
On Escherichia coli Resistance to Fluid Shear Stress and Its Significance for Water Disinfection


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
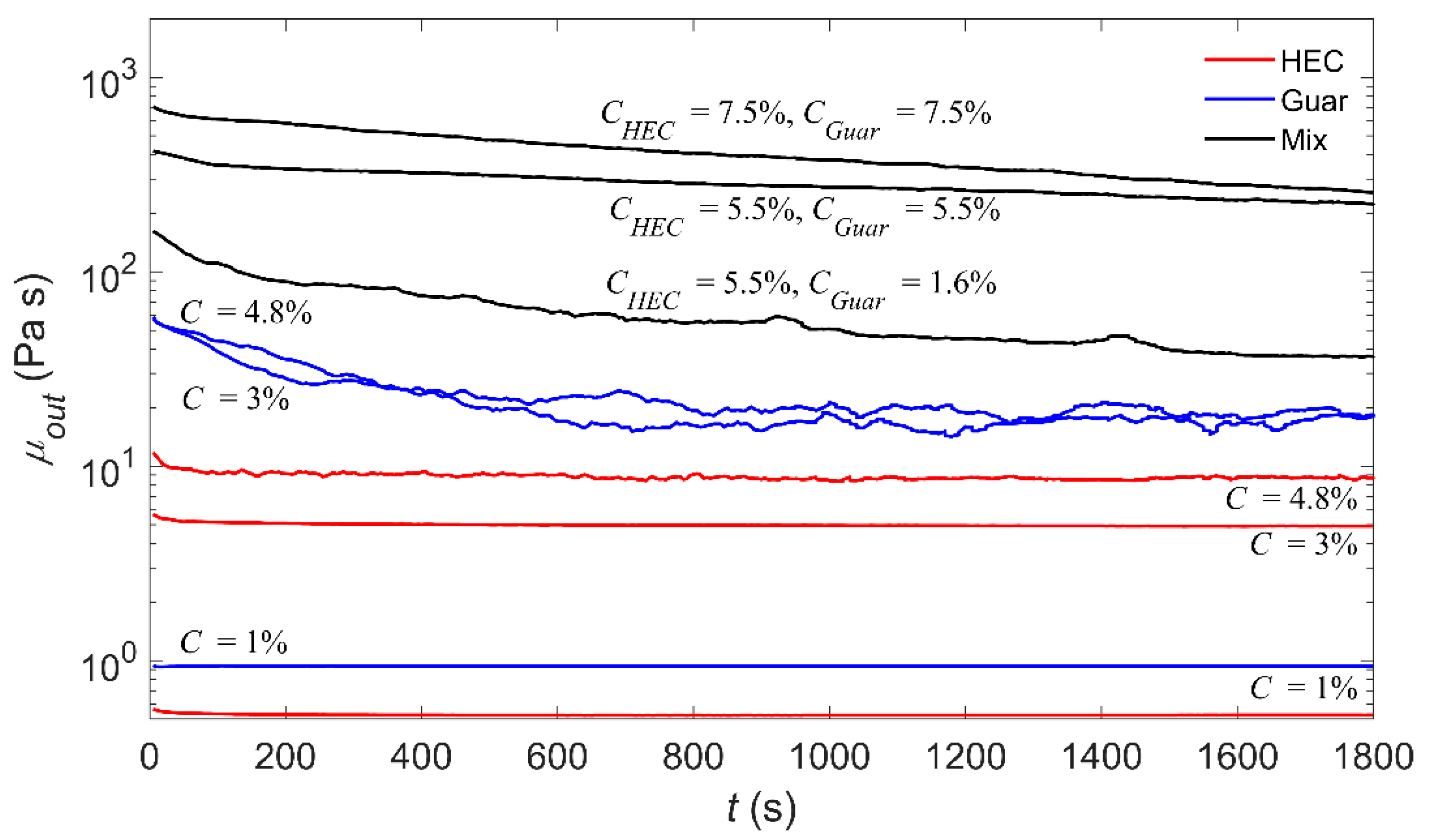
2.2. Substrate Preparation
2.3. Experimental Procedure

2.4. Experimental Procedure
3. Results
4. Discussion
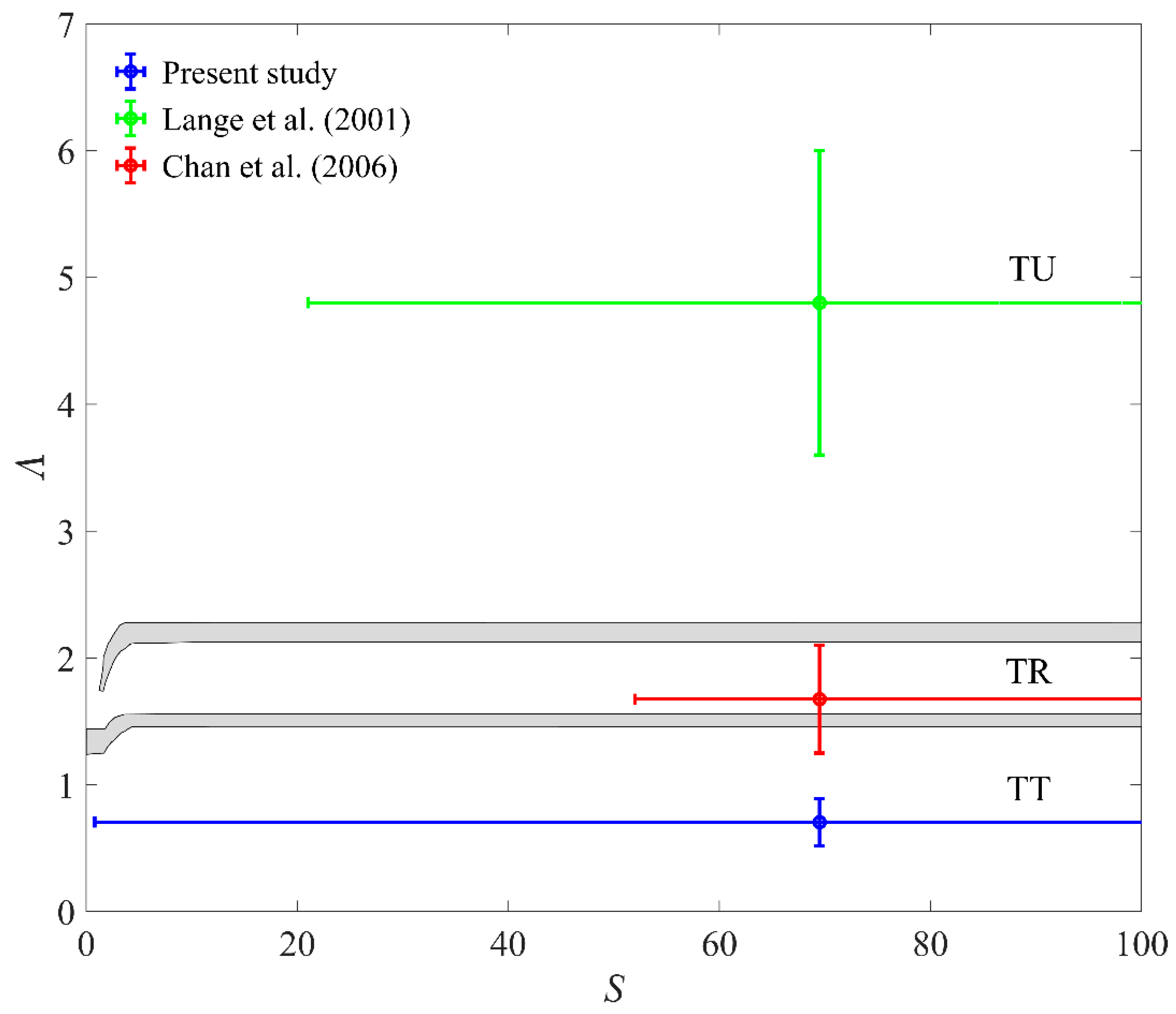
4.1. Comparison with Previous Studies on E.coli Lysis
4.2. Estimating Flow-Induced Stress for E. coli Lysis
4.3. On Modelling Cell Deformation and Dynamics
4.4. Reconciling with the Literature: The Role of Turbulence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, G.C. Handbook of Chlorination and Alternative Disinfectants; Wiley: New York, NY, USA, 2010; ISBN 0470180986. [Google Scholar]
- Li, X.-F.; Mitch, W.A. Drinking Water Disinfection Byproducts (DBPs) and Human Health Effects: Multidisciplinary Challenges and Opportunities. Environ. Sci. Technol. 2018, 52, 1681–1689. [Google Scholar] [CrossRef] [PubMed]
- EU. Directive (EU) 2020/2184 of The European Parliament and of the Council of 16 December 2020 on the Quality of Water Intended for Human Consumption (Recast). Off. J. Eur. Union 2020, L 435/1, 1–62. [Google Scholar]
- U.S. Environmental Protection Agency National Primary Drinking Water Regulations. Available online: https://www.epa.gov/ground-water-and-drinking-water/national-primary-drinking-water-regulations (accessed on 13 October 2021).
- World Health Organization; The United Nations Children’s Fund. Progress on Drinking Water, Sanitation and Hygiene: 2017 Update and Sdg Baselines; WHO: Geneva, Switzerland; UNICEF: New York, NY, USA, 2017.
- World Bank Group. Reducing Inequalities in Water Supply, Sanitation, and Hygiene in the Era of the Sustainable Development Goals: Synthesis Report of the WASH Poverty Diagnostic Initiative; World Bank Group: Singapore, 2017. [Google Scholar]
- Save, S.S.; Pandit, A.B.; Joshi, J.B. Microbial Cell Disruption: Role of Cavitation. Chem. Eng. J. Biochem. Eng. J. 1994, 55, B67–B72. [Google Scholar] [CrossRef]
- Burzio, E.; Bersani, F.; Caridi, G.C.A.; Vesipa, R.; Ridolfi, L.; Manes, C. Water Disinfection by Orifice-Induced Hydrodynamic Cavitation. Ultrason. Sonochem. 2019, 60, 104740. [Google Scholar] [CrossRef]
- Dular, M.; Griessler-Bulc, T.; Gutierrez-Aguirre, I.; Heath, E.; Kosjek, T.; Klemenčič, A.K.; Oder, M.; Petkovšek, M.; Rački, N.; Ravnikar, M. Use of Hydrodynamic Cavitation in (Waste) Water Treatment. Ultrason. Sonochem. 2016, 29, 577–588. [Google Scholar] [CrossRef]
- Šarc, A.; Kosel, J.; Stopar, D.; Oder, M.; Dular, M. Removal of Bacteria Legionella Pneumophila, Escherichia Coli, and Bacillus Subtilis by (Super) Cavitation. Ultrason. Sonochem. 2018, 42, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Brennen, C.E. Cavitation and Bubble Dynamics; Cambridge University Press: Cambridge, UK, 2014; ISBN 1107644763. [Google Scholar]
- Zupanc, M.; Pandur, Ž.; Perdih, T.S.; Stopar, D.; Petkovšek, M.; Dular, M. Effects of Cavitation on Different Microorganisms: The Current Understanding of the Mechanisms Taking Place behind the Phenomenon. A Review and Proposals for Further Research. Ultrason. Sonochem. 2019, 57, 147–165. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Liu, J.; Ji, L.; Wang, G.; Zhao, S.; Yoon, J.Y.; Chen, S. A Review on Hydrodynamic Cavitation Disinfection: The Current State of Knowledge. Sci. Total Environ. 2020, 737, 139606. [Google Scholar] [CrossRef]
- Zeng, Q.; Gonzalez-Avila, S.R.; Dijkink, R.; Koukouvinis, P.; Gavaises, M.; Ohl, C.-D. Wall Shear Stress from Jetting Cavitation Bubbles. J. Fluid Mech. 2018, 846, 341–355. [Google Scholar] [CrossRef]
- Dijkink, R.; Ohl, C.-D. Measurement of Cavitation Induced Wall Shear Stress. Appl. Phys. Lett. 2008, 93, 254107. [Google Scholar] [CrossRef]
- Sawant, S.S.; Anil, A.C.; Krishnamurthy, V.; Gaonkar, C.; Kolwalkar, J.; Khandeparker, L.; Desai, D.; Mahulkar, A.V.; Ranade, V.V.; Pandit, A.B. Effect of Hydrodynamic Cavitation on Zooplankton: A Tool for Disinfection. Biochem. Eng. J. 2008, 42, 320–328. [Google Scholar] [CrossRef]
- Schiffer, A.; Gardner, M.N.; Lynn, R.H.; Tagarielli, V.L. A New Apparatus to Induce Lysis of Planktonic Microbial Cells by Shock Compression, Cavitation and Spray. R. Soc. Open Sci. 2017, 4, 160939. [Google Scholar] [CrossRef] [PubMed]
- Rikmanis, M.; Berzinš, A.; Viesturs, U. Excess Turbulence as a Cause of Turbohypobiosis in Cultivation of Microorganisms. Cent. Eur. J. Biol. 2007, 2, 481–501. [Google Scholar] [CrossRef]
- Wang, C.; Lan, C.Q. Effects of Shear Stress on Microalgae—A Review. Biotechnol. Adv. 2018, 36, 986–1002. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.N.; Qiao, Q.S.; Lee, J.H.W.; Choi, K.W.; Huang, J.-C. Modeling of Mixing and Rapid Chlorine Demand in Sewage Disinfection with Dense Chlorine Jets. J. Environ. Eng. 2017, 143, 4017074. [Google Scholar] [CrossRef]
- Pourbozorg, M.; Li, T.; Law, A.W.K. Effect of Turbulence on Fouling Control of Submerged Hollow Fibre Membrane Filtration. Water Res. 2016, 99, 101–111. [Google Scholar] [CrossRef]
- Al-Homoud, A.; Hondzo, M. Enhanced Uptake of Dissolved Oxygen and Glucose by Escherichia Coli in a Turbulent Flow. Appl. Microbiol. Biotechnol. 2008, 79, 643–655. [Google Scholar] [CrossRef]
- Hondzo, M.; Al-Homoud, A. Model Development and Verification for Mass Transport to Escherichia Coli Cells in a Turbulent Flow. Water Resour. Res. 2007, 43, 1–11. [Google Scholar] [CrossRef]
- Hondzo, M.; Wüest, A. Do Microscopic Organisms Feel Turbulent Flows? Environ. Sci. Technol. 2009, 43, 764–768. [Google Scholar] [CrossRef]
- Durham, W.M.; Kessler, J.O.; Stocker, R. Disruption of Vertical Motility by Shear Triggers Formation of Thin Phytoplankton Layers. Science 2009, 323, 1067–1070. [Google Scholar] [CrossRef]
- Durham, W.M.; Climent, E.; Barry, M.; De Lillo, F.; Boffetta, G.; Cencini, M.; Stocker, R. Turbulence Drives Microscale Patches of Motile Phytoplankton. Nat. Commun. 2013, 4, 2148. [Google Scholar] [CrossRef] [PubMed]
- Lange, H.; Taillandier, P.; Riba, J. Effect of High Shear Stress on Microbial Viability. J. Chem. Technol. Biotechnol. 2001, 76, 501–505. [Google Scholar] [CrossRef]
- Chan, G.; Booth, A.J.; Mannweiler, K.; Hoare, M. Ultra Scale-down Studies of the Effect of Flow and Impact Conditions during E. Coli Cell Processing. Biotechnol. Bioeng. 2006, 95, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Mok, J.H.; Pyatkovskyy, T.; Yousef, A.; Sastry, S.K. Combined Effect of Shear Stress and Moderate Electric Field on the Inactivation of Escherichia Coli K12 in Apple Juice. J. Food Eng. 2019, 262, 121–130. [Google Scholar] [CrossRef]
- Goldberg, A.L.; Swamy, K.H.S.; Chung, C.H.; Larimore, F.S. Proteases in Escherichia Coli. Methods Enzymol. 1981, 80, 680–702. [Google Scholar]
- Balasundaram, B.; Harrison, S.T.L. Study of Physical and Biological Factors Involved in the Disruption of E. coli by Hydrodynamic Cavitation. Biotechnol. Prog. 2006, 22, 907–913. [Google Scholar] [CrossRef]
- Atis, S.; Weinstein, B.T.; Murray, A.W.; Nelson, D.R. Microbial Range Expansions on Liquid Substrates. Phys. Rev. X 2019, 9, 21058. [Google Scholar] [CrossRef]
- ISO 9308-2; Water Quality—Enumeration of Escherichia coli and Coliform Bacteria—Part 2: Most Probable Number Method. ISO: Geneva, Switzerland, 2012.
- Vang, Ó.K.; Corfitzen, C.B.; Smith, C.; Albrechtsen, H.-J. Evaluation of ATP Measurements to Detect Microbial Ingress by Wastewater and Surface Water in Drinking Water. Water Res. 2014, 64, 309–320. [Google Scholar] [CrossRef]
- Fricker, E.J.; Illingworth, K.S.; Fricker, C.R. Use of Two Formulations of Colilert and QuantiTrayTM for Assessment of the Bacteriological Quality of Water. Water Res. 1997, 31, 2495–2499. [Google Scholar] [CrossRef]
- Hamilton, W.P.; Kim, M.; Thackston, E.L. Comparison of Commercially Available Escherichia Coli Enumeration Tests: Implications for Attaining Water Quality Standards. Water Res. 2005, 39, 4869–4878. [Google Scholar] [CrossRef]
- Nilsen, V.; Christensen, E.; Myrmel, M.; Heistad, A. Spatio-Temporal Dynamics of Virus and Bacteria Removal in Dual-Media Contact-Filtration for Drinking Water. Water Res. 2019, 156, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Schlichting, H.; Gersten, K. Boundary-Layer Theory, 9th ed.; Springer: Berlin/Heidelberg, Germany, 2017; ISBN 366252919X. [Google Scholar]
- Dufrêne, Y.F.; Persat, A. Mechanomicrobiology: How Bacteria Sense and Respond to Forces. Nat. Rev. Microbiol. 2020, 18, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.R. An Introduction to Error Analysis; University Science Books: Sausalito, CA, USA, 1997; 327p. [Google Scholar]
- Keentok, M.; Xue, S.-C. Edge Fracture in Cone-Plate and Parallel Plate Flows. Rheol. Acta 1999, 38, 321–348. [Google Scholar] [CrossRef]
- Mille, Y.; Beney, L.; Gervais, P. Viability of Escherichia Coli after Combined Osmotic and Thermal Treatment: A Plasma Membrane Implication. Biochim. Biophys. Acta (BBA)-Biomembr. 2002, 1567, 41–48. [Google Scholar] [CrossRef]
- Sperandeo, P.; Martorana, A.M.; Polissi, A. Lipopolysaccharide Biosynthesis and Transport to the Outer Membrane of Gram-Negative Bacteria. Bact. Cell Walls Membr. 2019, 92, 9–37. [Google Scholar]
- Hwang, H.; Paracini, N.; Parks, J.M.; Lakey, J.H.; Gumbart, J.C. Distribution of Mechanical Stress in the Escherichia Coli Cell Envelope. Biochim. Biophys. Acta (BBA)-Biomembr. 2018, 1860, 2566–2575. [Google Scholar] [CrossRef]
- Xie, J.Y.; Ding, G.H.; Karttunen, M. Molecular Dynamics Simulations of Lipid Membranes with Lateral Force: Rupture and Dynamic Properties. Biochim. Biophys. Acta (BBA)-Biomembr. 2014, 1838, 994–1002. [Google Scholar] [CrossRef]
- Gan, L.; Chen, S.; Jensen, G.J. Molecular Organization of Gram-Negative Peptidoglycan. Proc. Natl. Acad. Sci. USA 2008, 105, 18953–18957. [Google Scholar] [CrossRef]
- Mitra, K.; Ubarretxena-Belandia, I.; Taguchi, T.; Warren, G.; Engelman, D.M. Modulation of the Bilayer Thickness of Exocytic Pathway Membranes by Membrane Proteins Rather than Cholesterol. Proc. Natl. Acad. Sci. USA 2004, 101, 4083–4088. [Google Scholar] [CrossRef]
- Inouye, M. A Three-Dimensional Molecular Assembly Model of a Lipoprotein from the Escherichia Coli Outer Membrane. Proc. Natl. Acad. Sci. USA 1974, 71, 2396–2400. [Google Scholar] [CrossRef]
- Willis, M.J.; Ahrens, T.J.; Bertani, L.E.; Nash, C.Z. Bugbuster—Survivability of Living Bacteria upon Shock Compression. Earth Planet. Sci. Lett. 2006, 247, 185–196. [Google Scholar] [CrossRef]
- Faghih, M.M.; Sharp, M.K. Modeling and Prediction of Flow-Induced Hemolysis: A Review. Biomech. Model. Mechanobiol. 2019, 18, 845–881. [Google Scholar] [CrossRef] [PubMed]
- Barthes-Biesel, D. Motion and Deformation of Elastic Capsules and Vesicles in Flow. Annu. Rev. Fluid Mech. 2016, 48, 25–52. [Google Scholar] [CrossRef]
- Chang, K.-S.; Olbricht, W.L. Experimental Studies of the Deformation and Breakup of a Synthetic Capsule in Steady and Unsteady Simple Shear Flow. J. Fluid Mech. 1993, 250, 609–633. [Google Scholar] [CrossRef]
- Lebedev, V.V.; Turitsyn, K.S.; Vergeles, S.S. Dynamics of Nearly Spherical Vesicles in an External Flow. Phys. Rev. Lett. 2007, 99, 218101. [Google Scholar] [CrossRef]
- Deschamps, J.; Kantsler, V.; Steinberg, V. Phase Diagram of Single Vesicle Dynamical States in Shear Flow. Phys. Rev. Lett. 2009, 102, 118105. [Google Scholar] [CrossRef]
- Semeraro, E.F.; Devos, J.M.; Porcar, L.; Forsyth, V.T.; Narayanan, T. In Vivo Analysis of the Escherichia Coli Ultrastructure by Small-Angle Scattering. IUCrJ 2017, 4, 751–757. [Google Scholar] [CrossRef]
- Mullineaux, C.W.; Nenninger, A.; Ray, N.; Robinson, C. Diffusion of Green Fluorescent Protein in Three Cell Environments in Escherichia Coli. J. Bacteriol. 2006, 188, 3442–3448. [Google Scholar] [CrossRef]
- Kelley, E.G.; Butler, P.D.; Ashkar, R.; Bradbury, R.; Nagao, M. Scaling Relationships for the Elastic Moduli and Viscosity of Mixed Lipid Membranes. Proc. Natl. Acad. Sci. USA 2020, 117, 23365–23373. [Google Scholar] [CrossRef]
- Deng, Y.; Sun, M.; Shaevitz, J.W. Direct Measurement of Cell Wall Stress Stiffening and Turgor Pressure in Live Bacterial Cells. Phys. Rev. Lett. 2011, 107, 158101. [Google Scholar] [CrossRef]
- Mathelié-Guinlet, M.; Asmar, A.T.; Collet, J.-F.; Dufrêne, Y.F. Lipoprotein Lpp Regulates the Mechanical Properties of the E. Coli Cell Envelope. Nat. Commun. 2020, 11, 1789. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Briones, M.A.; Chalmers, J.J. Flow Parameters Associated with Hydrodynamic Cell Injury. Biotechnol. Bioeng. 1994, 44, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Meneveau, C.; Sreenivasan, K.R. The Multifractal Nature of Turbulent Energy Dissipation. J. Fluid Mech. 1991, 224, 429–484. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Symbol | Range of Values | Units | Source | Notes |
|---|---|---|---|---|---|
| Thickness of outer membrane | 3.7 | nm | [47] | - | |
| Thickness of inner membrane | 7 | nm | [48] | - | |
| Thickness of peptidoglycan | <4 | nm | [46] | Only maximum value is reported | |
| Major axis | 1–4 | µm | [55] | - | |
| Minor axis | 0.4–0.8 | µm | [55] | - | |
| Cytoplasm viscosity | 9.7 | mPa s | [56] | Used as the cell inner fluid viscosity | |
| Cell envelope viscosity | 30 | nPa s m | [57] | For generic mixed lipid membranes | |
| Elastic modulus of cell envelope | 2–31 | MPa | [58,59] | From atomic force microscopy measurements | |
| Area expansion modulus of lipid membranes | 240 | mN/m | [44] | From experiments with a Langmuir trough | |
| Area expansion modulus of cell wall | 30–500 | mN/m | [44] | From molecular dynamics simulations |
| Present Study with Max (Beginning of Tests) | Present Study with min (End of Tests) | Lange et al. [27] | Chan et al. [28] | |
|---|---|---|---|---|
| 1 × 10−5–1.7 × 10−2 | 2 × 10−5–1.9 × 10−2 | 8.2 | 1.9 | |
| 3 × 10−5–1 × 10−1 | 3 × 10−5–6 × 10−2 | 7 × 10−4–3 × 10−2 | 2 × 10−3–8 × 10−2 | |
| 5 × 10−5–2 × 10−1 | 8 × 10−5–2.1 × 10−1 | 37–94 | 8.8–22 | |
| 0.9–240 | 0.8–387 | 21–118 | 52–296 | |
| 0.52–0.89 | 0.52–0.89 | 3.6–6 | 1.25–2.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vettori, D.; Manes, C.; Dalmazzo, D.; Ridolfi, L. On Escherichia coli Resistance to Fluid Shear Stress and Its Significance for Water Disinfection. Water 2022, 14, 2637. https://doi.org/10.3390/w14172637
Vettori D, Manes C, Dalmazzo D, Ridolfi L. On Escherichia coli Resistance to Fluid Shear Stress and Its Significance for Water Disinfection. Water. 2022; 14(17):2637. https://doi.org/10.3390/w14172637
Chicago/Turabian StyleVettori, Davide, Costantino Manes, Davide Dalmazzo, and Luca Ridolfi. 2022. "On Escherichia coli Resistance to Fluid Shear Stress and Its Significance for Water Disinfection" Water 14, no. 17: 2637. https://doi.org/10.3390/w14172637
APA StyleVettori, D., Manes, C., Dalmazzo, D., & Ridolfi, L. (2022). On Escherichia coli Resistance to Fluid Shear Stress and Its Significance for Water Disinfection. Water, 14(17), 2637. https://doi.org/10.3390/w14172637