
Mercury Accumulation in a Stream Ecosystem: Linking Labile Mercury in Sediment Porewaters to Bioaccumulative Mercury in Trophic Webs



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
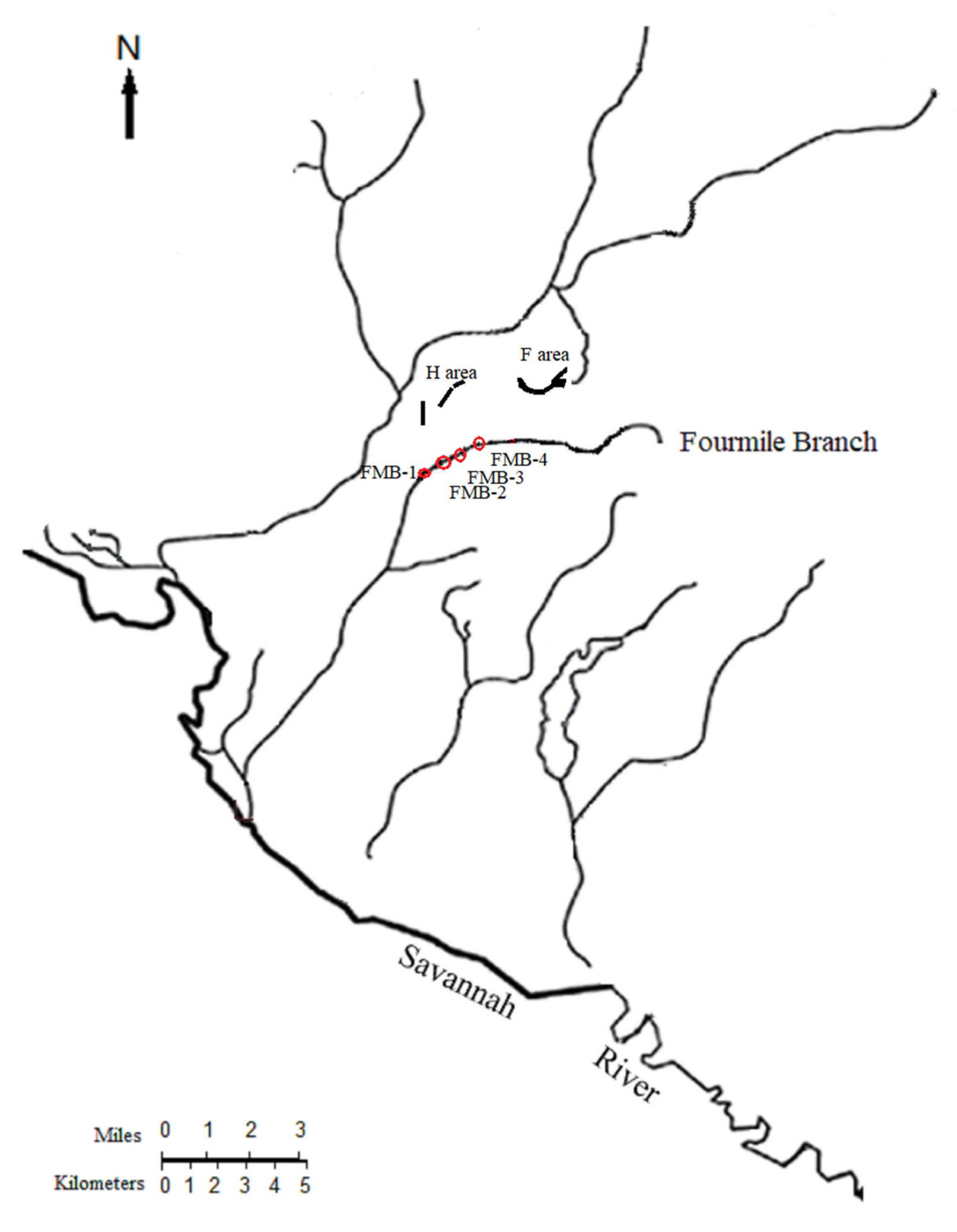
2.1. Sampling Site
2.2. Sampling Strategy
2.3. Sediment Porewater
2.4. Bulk Sediment
2.5. Biota
2.6. Chemical Analysis
2.7. Statistics
3. Results
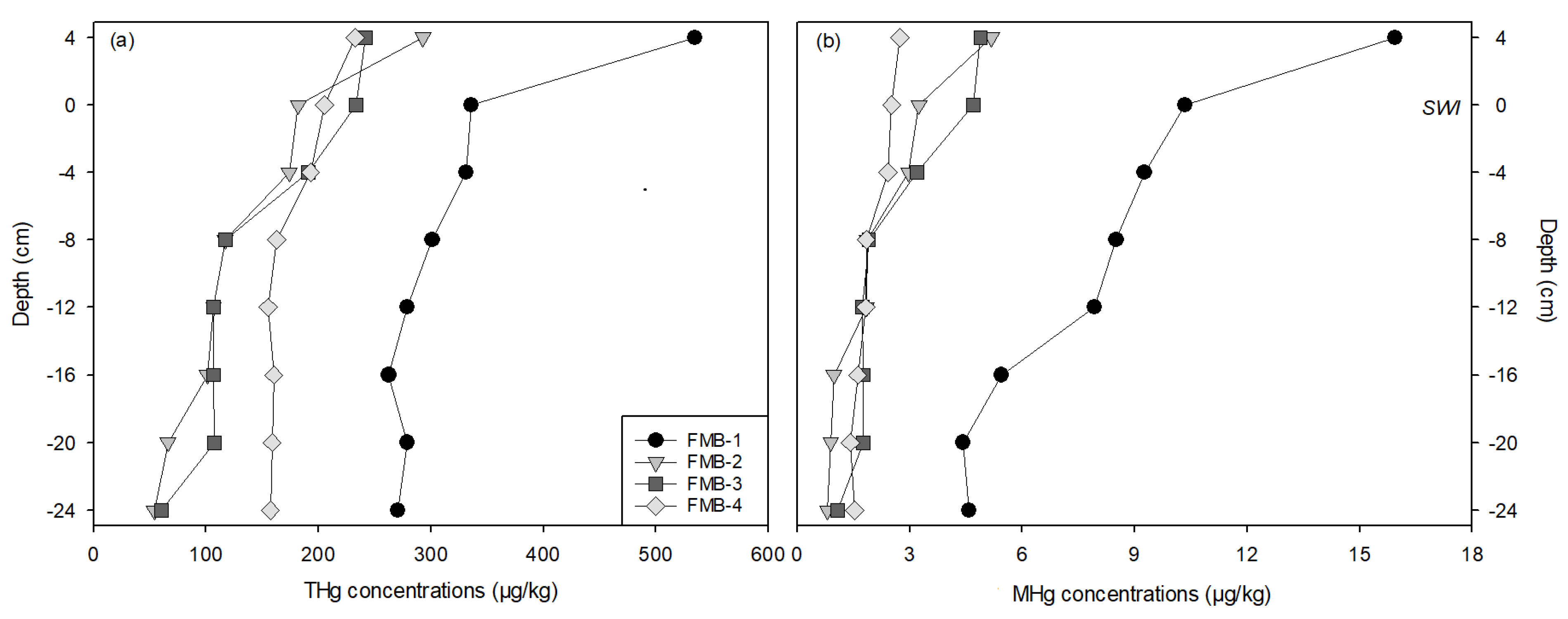
3.1. Mercury in Bulk Sediment
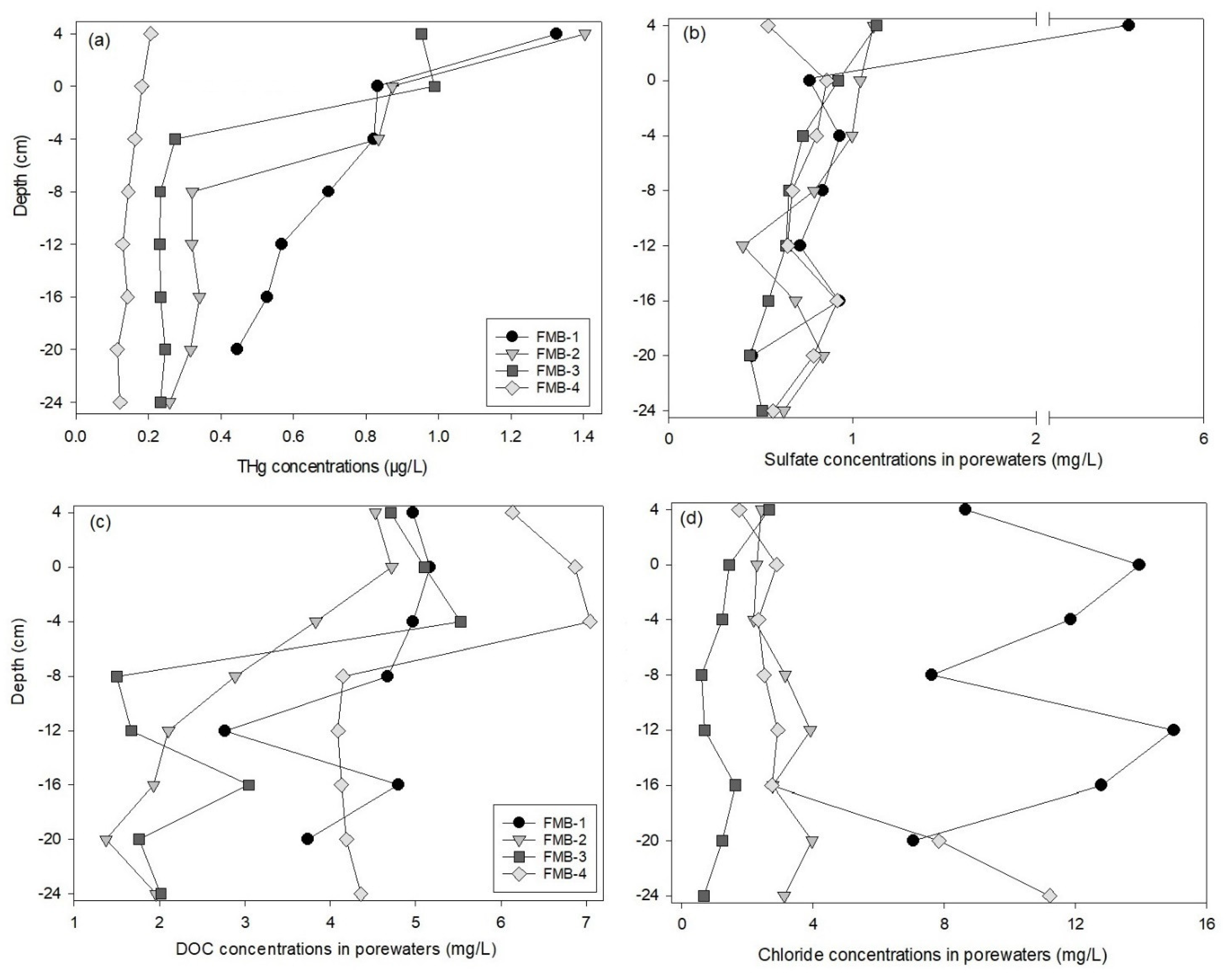
3.2. Mercury in Sediment Porewaters
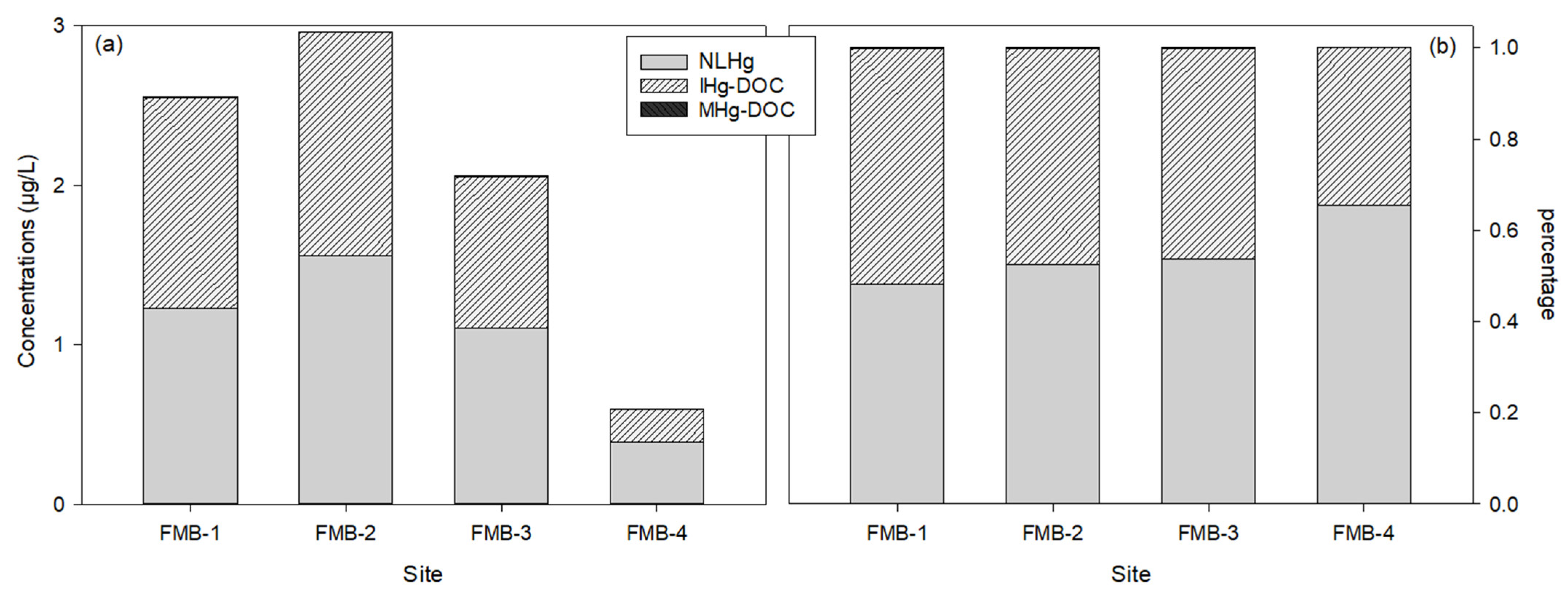
3.3. Mercury Speciation
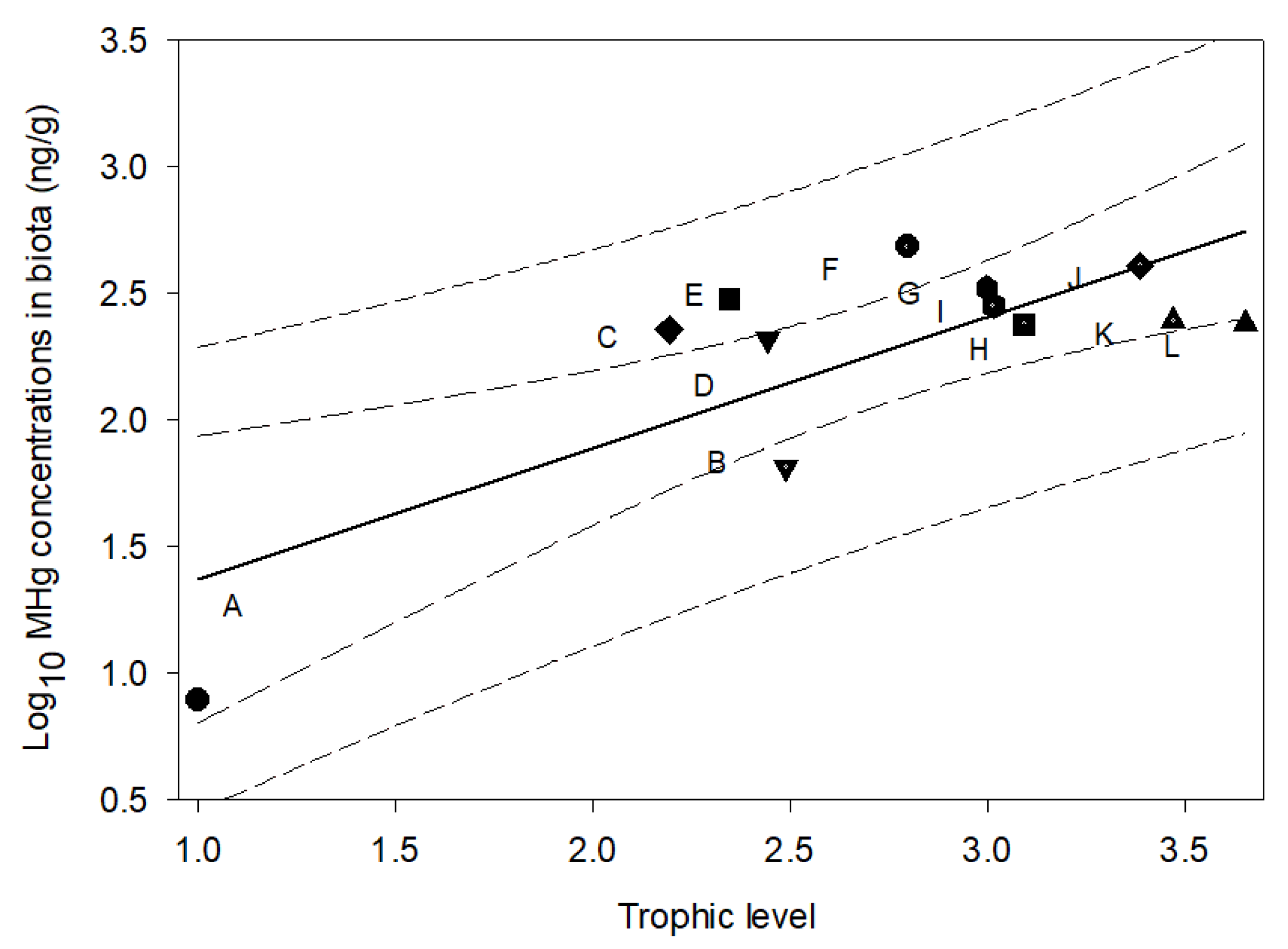
3.4. Mercury in Biota
4. Discussion
4.1. Mercury Deposition in the Sediment
4.2. Mercury Methylation and Methylmercury Production
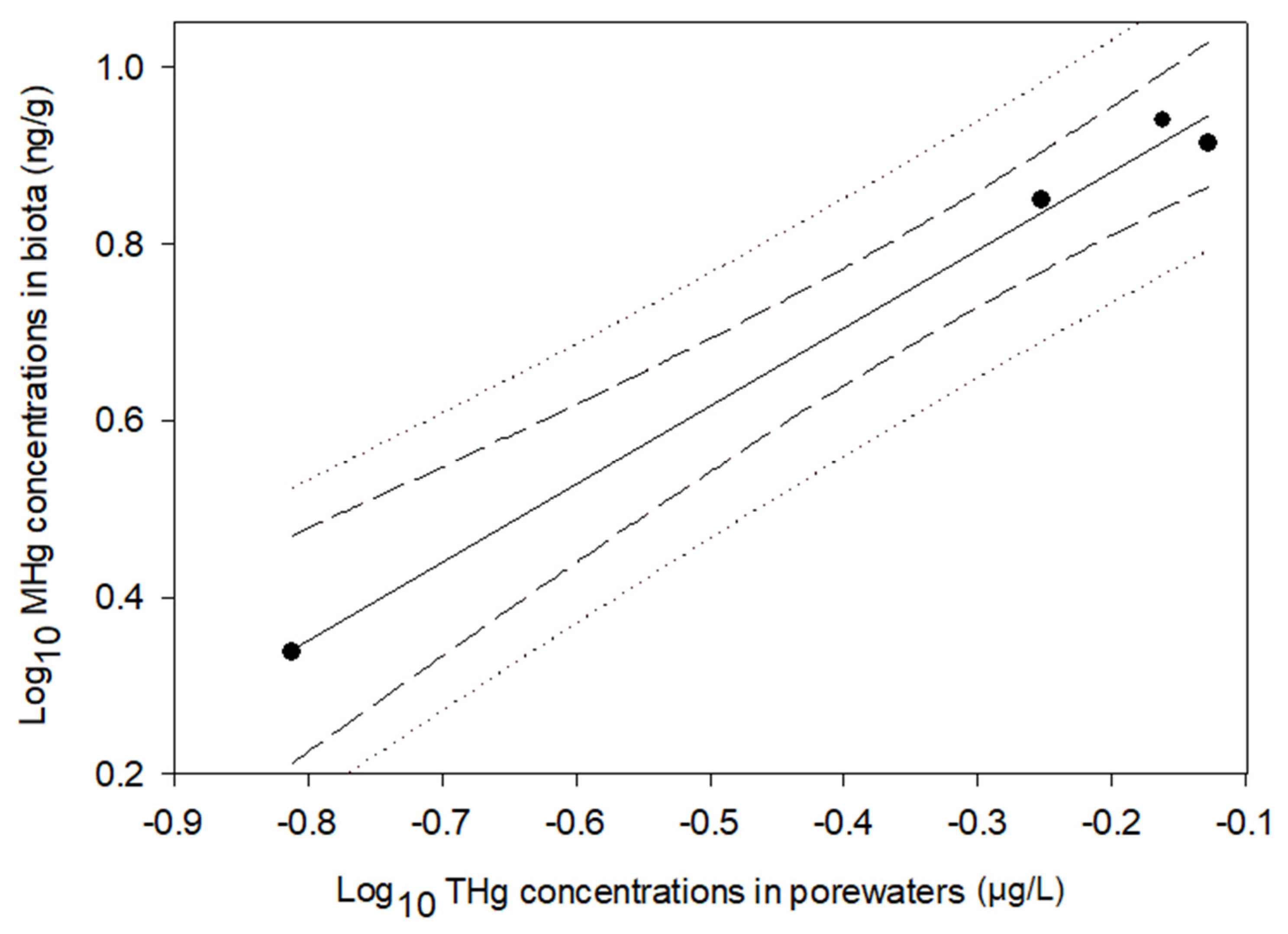
4.3. Mercury in Biotic and Abiotic Environments
4.4. Application of Peepers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, X.; Zhang, Q.; Wang, W.-X. Linking mercury, carbon, and nitrogen stable isotopes in Tibetan biota: Implications for using mercury stable isotopes as source tracers. Sci. Rep. 2016, 6, 25394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branco, V.; Caito, S.; Farina, M.; daRocha, J.T.; Aschner, M.; Carvalho, C. Biomarkers of mercury toxicity: Past, present, and future trends. J. Toxicol. Environ. Health Part B Crit. Rev. 2017, 20, 119–154. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.C.; Xu, X.; Condon, A.; Liang, L. Floodplain methylmercury biomagnification factor higher than that of the contiguous river (South River, Virginia USA). Environ. Pollut. 2011, 159, 2840–2844. [Google Scholar] [CrossRef] [PubMed]
- Jackson, T.A. Long-range atmospheric transport of mercury to ecosystems, and the importance of anthropogenic emissions—A critical review and evaluation of the published evidence. Environ. Rev. 1997, 5, 99–120. [Google Scholar] [CrossRef]
- Hudelson, K.E.; Drevnick, P.E.; Wang, F.; Armstrong, D.; Fisk, A.T. Mercury methylation and demethylation potentials in Arctic lake sediments. Chemosphere 2020, 248, 126001. [Google Scholar] [CrossRef]
- Liang, P.; Wu, S.; Zhang, C.; Xu, J.; Christie, P.; Zhang, J.; Cao, Y. The role of antibiotics in mercury methylation in marine sediments. J. Hazard Mater. 2018, 360, 1–5. [Google Scholar] [CrossRef]
- Ma, M.; Du, H.; Wang, D.; Kang, S.; Sun, T. Biotically mediated mercury methylation in the soils and sediments of Nam Co Lake, Tibetan Plateau. Environ. Pollut. 2017, 227, 243–251. [Google Scholar] [CrossRef]
- Compeau, G.; Bartha, R. Sulfate-reducing bacteria: Principal methylators of mercury in anoxic estuarine sediment. Appl. Environ. Microbiol. 1985, 50, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Gerbig, C.; Kim, C.; Stegemeier, J.; Ryan, J.N.; Aiken, G.R. Formation of nanocolloidal metacinnabar in mercury-DOM-sulfide systems. Environ. Sci. Technol. 2011, 45, 9180–9187. [Google Scholar] [CrossRef]
- Deonarine, A.; Hsu-Kim, H. Precipitation of mercuric sulfide nanoparticles in NOM-containing water: Implications for the natural environment. Environ. Sci. Technol. 2009, 43, 2368–2373. [Google Scholar] [CrossRef]
- Wang, M.; Li, Y.; Zhao, D.; Zhuang, L.; Yang, G.; Gong, Y. Immobilization of mercury by iron sulfide nanoparticles alters mercury speciation and microbial methylation in contaminated groundwater. Chem. Eng. J. 2020, 381, 122664. [Google Scholar] [CrossRef]
- Gionfriddo, C.M.; Stott, M.B.; Power, J.F.; Ogorek, J.M.; Krabbenhoft, D.P.; Wick, R.; Holt, K.; Chen, L.; Thomas, B.C.; Banfield, J.F.; et al. Genome-resolved metagenomics and detailed geochemical speciation analyses yield new insights into microbial mercury cycling in geothermal springs. Appl. Environ. Microbiol. 2020, 86, e00176-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Whitman, W.B. Metabolic, phylogenetic, and ecological diversity of the methanogenic archaea. Ann. N. Y. Acad. Sci. 2008, 1125, 171–189. [Google Scholar] [CrossRef] [PubMed]
- Fleming, E.J.; Mack, E.E.; Green, P.G.; Nelson, D.C. Mercury methylation from unexpected sources: Molybdateinhibited freshwater sediments and an iron-reducing bacterium. Appl. Environ. Microbiol. 2006, 72, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Kerin, E.J.; Gilmour, C.C.; Roden, E.; Suzuki, M.T.; Coates, J.D.; Mason, R.P. Mercury methylation by dissimilatory ironreducing bacteria. Appl. Environ. Microbiol. 2006, 72, 7919–7921. [Google Scholar] [CrossRef] [Green Version]
- Gilmour, C.C.; Podar, M.; Bullock, A.L.; Graham, A.M.; Brown, S.D.; Somenahally, A.C.; Johs, A.; Hurt, J.R.A.; Bailey, K.L.; Elias, D.A. Mercury methylation by novel microorganisms from new environments. Environ. Sci. Technol. 2013, 47, 11810–11820. [Google Scholar] [CrossRef]
- Yu, R.-O.; Reinfelder, J.R.; Hines, M.E.; Barkay, T. Mercury methylation by the methanogen Methanospirillum hungatei. Appl. Environ. Microbiol. 2013, 79, 6325–6330. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Jiang, T.; Wang, F.; Zhang, J.; Wang, D.; Huang, R.; Yin, D.; Liu, Z.; Wang, J. Inorganic sulfur and mercury speciation in the water level fluctuation zone of the Three Gorges Reservoir, China: The role of inorganic reduced sulfur on mercury methylation. Environ. Pollut. 2018, 237, 1112–1123. [Google Scholar] [CrossRef]
- Mason, R.P.; Sheu, G.R. The role of the ocean in the global mercury cycle. Glob. Biogeochem. Cycles 2002, 16, 1093. [Google Scholar] [CrossRef]
- Schuster, P.F.; Shanley, J.B.; Marvin-Dipasquale, M.; Reddy, M.M.; Aiken, G.R.; Roth, D.A.; Taylor, H.E.; Krabbenhoft, D.P.; DeWild, J.F. Mercury and organic carbon dynamics during runoff episodes from a northeastern USA watershed. Water Air Soil 2008, 187, 89–108. [Google Scholar] [CrossRef] [Green Version]
- Benoit, J.M.; Gilmour, C.C.; Mason, R.P.; Heyes, A. Sulfide controls on mercury speciation and bioavailability to methylating bacteria in sediment pore waters. Environ. Sci. Technol. 1999, 33, 951–957. [Google Scholar] [CrossRef]
- Hsu-Kim, H.; Eckley, C.S.; Acha, D.; Feng, X.; Gilmour, C.C.; Jonsson, S.; Mitchell, C.P.J. Challenges and opportunities for managing aquatic mercury pollution in altered landscapes. Ambio 2018, 47, 141–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Bryan, A.L.; Mills, G.L.; Korotasz, A.M. Mercury speciation, bioavailability, and biomagnification in contaminated streams on the Savannah River Site (SC, USA). Sci. Total Environ. 2019, 668, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Obrist, D.; Kirk, J.L.; Zhang, L.; Sunderland, E.M.; Jiskra, M.; Selin, N.E. A review of global environmentalmercury processes in response to human and natural perturbations: Changes of emissions, climate, and land use. Ambio 2018, 47, 116–140. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.-L.; Liu, Y.-R.; Guan, W.-Y.; Zhang, H.; Qu, X.-M.; Zhang, T. Understanding mercury methylation in the changing environment: Recent advances in assessing microbial methylators and mercury bioavailability. Sci. Total Environ. 2020, 714, 136827. [Google Scholar] [CrossRef]
- Rivera, N.A.; Bippus, P.M.; Hsu-Kim, H. Relative reactivity and bioavailability of mercury sorbed to or coprecipitated with aged iron sulfides. Environ. Sci. Technol. 2019, 53, 7391–7399. [Google Scholar] [CrossRef]
- Jiskra, M.; Saile, D.; Wiederhold, J.G.; Bourdon, B.; Bjorn, E.; Kretzschmar, R. Kinetics of Hg(II) exchange between organic ligands, goethite, and natural organic matter studied with an enriched stable isotope approach. Environ. Sci. Technol. 2014, 48, 13207–13217. [Google Scholar] [CrossRef] [Green Version]
- Stenzler, B.; Hinz, A.; Ruuskanen, M.; Poulain, A.J. Ionic strength differentially affects the bioavailability of neutral and negatively charged inorganic Hg complexes. Environ. Sci. Technol. 2017, 51, 9653–9662. [Google Scholar] [CrossRef]
- Ndu, U.; Christensen, G.A.; Rivera, N.A.; Gionfriddo, C.M.; Deshusses, M.A.; Elias, D.A.; Hsu-Kim, H. Quantification of mercury bioavailability for methylation using diffusive gradient in thin-film samplers. Environ. Sci. Technol. 2018, 52, 8521–8529. [Google Scholar] [CrossRef]
- Hesslein, R.H. An in situ sampler for close interval pore water studies. Limnol. Oceanogr. 1976, 21, 912–914. [Google Scholar] [CrossRef]
- Davison, W.; Grime, G.W.; Morgan, J.A.W.; Clarke, K. Distribution of dissolved iron in sediment pore waters at submillimeter resolution. Nature 1991, 352, 323–325. [Google Scholar] [CrossRef]
- Clarisse, O.; Foucher, D.; Hintelmann, H. Methylmercury speciation in the dissolved phase of a stratified lake using the diffusive gradient in thin film technique. Environ. Pollut. 2009, 157, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Clarisse, O.; Dimock, B.; Hintelmann, H.; Best, E.P. Predicting net mercury methylation in sediments using diffusive gradient in thin films measurements. Environ. Sci. Technol. 2011, 45, 1506–1512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Davison, W. Performance characteristics of diffusion gradients in thin films for the in situ measurement of trace metals in aqueous solution. Anal. Chem. 1995, 67, 3391–3400. [Google Scholar] [CrossRef]
- Philipps, R.R.; Xu, X.Y.; Bringolf, R.B.; Mills, G.L. Evaluation of the DGT technique for predicting uptake of metal mixtures by fathead minnow (Pimephales promelas) and yellow lampmussel (Lampsilis cariosa). Environ. Toxicol. Chem. 2019, 38, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Philipps, R.R.; Xu, X.Y.; Mills, G.L.; Bringolf, R.B. Evaluation of diffusive gradients in thin films for prediction of copper bioaccumulation by yellow lampmussel (Lampsilis cariosa) and fathead minnow (Pimephales promelas). Environ. Toxicol. Chem. 2018, 37, 1535–1544. [Google Scholar] [CrossRef]
- Serbst, J.R.; Burgess, R.M.; Kuhn, A.; Edwards, P.A.; Cantwell, M.G.; Pelletier, M.C.; Berry, W.J. Precision of dialysis (peeper) sampling of cadmium in marine sediment interstitial water. Arch. Environ. Contam. Toxicol. 2003, 45, 297–305. [Google Scholar] [CrossRef]
- Liu, J.L.; Feng, X.B.; Qiu, G.L.; Yao, H.; Shang, L.H.; Yan, H.Y. Intercomparison and Applicability of Some Dynamic and Equilibrium Approaches to Determine Methylated Mercury Species in Pore Water. Environ. Toxicol. Chem. 2011, 30, 1739–1744. [Google Scholar] [CrossRef]
- Mason, R.; Bloom, N.; Cappellino, S.; Gill, G.; Benoit, J.; Dobbs, C. Investigation of porewater sampling methods for mercury and methylmercury. Environ. Sci. Technol. 1998, 32, 4031–4040. [Google Scholar] [CrossRef]
- Montgomery, S.; Mucci, A.; Lucotte, M. The application of in situ dialysis samplers for close interval investigations of total dissolved mercury in interstitial waters. Water Air Soil Poll. 1996, 87, 219–229. [Google Scholar] [CrossRef]
- Wang, J.; Newman, M.C.; Xiaoyu, X.; Condon, A.; Liang, L. Floodplain methylmercury biomagnification factor higher and more variable than that of the contiguous South River (Virginia USA). Ecotoxicol. Environ. Saf. 2013, 92, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.P.; Reinfelder, J.R.; Morel, F.M.M. Uptake, toxicity and trophic transfer of mercury in a coastal diatom. Environ. Sci. Technol. 1996, 30, 1835–1845. [Google Scholar] [CrossRef]
- Xu, X.; Newman, M.C.; Fabrizio, M.C.; Liang, L. An ecologically framed mercury survey of finfish of the lower Chesapeake Bay. Arch. Environ. Contam. Toxicol. 2013, 65, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Watras, C.J.; Bloom, N.S. Mercury and Methylmercury in Individual Zooplankton—Implications for Bioaccumulation. Limnol. Oceanogr. 1992, 37, 1313–1318. [Google Scholar] [CrossRef]
- Bishop, C.A.; Koster, M.D.; Chek, A.A.; Hussell, D.J.T.; Jock, K. Chlorinated Hydrocarbons and Mercury in Sediments, Red-Winged Blackbirds (Agelaius-Phoeniceus) and Tree Swallows (Tachycineta Bicolor) from Wetlands in the Great-Lakes St-Lawrence-River Basin. Environ. Toxicol. Chem. 1995, 14, 491–501. [Google Scholar] [CrossRef]
- Horton, J.H. Mercury in the Separations Areas Seepage Basins; DPST-74-231; Du Pont: Aiken, SC, USA, 1997. [Google Scholar]
- Kuhne, W.W.; Halverson, N.V.; Jackson, D.G.; Jannik, G.T.; Looney, B.B.; Paller, M.H. 2015 Assessment of Mercury in the savannah River Site Environment and Responses to the Agency for Toxic Substances and Disease Registry 2012: Report on Assessment of Biota Exposure to Mercury Originating from the Savannah River Site; Savannah River Nuclear Solutions: Aiken, SC, USA, 2015. [Google Scholar]
- Smith, L.; Jagoe, C.; Carl, F. Chlor-alkali plant contributes to mercury contamination in the Savannah River. In Proceedings of the 2007 Georgia Water Resources Conference at the University of Georgia, Athens, GA, USA, 27–29 March 2007. [Google Scholar]
- Newman, M.C. Comprehensive cooling water report—Volume 2: Water quality. Natl. Tech. Inf. Serv. 1986, 2, 13–19. [Google Scholar]
- Haskins, D.; Brown, M.K.; Qin, C.; Xu, X.; Pilgrim, M.; Tuberville, T.D. Multi-decadal trends in mercury and methylmercury concentrations in the brown watersnake (Nerodia taxispilota). Environ. Pollut. 2021, 276, 116722. [Google Scholar] [CrossRef]
- Killian, T.H.; Colb, N.L.; Corbo, P.; Marine, I.W. Environmental Information Document F-Area Seepage Basin; Savannah River Lab: Aiken, SC, USA, 1987. [Google Scholar]
- Killian, T.H.; Colb, N.L.; Corbo, P.; Marine, I.W. Environmental Information Document H-Area Seepage Basin; Savannah River Lab: Aiken, SC, USA, 1987. [Google Scholar]
- Lanier, T.H. Determination of the 100-Year Flood Plain on Fourmile Branch at the Savannah River Site, South Carolina, 1996; Water-Resources Investigations Report 96-4271; U.S. Geological Survey: Columbia, SC, USA, 1997. [Google Scholar]
- Pesch, C.E.; Hansen, D.J.; Boothman, W.S.; Berry, W.J.; Mahony, J.D. The Role of Acid Volatile Sulfide and Interstitial Water Metal Concentrations in Determining Bioavailability of Cadmium and Nickel from Contaminated Sediments to the Marine Polychaete Neanthes Arenaceodentata. Environ. Toxicol. Chem. 1995, 14, 129–141. [Google Scholar] [CrossRef]
- Bufflap, S.E.; Allen, H.E. Comparison of pore-water sampling techniques for trace-metals. Water Res. 1995, 29, 2051–2054. [Google Scholar] [CrossRef]
- Brown, K.; Caldwell, D. Sampling Pore Water Sediments; Brown and Caldwell: Andover, MA, USA; Upper Saddle River, NJ, USA, 2016. [Google Scholar]
- Qin, C.; Xu, X.; Peck, E. Sink or source? Insights into the behavior of copper and zinc in the sediment porewater of a constructed wetland by peepers. Sci. Total Environ. 2022, 821, 153127. [Google Scholar] [CrossRef]
- USEPA. Method 1631, Revision E: Mercury in Water by Oxidation, Purge and Trap, and Cold Vapor Atomic Fluorescence Spectrometry; United States Environmental Protection Agency, Office of Water: Washington, DC, USA, 2002. [Google Scholar]
- Liang, L.; Horvat, M.; Feng, X.; Shang, L.; Li, H.; Pang, P. Re-evaluation of distillation and comparison with HNO3 leaching/solvent extraction for isolation of methylmercury compounds from sediment/soil samples. Appl. Organometal. Chem. 2004, 18, 264–270. [Google Scholar] [CrossRef]
- Liang, L.; Bloom, N.S.; Horvat, M. Simultaneous determination of mercury speciation in biological materials by GC/CVAFS after ethylation and room temperature precollection. Clin. Chem. 1994, 40, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Horvat, M.; Bloom, N.S. An improved speciation method for mercury by GC/CVAFS after aqueous phase ethylation and room temperature precollection. Talanta 1994, 41, 371–379. [Google Scholar] [CrossRef]
- Xu, X.; Mills, G.L. Do constructed wetlands remove metals or increase metal bioavailability? J. Environ. Manag. 2018, 218, 245–255. [Google Scholar] [CrossRef]
- Baird, D.; Ulanowicz, R.E. The seasonal dynamics of the Chesapeake Bay ecosystem. Ecol. Monogr. 1989, 59, 329–364. [Google Scholar] [CrossRef]
- Minagawa, M.; Wada, E. Stepwise enrichment of 15 N along food chains: Further evidence and the relation between 15 N and animal age. Geochim. Cosmochim. Acta 1984, 48, 1135–1140. [Google Scholar] [CrossRef]
- USEPA. EPA Releases Report on Fish Contamination in US Lakes and Reservoirs; United States Environmental Protection Agency, Office of Water: Washington, DC, USA, 2009. [Google Scholar]
- Shi, J.B.; Ip, C.C.M.; Zhang, G.; Jiang, G.B.; Li, X.D. Mercury profiles in sediments of the Pearl River Estuary and the surrounding coastal area of South China. Environ. Pollut. 2010, 158, 1974–1979. [Google Scholar] [CrossRef] [Green Version]
- Wasserman, J.C.; Freitas-Pinto, A.A.P.; Amouroux, D. Mercury concentrations in sediment profiles of a degraded tropical coastal environment. Environ. Technol. 2000, 21, 297–305. [Google Scholar] [CrossRef]
- Lambertsson, L.; Nilsson, M. Organic material: The primary control on mercury methylation and ambient methyl mercury concentrations in estuarine sediments. Environ. Sci. Technol. 2006, 40, 1822–1829. [Google Scholar] [CrossRef]
- Vairavamurthy, M.A.; Maletic, D.; Wang, S.K.; Manowitz, B.; Eglinton, T.; Lyons, T. Characterization of sulfur-containing functional groups in sedimentary humic substances by X-ray absorption near-edge structure spectroscopy. Energy Fuels 1997, 11, 546–553. [Google Scholar] [CrossRef]
- Skyllberg, U. Chemical speciation of mercury in soil and sediment. In Environmental Chemistry and Toxicology of Hg; Liu, G., Cai, Y., O’Driscoll, N., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012. [Google Scholar]
- Karlsson, T.; Skyllberg, U. Bonding of ppb levels of methyl mercury to reduced sulfur groups in soil organic matter. Environ. Sci. Technol. 2003, 37, 4912–4918. [Google Scholar] [CrossRef] [PubMed]
- Black, F.J.; Bruland, K.W.; Flegal, A.R. Competing ligand exchange-solid phase extraction method for the determination of the complexation of dissolved inorganic mercury(II) in natural waters. Anal. Chim. Acta 2007, 598, 318–333. [Google Scholar] [CrossRef] [PubMed]
- Skyllberg, U.; Bloom, P.R.; Qian, J.; Lin, C.M.; Bleam, W.F. Complexation of mercury(II) in soil organic matter: EXAFS evidence for linear two-coordination with reduced sulfur groups. Environ. Sci. Technol. 2006, 40, 4174–4180. [Google Scholar] [CrossRef] [PubMed]
- He, T.R.; Lu, J.; Yang, F.; Feng, X.B. Horizontal and vertical variability of mercury species in pore water and sediments in small lakes in Ontario. Sci. Total Environ. 2007, 386, 53–64. [Google Scholar] [CrossRef]
- Matsuyama, A.; Yano, S.; Taninaka, T.; Kindaichi, M.; Sonoda, I.; Tada, A.; Akagi, H. Chemical characteristics of dissolved mercury in the pore water of Minamata Bay sediments. Mar. Pollut. Bull. 2018, 129, 503–511. [Google Scholar] [CrossRef]
- Pisanello, F.; Marziali, L.; Rosignoli, F.; Poma, G.; Roscioli, C.; Pozzoni, F.; Guzzella, L. In situ bioavailability of DDT and Hg in sediments of the Toce River (Lake Maggiore basin, Northern Italy): Accumulation in benthic invertebrates and passive samplers. Environ. Sci. Pollut. Res. 2016, 23, 10542–10555. [Google Scholar] [CrossRef]
- Washburn, S.J.; Blum, J.D.; Kurz, A.Y.; Pizzuto, J.E. Spatial and temporal variation in the isotopic composition of mercury in the South River, VA. Chem. Geol. 2018, 494, 96–108. [Google Scholar] [CrossRef]
- Kneer, M.L.; White, A.; Rolfhus, K.R.; Jeremiason, J.D.; Johnson, N.W.; Ginder-Vogel, M. Impact of Dissolved Organic Matter on Porewater Hg and MeHg Concentrations in St. Louis River Estuary Sediments. ACS Earth Space Chem. 2020, 4, 1386–1397. [Google Scholar] [CrossRef]
- Liu, G.; Li, Y.; Cai, Y. Adsorption of mercury on soils in the aquatic environment. In Environmental Chemistry and Toxicology of Hg; Liu, G., Cai, Y., O’Driscoll, N., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012. [Google Scholar]
- Zhong, H.; Wang, W.X. Inorganic Mercury Binding with Different Sulfur Species in Anoxic Sediments and Their Gut Juice Extractions. Environ. Toxicol. Chem. 2009, 28, 1851–1857. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, W.F.; Lamborg, C.H.; Hammerschmidt, C.R. Marine biogeochemical cycling of mercury. Chem. Rev. 2007, 107, 641–662. [Google Scholar] [CrossRef]
- Skyllberg, U.; Qian, J.; Frech, A. Combined XANES and EXAFS study on the bonding of methyl mercury to thiol groups in soil and aquatic organic matter. Phys. Scripta 2005, T115, 894–896. [Google Scholar] [CrossRef]
- Khwaja, A.R.; Bloom, P.R.; Brezonik, P.L. Binding strength of methylmercury to aquatic NOM. Environ. Sci. Technol. 2010, 44, 6151–6156. [Google Scholar] [CrossRef]
- Aiken, G.; Cotsaris, E. Soil and hydrology—Their effect on NOM. J. Am. Water Work. Assoc. 1995, 87, 36–45. [Google Scholar] [CrossRef]
- Pettit, R.E. Organic Matter, Humus, Humate, Humic Acid, Fulvic Acid and Humin: Their Importance in Soil Fertility and Plant Health. Available online: http://www.humates.com/pdf/ORGANICMATTERPettit.pdf (accessed on 1 May 2022).
- Gerbig, C.; Ryan, J.N.; Aiken, G.R. The effects of dissolved organic matter on mercury biogeochemistry. In Environmental Chemistry and Toxicology of Hg; Liu, G., Cai, Y., O’Driscoll, N., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012. [Google Scholar]
- Parks, J.M.; Johs, A.; Podar, M.; Bridou, R.; Hurt, R.A.; Smith, S.D.; Tomanicek, S.J.; Qian, Y.; Brown, S.D.; Brandt, C.C.; et al. The genetic basis for bacterial mercury methylation. Science 2013, 339, 1332–1335. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Riccardi, D.; Beste, A.; Smith, J.C.; Parks, J.M. Mercury Methylation by HgcA: Theory Supports Carbanion Transfer to Hg(II). Inorg. Chem. 2014, 53, 772–777. [Google Scholar] [CrossRef]
- Schaefer, J.K.; Rocks, S.S.; Zheng, W.; Liang, L.; Gu, B.; Morel, F.M.M. Active transport, substrate specificity, and methylation of Hg(II) in anaerobic bacteria. Proc. Natl. Acad. Sci. USA 2011, 108, 8714–8719. [Google Scholar] [CrossRef] [Green Version]
- Adediran, G.A.; Van, L.N.; Song, Y.; Schaefer, J.K.; Skyllberg, U.; Bjorn, E. Microbial biosynthesis of thiol compounds: Implications for speciation, cellular uptake, and methylation of Hg(II). Environ. Sci. Technol. 2019, 53, 8187–8196. [Google Scholar] [CrossRef]
- Barkay, T.; Gillman, M.; Turner, R.R. Effects of dissolved organic carbon and salinity on bioavailability of mercury. Appl. Environ. Microbiol. 1997, 63, 4267–4271. [Google Scholar] [CrossRef] [Green Version]
- Hines, M.E.; Horvat, M.; Faganeli, J.; Bonzongo, J.C.J.; Barkay, T.; Major, E.B.; Scott, K.J.; Bailey, E.A.; Warwick, J.J.; Lyons, W.B. Mercury biogeochemistry in the Idrija River, Slovenia, from above the mine into the Gulf of Trieste. Environ. Res. 2000, 83, 129–139. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, J.; Fu, J.; Shi, J.; Jiang, G. Biomonitoring: An appealing tool for assessment of metal pollution in the aquatic ecosystem. Anal. Chim. Acta 2008, 606, 135–150. [Google Scholar] [CrossRef]
- Xu, X.; Peck, E.; Fletcher, D.E.; Korotasz, A.; Perry, J. Limitations of applying diffusive gradients in thin films to predict bioavailability of metal mixtures in aquatic systems with unstable water chemistries. Environ. Toxicol. Chem. 2020, 39, 2485–2495. [Google Scholar] [CrossRef]
- Teasdale, P.R.; Batley, G.E.; Apte, S.C.; Webster, I.T. Pore-water sampling with sediment peepers. Trac-Trend Anal. Chem. 1995, 14, 250–256. [Google Scholar] [CrossRef]
- Devereux, R.; Winfrey, M.R.; Winfrey, J.; Stahl, D.A. Depth profile of sulfate-reducing bacterial ribosomal RNA and mercury methylation in an estuarine sediment. FEMS Microbiol. Ecol. 1996, 20, 23–31. [Google Scholar] [CrossRef]
- Weston, N.B.; Porubsky, W.P.; Samarkin, V.A.; Erickson, M.; Macavoy, S.E.; Macavoy, S.B. Porewater stoichiometry of terminal metabolic products, sulfate, and dissolved organic carbon and nitrogen in estuarine intertidal creek-bank sediments. Biogeochemistry 2006, 77, 375–408. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Bryan, A.L.; Parks, J.R.; Gibson, K.N. Mercury Accumulation in a Stream Ecosystem: Linking Labile Mercury in Sediment Porewaters to Bioaccumulative Mercury in Trophic Webs. Water 2022, 14, 2003. https://doi.org/10.3390/w14132003
Xu X, Bryan AL, Parks JR, Gibson KN. Mercury Accumulation in a Stream Ecosystem: Linking Labile Mercury in Sediment Porewaters to Bioaccumulative Mercury in Trophic Webs. Water. 2022; 14(13):2003. https://doi.org/10.3390/w14132003
Chicago/Turabian StyleXu, Xiaoyu, Albert L. Bryan, Jasmine R. Parks, and Kara N. Gibson. 2022. "Mercury Accumulation in a Stream Ecosystem: Linking Labile Mercury in Sediment Porewaters to Bioaccumulative Mercury in Trophic Webs" Water 14, no. 13: 2003. https://doi.org/10.3390/w14132003
APA StyleXu, X., Bryan, A. L., Parks, J. R., & Gibson, K. N. (2022). Mercury Accumulation in a Stream Ecosystem: Linking Labile Mercury in Sediment Porewaters to Bioaccumulative Mercury in Trophic Webs. Water, 14(13), 2003. https://doi.org/10.3390/w14132003