidelines for freshwater ecosystems. Arch. Environ. Contam. Toxicol. 2000, 39, 20–31. [Google Scholar] [CrossRef]
- Persaud, D.; Jaagumagi, R.; Hayton, A. Guidelines for the Protection and Management of Aquatic374sediment Quality in Ontario; Ontario Ministry of the Environment: Toronto, ON, Canada, 1993; Volume 27. [Google Scholar]
- Wei, B.; Yang, L. A review of heavy metal contaminations in urban soils, urban road dusts and agricultural soils from China. Microchem. J. 2010, 94, 99–107. [Google Scholar] [CrossRef]
- Qiu, Y.; Shi, H.C.; He, M. Nitrogen and phosphorous removal in municipal wastewater treatment plants378in China: A review. Int. J. Chem. Eng. 2010, 2010, 914159. [Google Scholar] [CrossRef] [Green Version]
- Borruso, L.; Esposito, A.; Bani, A.; Ciccazzo, S.; Papa, M.; Zerbe, S.; Brusetti, L. Ecological diversity of sediment rhizobacteria associated with Phragmites australis along a drainage canal in the Yellow River watershed. J. Soil. Sed. 2017, 17, 253–265. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, S.; Wang, M.; Sun, W.; Xie, Y.; Peng, H.; Zhong, A.; Liu, H.; Zhang, X.; Yu, H.; et al. Acid mine drainage affects the diversity and metal resistance gene profile of sediment bacterial community along a river. Chemosphere 2019, 217, 790–799. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Kumar, U.; Singh, N.R.; Thatoi, H. Functional diversity and metabolic profile of microbial community of mine soils with different levels of chromium contamination. Int. J. Environ. Health Res. 2020, 30, 461–473. [Google Scholar] [CrossRef]
- Guo, Q.; Majeed, S.; Xu, R.; Zhang, K.; Kakade, A.; Khan, A.; Hafeez, F.Y.; Mao, C.; Liu, P.; Li, X. Heavy metals interact with the microbial community and affect biogas production in anaerobic digestion: A review. J. Environ. Manag. 2019, 240, 266–272. [Google Scholar] [CrossRef]
- Sun, W.; Xiao, E.; Xiao, T.; Krumins, V.; Wang, Q.; Häggblom, M.; Dong, Y.; Tang, S.; Hu, M.; Li, B.; et al. Response of soil microbial communities to elevated antimony and arsenic contamination indicates the relationship between the innate microbiota and contaminant fractions. Environ. Sci. Technol. 2017, 51, 9165–9175. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–3945120. [Google Scholar] [CrossRef] [Green Version]
- Poladyan, A.; Blbulyan, S.; Sahakyan, M.; Lenz, O.; Trchounian, A. Growth of the facultative chemolithoautotrophy Ralstonia eutropha on organic waste materials: Growth characteristics, redox regulation and hydrogenase activity. Microb. Cell. Fact. 2019, 18, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinecke, F.; Steinbüchel, A. Ralstonia eutropha strain H16 as model organism for PHA metabolism and for biotechnological production of technically interesting biopolymers. J. Mol. Microbiol. Biotech. 2009, 16, 91–108. [Google Scholar] [CrossRef]
- Köchling, T.; Sanz, J.L.; Galdino, L.; Florencio, L.; Kato, M.T. Impact of pollution on the microbial diversity of a tropical river in an urbanized region of north eastern Brazil. Int. Microbiol. 2017, 20, 11–24. [Google Scholar]
- Riviere, D.; Desvignes, V.; Pelletier, E.; Chaussonnerie, S.; Guermazi, S.; Weissenbach, J.; Li, T.; Camacho, P.; Sghir, A. Towards the definition of a core of microorganisms involved in anaerobic digestion of sludge. ISME J. 2009, 3, 700–714. [Google Scholar] [CrossRef] [Green Version]
- Hernández, M.; Dumont, M.G.; Yuan, Q.; Conrad, R. Different bacterial populations associated with the roots and rhizosphere of rice incorporate plant-derived carbon. Appl. Environ. Microbiol. 2015, 81, 2244–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.; Zhang, W.; Xie, S.; Zeng, M.; Liu, H.; Yang, J.; Liu, X.; Yang, F. Increasing prevalence of antibiotic resistance genes in manured agricultural soils in northern China. Front. Env. Sci. Eng. 2019, 14, 1. [Google Scholar] [CrossRef]
- Ito, T.; Yoshiguchi, K.; Ariesyady, H.D.; Okabe, S. Identification of a novel acetate-utilizing bacterium belonging to Synergistes group in anaerobic digester sludge. ISME J. 2011, 5, 1844–1856. [Google Scholar] [CrossRef] [Green Version]
- Plugge, C.M.; Balk, M.; Zoetendal, E.G.; Stams, A.J. Gelria glutamica gen. nov., sp. nov., a thermophilic, obligately syntrophic, glutamate-degrading anaerobe. Int. J. Syst. Evol. Microbiol. 2002, 52, 401–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Rinta-Kanto, J.M.; Kettunen, R.; Lens, P.; Collins, G.; Kokko, M.; Rintala, J. Acetotrophic activity facilitates methanogenesis from LCFA at low temperatures: Screening from mesophilic inocula. Archaea 2019, 2019, 1751783. [Google Scholar] [CrossRef]
- Dugan, P.R.; Stoner, D.L.; Pickrum, H.M. The Genus Zoogloea. In The Prokaryotes: Volume 7:416. Proteobacteria: Delta, Epsilon Subclass; Springer: New York, NY, USA, 2006; pp. 960–970. [Google Scholar]
- Whiteley, A.S.; Bailey, M.J. Bacterial community structure and physiological state within an industrial phenol bioremediation system. Appl. Environ. Microbiol. 2000, 66, 2400–2407. [Google Scholar] [CrossRef] [Green Version]
- Bernardet, J.F.; Segers, P.; Vancanneyt, M.; Berthe, F.; Kersters, K.; Vandamme, P. Cutting a Gordian knot: Emended classification and420description of the genus Flavobacterium, emended description of the family Flavobacteriaceae, and proposal of Flavobacterium hydatis nom. Nov. (basonym, Cytophaga aquatilis Strohland Trait 4221978). Int. J. Syst. Evol. Microbiol. 1996, 46, 128–148. [Google Scholar]
- Liu, J.L.; Yao, J.; Duran, R.; Mihucz, V.G.; Hudson-Edwards, K.A. Bacterial shifts during in-situ mineralization bio-treatment to non-ferrous metal (loid) tailings. Environ. Pollut. 2019, 255, 113165. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Meng, D.; Li, J.; Yin, H.; Liu, H.; Liu, X.; Cheng, C.; Xiao, Y.; Liu, Z.; Yan, M. Response of soil microbial communities and microbial interactions to426long-term heavy metal contamination. Environ. Pollut. 2017, 231, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Tipayno, S.C.; Truu, J.; Samaddar, S.; Truu, M.; Preem, J.-K.; Oopkaup, K.; Espenberg, M.; Chatterjee, P.; Kang, Y.; Kim, K.; et al. The bacterial community structure and functional profile in the heavy metal contaminated paddy soils, surrounding a nonferrous smelter in South Korea. Ecol. Evol. 2018, 8, 6157–6168. [Google Scholar] [CrossRef]
- Wittebolle, L.; Marzorati, M.; Clement, L.; Balloi, A.; Daffonchio, D.; Heylen, K.; De Vos, P.; Verstraete, W.; Boon, N. Initial community evenness favours functionality under selective stress. Nature 2009, 458, 623–626. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Land Use | Latitude (GPS) | Longitude (GPS) | Altitude (m) | Time | Date | Description of Environment |
|---|---|---|---|---|---|---|---|
| ZY21 | Agricultural area | N38 58.884 | E100 28.891 | 1454 m | 10:30 | 11 June 2011 | Drainage channel from city to wastewater plant, probable chemical pollution. Small channel. Height of reeds is about 2.5 m. Near the channel is a small pond full of reeds. |
| ZY23 | Industrial area | N38 58.989 | E100 30.001 | 1456 m | 11:30 | 11 June 2011 | Outgoing industrial channel between ZY24 and ZY22. High level of chemical pollution. Small pond surrounded by a dead forest (E, S, W) and a street with a railway (N). Reeds are about 1.5 m high. |
| ZY24 | Industrial area | N38 58.874 | E100 29.999 | 1458 m | 12:00 | 11 June 2011 | Big pond in the channel of high level of only industrial polluted water. Water was black. Dead trees. Nearby, there was big coal piles. Reeds are about 2 m high. |
| ZY25 | Industrial area | N38 58.820 | E100 29.953 | 1456 m | 12:20 | 11 June 2011 | Industrial lake with black water. High level of chemical pollution and bloom algae. Mixed vegetation of reeds and Typha. Near a factory of coal dealing. |
| ZY26 | Industrial area | N38 58.848 | E100 30.206 | 1457 m | 12:40 | 11 June 2011 | Industrial lake with high level of chemical pollution and bloom algae; the height of the reeds is about 3 m. |
| ZY28 | Agricultural area | N39 03.065 | E100 26.927 | 1435 m | 09:20 | 12 June 2011 | Shandan River, as an outgoing channel from an industrial area; Livestock, grassland and agricultural area; height of the reeds about 3 m. |
| ZY29 | Agricultural area | N39 03.065 | E100 26.927 | 1435 m | 09:40 | 12 June 2011 | 1 km far from the entrance of the Shandan River to the Heihe River. Agriculture, corn crops, some livestock; some foam in the water, not so far to a small river dam |
| ZY30 | Agricultural area | N39 03.113 | E100 26.807 | 1432 m | 10:10 | 12 June 2011 | Entrance of the Shandan River into the Heihe River; agricultural and natural area. Wild birds; lot of paw prints from livestock. |
| ZY31 | Agricultural area | N39 03.121 | E100 26.800 | 1432 m | 10:10 | 12 June 2011 | Entrance of the Shandan River into the Heihe River; agricultural and natural area. Wild birds; lot of paw prints from livestock. |
| ZY34 | Natural area | N38 59.258 | E100 27.793 | 1455 m | 12:00 | 12 June 2011 | National natural wetland park. Extensive reed stands; wild birds; not flowing water |
| ZY35 | Natural area | N38 59.314 | E100 27.704 | 1452 m | 12:00 | 12 June 2011 | National natural wetland park. Extensive reed stands; wild birds; not flowing water |
| ZY36 | Urban area | N38 55.992 | E100 28.076 | 1481 m | 09:00 | 13 June 2011 | In Zhangye city, the water source of the urban channel. The reeds are about 3 m high. Organic pollution, although recently a wastewater pipeline was built close to this channel |
| ZY37 | Urban area | N38 56.107 | E100 28.066 | 1483 m | 09:00 | 13 June 2011 | In Zhangye city, the water source of the urban channel. The reeds are about 3 m high. Organic pollution, although recently a wastewater pipeline was built close to this channel |
| ZY38 | Urban area | N38 56.146 | E100 28.085 | 1483 m | 10:20 | 13 June 2011 | In Zhangye city, the water source of the urban channel. The reeds are about 3 m high. Organic pollution, although recently a wastewater pipeline was built close to this channel |
| ZY39 | Urban area | N38 56.280 | E100 28.095 | 1483 m | 11:00 | 13 June 2011 | Presence of antibiotic vials in the water; some handcraft factories; height of reeds is about 2.5 m. |
| ZY40 | Urban area | N38 56.279 | E100 28.092 | 1483 m | 11:20 | 13 June 2011 | Presence of antibiotic vials in the water; some handcraft factories; height of reeds is about 2.5 m. |
| ZY41 | Urban area | N38 56.278 | E100 28.091 | 1483 m | 11:20 | 13 June 2011 | Presence of antibiotic vials in the water; some handcraft factories; height of reeds is about 2.5 m. |
| Site | pH | N % | P (a) | Fe (a) | Al (a) | Mn (b) | Cu (b) | Zn (b) | Cr (b) | Ni (b) | Pb (b) | Co (b) | Hg (b) | Cd (b) | As (b) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ZY21 | 7.6 | 0.16 | 0.88 | 30.3 | 23.4 | 623 | 41 | 102.1 | 56.5 | 47.1 | 23.2 | 14.8 | 0.3 | 0.24 | 12 |
| ZY23 | 8.6 | 0.22 | 1.07 | 28.5 | 27.1 | 570 | 151 | 2023 | 50.2 | 41.8 | 720 | 15.4 | 2.22 | 74.24 | 248 |
| ZY24 | 7.8 | 0.6 | 2.37 | 29.9 | 29.9 | 601 | 111.6 | 313.7 | 81.1 | 49.9 | 201.5 | 14.3 | 0.95 | 6.39 | 184 |
| ZY25 | 8.5 | 0.27 | 1.3 | 30 | 27.1 | 697 | 85.2 | 687.5 | 50.1 | 43.2 | 139.5 | 16.8 | 0.88 | 12.9 | 296 |
| ZY26 | 7.9 | 0.15 | 0.82 | 28.7 | 25.1 | 624 | 50 | 163.1 | 85.8 | 52.3 | 52.6 | 12.8 | 0.09 | 1.22 | 80.5 |
| ZY28 | 7.9 | 0.11 | 0.64 | 22.5 | 14.4 | 476 | 23.4 | 133 | 93.4 | 52.6 | 34.1 | 9.24 | 0.19 | 2.07 | 14 |
| ZY29 | 8.5 | 0.02 | 0.57 | 19 | 13 | 508 | 12 | 34.5 | 20.8 | 14.2 | 12.2 | 7.89 | 0.01 | 0.26 | 10 |
| ZY30 | 8.2 | 0.02 | 0.55 | 20.5 | 14.2 | 536 | 13.1 | 40.5 | 26.3 | 19.8 | 13.2 | 8.5 | 0.01 | 0.14 | 8.6 |
| ZY31 | 8.2 | 0.05 | 0.67 | 26.5 | 15.5 | 565 | 14 | 41 | 30.9 | 17 | 15 | 9.3 | 0.02 | 0.23 | 12.1 |
| ZY34 | 8.2 | 0.1 | 0.57 | 27.1 | 22.3 | 663 | 29 | 56.39 | 45.2 | 35.8 | 15.3 | 13.4 | 0.02 | 0.2 | 16.9 |
| ZY35 | 8.1 | 0.12 | 0.57 | 30.4 | 25.1 | 723 | 31 | 62.6 | 44.1 | 38.9 | 16.1 | 14.6 | 0.02 | 0.21 | 15.4 |
| ZY36 | 7.7 | 0.99 | 2.47 | 32.5 | 28.4 | 462 | 64.1 | 184 | 116 | 54.9 | 28.4 | 13.6 | 0.15 | 0.29 | 9.2 |
| ZY37 | 8.1 | 0.49 | 1.66 | 29.2 | 21.5 | 527 | 50.6 | 172 | 71.1 | 46 | 29.3 | 13.3 | 2.09 | 0.31 | 11.6 |
| ZY38 | 7.8 | 0.23 | 0.8 | 35.6 | 30.4 | 768 | 36.9 | 90.1 | 65.4 | 54.1 | 22.5 | 16.8 | 0.32 | 0.12 | 12.8 |
| ZY39 | 8.3 | 0.74 | 2.21 | 36.3 | 32.8 | 614 | 71.8 | 264.2 | 95.1 | 52 | 40.4 | 15.6 | 5.95 | 0.49 | 18 |
| ZY40 | 7.9 | 0.41 | 1.48 | 25.8 | 21 | 426 | 37.1 | 127.9 | 70 | 42.7 | 23.8 | 12.3 | 5.51 | 0.26 | 8.3 |
| ZY41 | 8.6 | 0.25 | 1.01 | 26.6 | 15.6 | 506 | 31.4 | 82.6 | 57.4 | 42.7 | 18.4 | 13.3 | 1.42 | 0.13 | 6.5 |
| Richness | Shannon | Evenness | |
|---|---|---|---|
| ZY21 | 369 | 228.53 | 0.637 |
| ZY23 | 599 | 421.14 | 0.655 |
| ZY24 | 441 | 197.01 | 0.601 |
| ZY25 | 643 | 489.04 | 0.664 |
| ZY26 | 496 | 208.30 | 0.596 |
| ZY28 | 528 | 289.21 | 0.627 |
| ZY29 | 544 | 340.29 | 0.642 |
| ZY30 | 519 | 331.42 | 0.643 |
| ZY31 | 527 | 296.64 | 0.630 |
| ZY34 | 564 | 408.14 | 0.658 |
| ZY35 | 591 | 399.30 | 0.651 |
| ZY36 | 459 | 239.70 | 0.620 |
| ZY37 | 396 | 133.16 | 0.567 |
| ZY38 | 457 | 245.96 | 0.623 |
| ZY39 | 349 | 90.77 | 0.534 |
| ZY40 | 412 | 184.03 | 0.600 |
| ZY41 | 459 | 209.66 | 0.605 |
| Environmental Variables | Diversity Indices | |||||
|---|---|---|---|---|---|---|
| Richness | p Value | Shannon | p Value | Evenness | p Value | |
| N | −0.47 | n.s. | −0.55 | 0.021 | −0.54 | 0.022 |
| P | −0.52 | 0.031 | −0.60 | 0.009 | −0.60 | 0.009 |
| pH | 0.39 | n.s. | 0.29 | n.s. | 0.22 | n.s. |
| Fe | −0.34 | n.s. | −0.38 | n.s. | −0.36 | n.s. |
| Al | −0.2 | n.s. | −0.24 | n.s. | −0.24 | n.s. |
| Mn | 0.26 | n.s. | 0.25 | n.s. | 0.23 | n.s. |
| Cu | −0.10 | n.s. | −0.20 | n.s. | −0.22 | n.s. |
| Zn | 0.10 | n.s. | −0.02 | n.s. | −0.08 | n.s. |
| Cr | −0.48 | 0.048 | −0.56 | 0.018 | −0.56 | 0.019 |
| Ni | −0.35 | n.s. | −0.38 | n.s. | −0.36 | n.s. |
| Pb | 0.2 | n.s. | 0.09 | n.s. | 0.03 | n.s. |
| Co | −0.14 | n.s. | −0.15 | n.s. | −0.14 | n.s. |
| Hg | −0.48 | 0.049 | −0.57 | 0.015 | −0.57 | 0.016 |
| Cd | 0.41 | n.s. | 0.27 | n.s. | 0.18 | n.s. |
| As | 0.42 | n.s. | 0.28 | n.s. | 0.19 | n.s. |
| Permanova | |||||
|---|---|---|---|---|---|
| Df | Sum Of Sqs | R2 | F | Pr (>F) | |
| Land_use | 3 | 1.8013 | 0.2981 | 1.8404 | 0.001 |
| Residual | 13 | 4.2413 | 0.7019 | ||
| Total | 16 | 6.0426 | 1.0000 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piergiacomo, F.; Borruso, L.; Esposito, A.; Zerbe, S.; Brusetti, L. The Geochemical Drivers of Bacterial Community Diversity in the Watershed Sediments of the Heihe River (Northern China). Water 2022, 14, 1948. https://doi.org/10.3390/w14121948
Piergiacomo F, Borruso L, Esposito A, Zerbe S, Brusetti L. The Geochemical Drivers of Bacterial Community Diversity in the Watershed Sediments of the Heihe River (Northern China). Water. 2022; 14(12):1948. https://doi.org/10.3390/w14121948
Chicago/Turabian StylePiergiacomo, Federica, Luigimaria Borruso, Alfonso Esposito, Stefan Zerbe, and Lorenzo Brusetti. 2022. "The Geochemical Drivers of Bacterial Community Diversity in the Watershed Sediments of the Heihe River (Northern China)" Water 14, no. 12: 1948. https://doi.org/10.3390/w14121948
APA StylePiergiacomo, F., Borruso, L., Esposito, A., Zerbe, S., & Brusetti, L. (2022). The Geochemical Drivers of Bacterial Community Diversity in the Watershed Sediments of the Heihe River (Northern China). Water, 14(12), 1948. https://doi.org/10.3390/w14121948