
Long-Term Exposure to Polystyrene Nanoplastics Impairs the Liver Health of Medaka

 ,
,  and
and 
Abstract
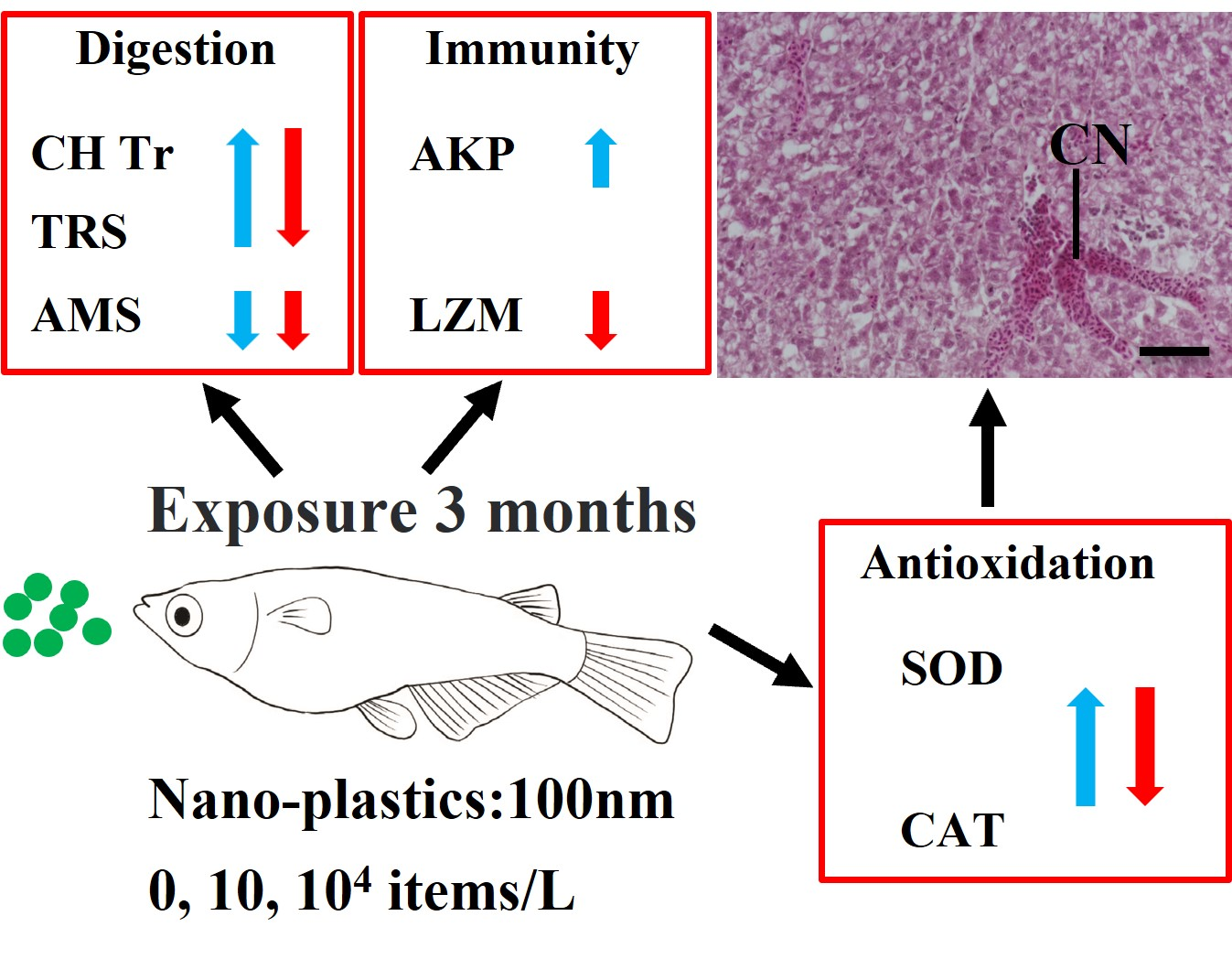
:Highlights
- The hepatotoxic effects of nanoplastics (NPs) are evaluated in medaka fish.
- Digestive, innate immune, and antioxidant enzyme activities are affected by NP exposure.
- Histopathology confirmed direct liver tissue damage by NP exposure.
- We conclude that NPs at as low as 10 items/L can be toxic to medaka in long term.
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Nanoplastics
2.2. Experimental Fish
2.3. Experimental Design
2.4. Liver Histopathology
2.5. Biochemical Analysis
2.6. Statistics
3. Results
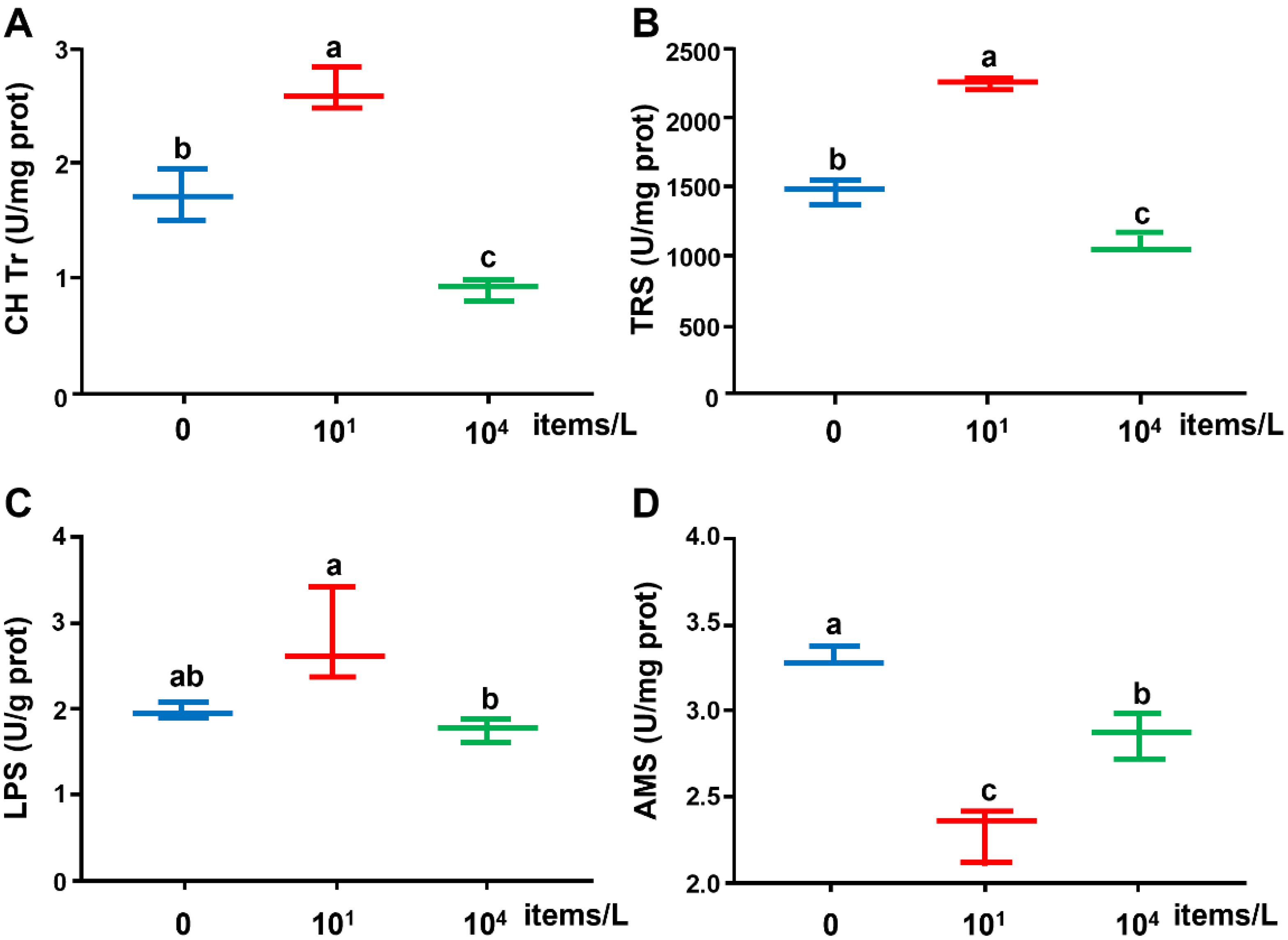
3.1. Digestive Enzyme Activities
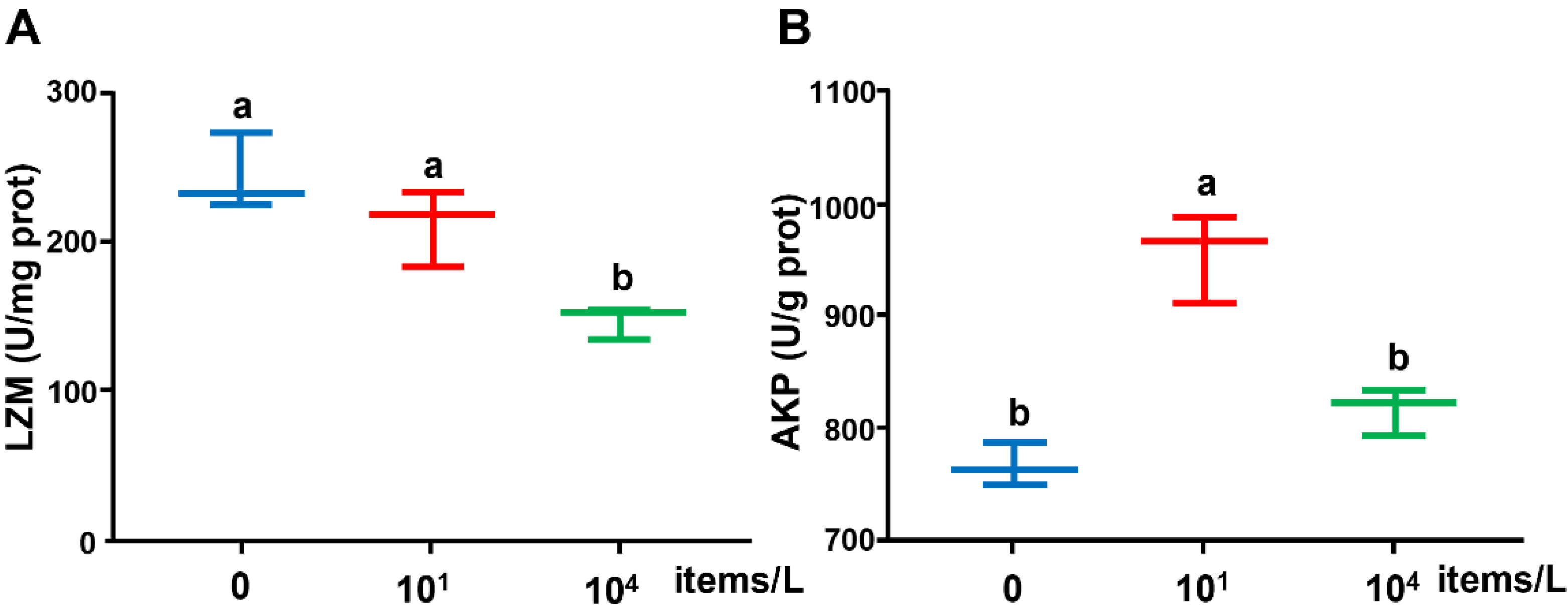
3.2. Innate Immune Enzyme Activities
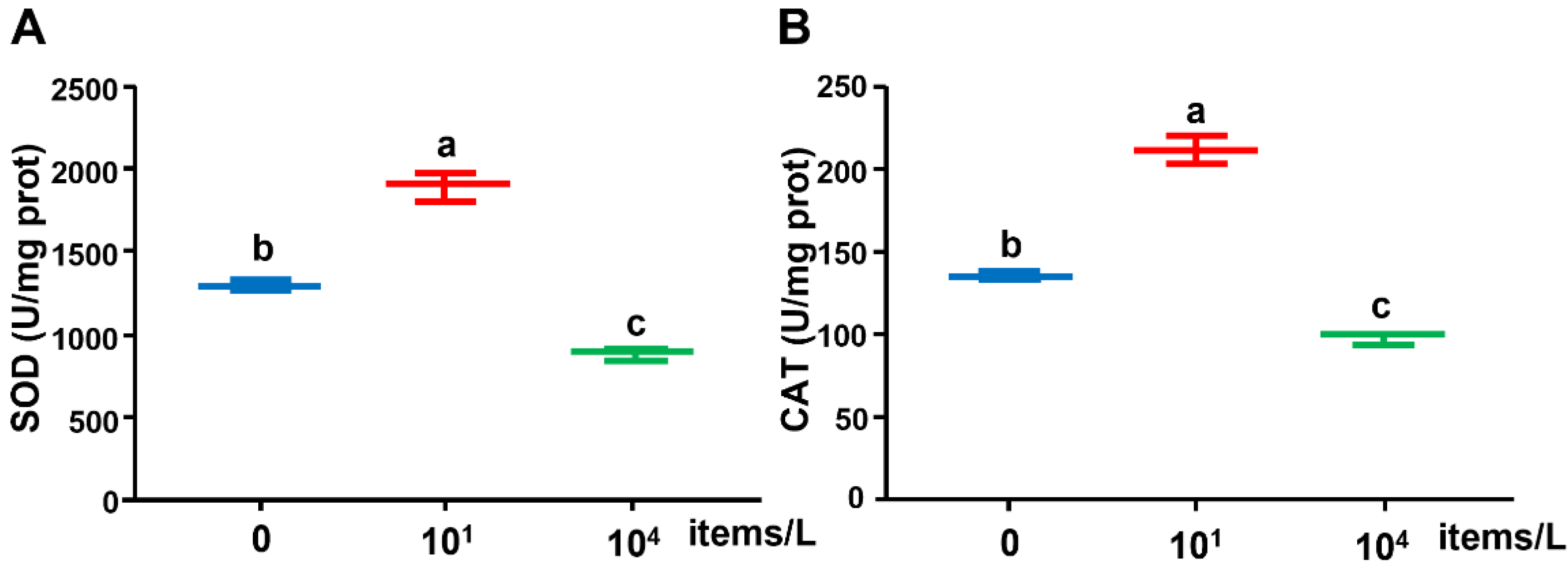
3.3. Antioxidant Enzyme Activities
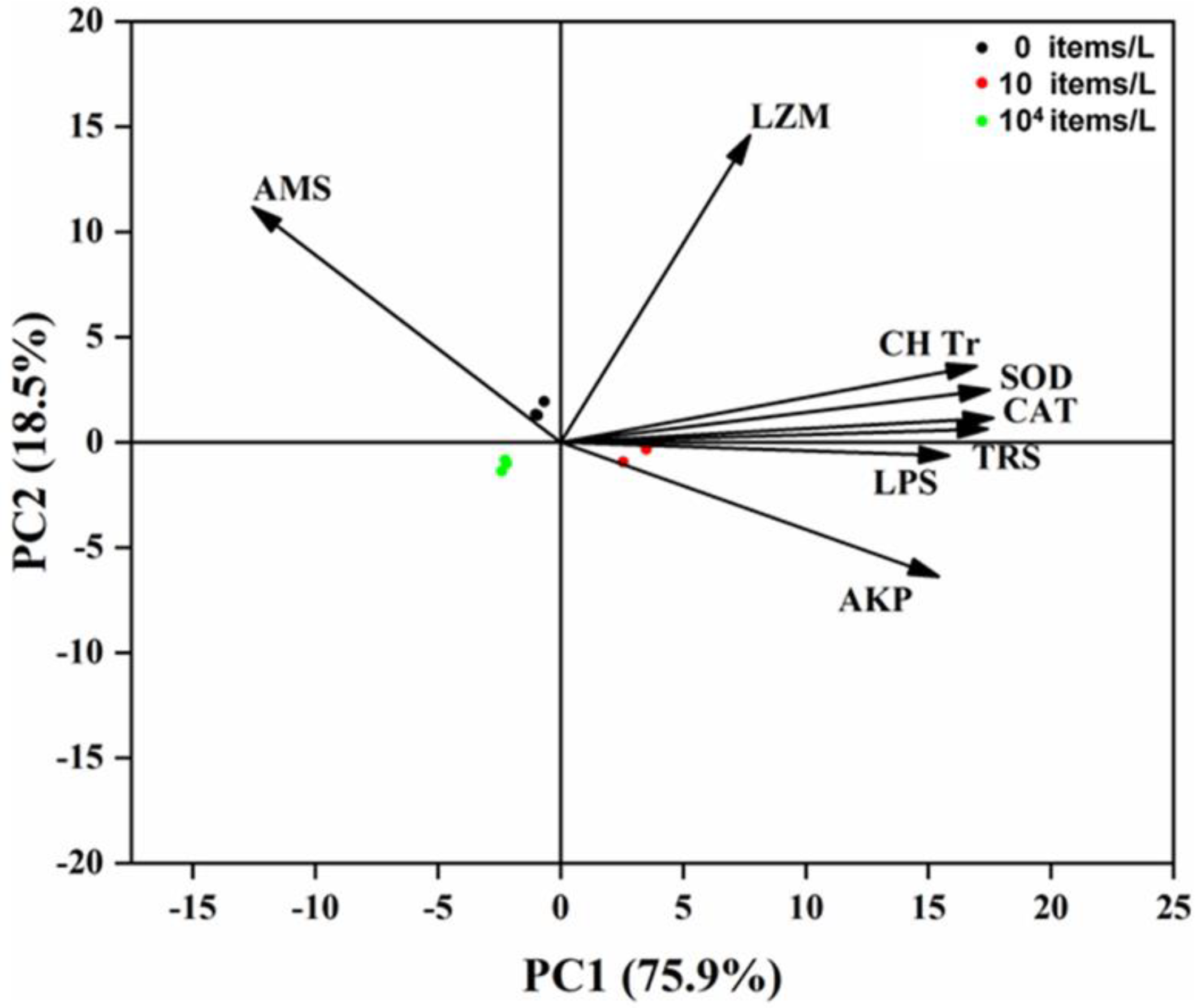
3.4. Principal Component Analysis
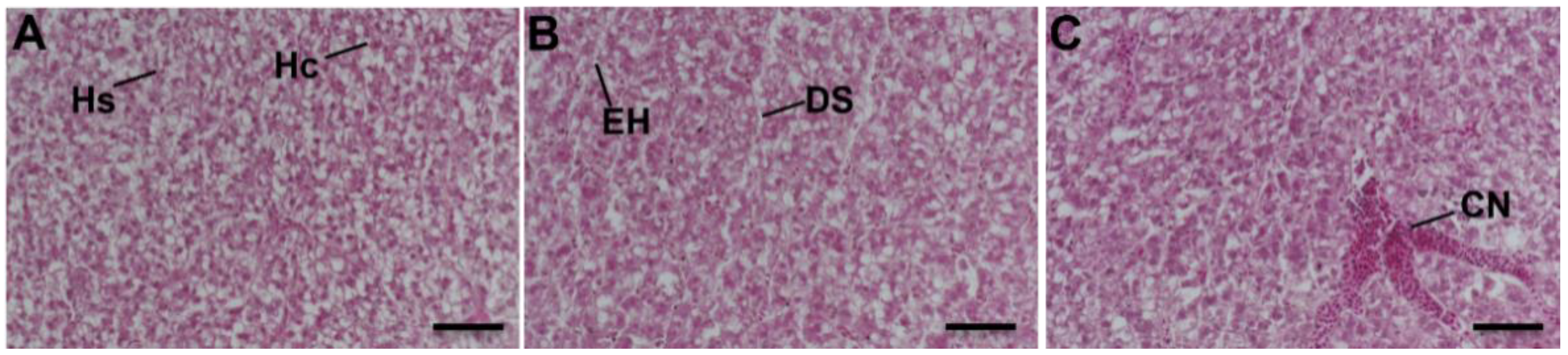
3.5. Liver Histopathology
4. Discussion
4.1. Effects of Long-Term Nanoplastics Exposure on Digestive Enzymes
4.2. Effects of Long-Term Nanoplastics Exposure on Innate Immune Enzymes
4.3. Effects of Long-Term Nanoplastics Exposure on Oxidase Enzymes
4.4. Histopathological Effects of Long-Term Nanoplastics Exposure on Liver
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Y.; Liu, Z.; Li, M.; Jiang, Q.; Wu, D.; Huang, Y.; Jiao, Y.; Zhang, M.; Zhao, Y. Effects of Nanoplastics on Antioxidant and Immune Enzyme Activities and Related Gene Expression in Juvenile Macrobrachium Nipponense. J. Hazard. Mater. 2020, 398, 122990. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Wang, S.; Wang, X.; Yu, X.; Hu, M.; Huang, W.; Wang, Y. Nanoplastics Impair the Intestinal Health of the Juvenile Large Yellow Croaker Larimichthys Crocea. J. Hazard. Mater. 2020, 397, 122773. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Ma, J.; Ji, R.; Pan, K.; Miao, A.J. Microplastics in Aquatic Environments: Occurrence, Accumulation, and Biological Effects. Sci. Total Environ. 2020, 703, 134699. [Google Scholar] [CrossRef] [PubMed]
- Shim, W.J.; Thomposon, R.C. Microplastics in the Ocean. Arch. Environ. Contam. Toxicol. 2015, 69, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Chen, Q.; Jiang, W.; Wang, L.; Xie, H.; Kalogerakis, N.; Ma, Y.; Ji, R. A Carbon-14 Radiotracer-Based Study on the Phototransformation of Polystyrene Nanoplastics in Water: Versus in Air. Environ. Sci. Nano 2019, 6, 2907–2917. [Google Scholar] [CrossRef]
- Zhang, B.; Chao, J.; Chen, L.; Liu, L.; Yang, X.; Wang, Q. Research Progress of Nanoplastics in Freshwater. Sci. Total Environ. 2021, 757, 143791. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, Y.; Deng, Y.; Jiang, W.; Zhao, Y.; Geng, J.; Ding, L.; Ren, H. Uptake and Accumulation of Polystyrene Microplastics in Zebrafish (Danio rerio) and Toxic Effects in Liver. Environ. Sci. Technol. 2016, 50, 4054–4060. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, B.; Bezirci, G.; Çağan, A.S.; Coppens, J.; Levi, E.E.; Oluz, Z.; Tuncel, E.; Duran, H.; Beklioğlu, M. Food Web Effects of Titanium Dioxide Nanoparticles in an Outdoor Freshwater Mesocosm Experiment. Nanotoxicology 2016, 10, 902–912. [Google Scholar] [CrossRef]
- Xie, S.; Zhou, A.; Wei, T.; Li, S.; Yang, B.; Xu, G.; Zou, J. Nanoplastics Induce More Serious Microbiota Dysbiosis and Inflammation in the Gut of Adult Zebrafish than Microplastics. Bull. Environ. Contam. Toxicol. 2021, 107, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, C.; Cuesta, A.; Esteban, M.Á. Effects of Dietary Polyvinylchloride Microparticles on General Health, Immune Status and Expression of Several Genes Related to Stress in Gilthead Seabream (Sparus aurata L.). Fish Shellfish Immunol. 2017, 68, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Romano, N.; Ashikin, M.; Teh, J.C.; Syukri, F.; Karami, A. Effects of Pristine Polyvinyl Chloride Fragments on Whole Body Histology and Protease Activity in Silver Barb Barbodes gonionotus Fry. Environ. Pollut. 2018, 237, 1106–1111. [Google Scholar] [CrossRef]
- Jin, Y.; Zhang, X.; Shu, L.; Chen, L.; Sun, L.; Qian, H.; Liu, W.; Fu, Z. Oxidative Stress Response and Gene Expression with Atrazine Exposure in Adult Female Zebrafish (Danio rerio). Chemosphere 2010, 78, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yan, L.; Junaid, M.; Chen, X.; Liao, H.; Gao, D.; Wang, Q.; Zhang, Y.; Wang, J. Impacts of Polystyrene Nanoplastics on the Oxidative Stress, Immune Responses, and Gut Microbiota to Grass Carp (Ctenopharyngodon idella). SSRN Electron. J. 2022, 1–27. [Google Scholar] [CrossRef]
- Li, Z.; Feng, C.; Wu, Y.; Guo, X. Impacts of Nanoplastics on Bivalve: Fluorescence Tracing of Organ Accumulation, Oxidative Stress and Damage. J. Hazard. Mater. 2020, 392, 122418. [Google Scholar] [CrossRef] [PubMed]
- Brandts, I.; Teles, M.; Tvarijonaviciute, A.; Pereira, M.L.; Martins, M.A.; Tort, L.; Oliveira, M. Effects of Polymethylmethacrylate Nanoplastics on Dicentrarchus Labrax. Genomics 2018, 110, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Ding, R.; Ma, Y.; Sun, Q.; Ren, X.; Sun, Z.; Duan, J. Cardiovascular Toxicity Assessment of Polyethylene Nanoplastics on Developing Zebrafish Embryos. Chemosphere 2021, 282, 131124. [Google Scholar] [CrossRef]
- Mattsson, K.; Johnson, E.V.; Malmendal, A.; Linse, S.; Hansson, L.A.; Cedervall, T. Brain Damage and Behavioural Disorders in Fish Induced by Plastic Nanoparticles Delivered through the Food Chain. Sci. Rep. 2017, 7, 11452. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Cai, M.; Wu, D.; Yu, P.; Jiao, Y.; Jiang, Q.; Zhao, Y. Effects of Nanoplastics at Predicted Environmental Concentration on Daphnia Pulex after Exposure through Multiple Generations. Environ. Pollut. 2020, 256, 113506. [Google Scholar] [CrossRef] [PubMed]
- Botterell, Z.L.R.; Beaumont, N.; Dorrington, T.; Steinke, M.; Thompson, R.C.; Lindeque, P.K. Bioavailability and Effects of Microplastics on Marine Zooplankton: A Review. Environ. Pollut. 2019, 245, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Rist, S.; Baun, A.; Hartmann, N.B. Ingestion of Micro- and Nanoplastics in Daphnia Magna—Quantification of Body Burdens and Assessment of Feeding Rates and Reproduction. Environ. Pollut. 2017, 228, 398–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comfort, K.K.; Braydich-Stolle, L.K.; Maurer, E.I.; Hussain, S.M. Less Is More: Long-Term in vitro Exposure to Low Levels of Silver Nanoparticles Provides New Insights for Nanomaterial Evaluation. ACS Nano 2014, 8, 3260–3271. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Iwasaki, N.; Chen, K.; Shimasaki, Y.; Oshima, Y. Tributyltin and Perfluorooctane Sulfonate Play a Synergistic Role in Promoting Excess Fat Accumulation in Japanese Medaka (Oryzias latipes) via in ovo Exposure. Chemosphere 2019, 220, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Gu, H.; Chang, X.; Huang, W.; Sokolova, I.M.; Wei, S.; Sun, L.; Li, S.; Wang, X.; Hu, M.; et al. Oxidative Stress Induced by Nanoplastics in the Liver of Juvenile Large Yellow Croaker Larimichthys crocea. Mar. Pollut. Bull. 2021, 170, 112661. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jia, X.; Zhu, H.; Zhang, Q.; He, Y.; Shen, Y.; Xu, X.; Li, J. The Effects of Exposure to Microplastics on Grass Carp (Ctenopharyngodon Idella) at the Physiological, Biochemical, and Transcriptomic Levels. Chemosphere 2022, 286, 131831. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Q.; Le Manach, S.; Sotton, B.; Huet, H.; Duvernois-Berthet, E.; Paris, A.; Duval, C.; Ponger, L.; Marie, A.; Blond, A.; et al. Deep Sexual Dimorphism in Adult Medaka Fish Liver Highlighted by Multi-Omic Approach. Sci. Rep. 2016, 6, 32459. [Google Scholar] [CrossRef] [PubMed]
- Mathalon, A.; Hill, P. Microplastic Fibers in the Intertidal Ecosystem Surrounding Halifax Harbor, Nova Scotia. Mar. Pollut. Bull. 2014, 81, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Liu, Z.; Wu, D.; Chen, M.; Lv, W.; Zhao, Y. Accumulation of Polystyrene Microplastics in Juvenile Eriocheir Sinensis and Oxidative Stress Effects in the Liver. Aquat. Toxicol. 2018, 200, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Gui, L.; Liu, R.; Hong, Y.; Li, M. Medaka Oct4 Is Essential for Gastrulation, Central Nervous System Development and Angiogenesis. Gene 2020, 733, 144270. [Google Scholar] [CrossRef]
- Candiano, G.; Bruschi, M.; Musante, L.; Santucci, L.; Ghiggeri, G.M.; Carnemolla, B.; Orecchia, P.; Zardi, L.; Righetti, P.G. Blue Silver: A Very Sensitive Colloidal Coomassie G-250 Staining for Proteome Analysis. Electrophoresis 2004, 25, 1327–1333. [Google Scholar] [CrossRef]
- Wang, Q.; Huang, F.; Liang, K.; Niu, W.; Duan, X.; Jia, X.; Wu, X.; Xu, P.; Zhou, L. Polystyrene Nanoplastics Affect Digestive Function and Growth in Juvenile Groupers. Sci. Total Environ. 2022, 808, 152098. [Google Scholar] [CrossRef]
- Trestrail, C.; Walpitagama, M.; Miranda, A.; Nugegoda, D.; Shimeta, J. Microplastics Alter Digestive Enzyme Activities in the Marine Bivalve, Mytilus Galloprovincialis. Sci. Total Environ. 2021, 779, 146418. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.L.; Rowe, D.; Thompson, R.C.; Galloway, T.S. Microplastic Ingestion Decreases Energy Reserves in Marine Worms. Curr. Biol. 2013, 23, R1031–R1033. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.N.; Wen, B.; Zhu, J.G.; Zhang, Y.S.; Gao, J.Z.; Chen, Z.Z. Exposure to Microplastics Impairs Digestive Performance, Stimulates Immune Response and Induces Microbiota Dysbiosis in the Gut of Juvenile Guppy (Poecilia Reticulata). Sci. Total Environ. 2020, 733, 138929. [Google Scholar] [CrossRef] [PubMed]
- Cara, B.; Moyano, F.J.; Zambonino, J.L.; Fauvel, C. Trypsin and Chymotrypsin as Indicators of Nutritional Status of Post-Weaned Sea Bass Larvae. J. Fish Biol. 2007, 70, 1798–1808. [Google Scholar] [CrossRef]
- Feng, S.; Zeng, Y.; Cai, Z.; Wu, J.; Chan, L.L.; Zhu, J.; Zhou, J. Polystyrene Microplastics Alter the Intestinal Microbiota Function and the Hepatic Metabolism Status in Marine Medaka (Oryzias melastigma). Sci. Total Environ. 2021, 759, 143558. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Gui, L.; Wei, W.; Xu, E.G.; Zhu, Y.; Zhou, W.; Sokolova, I.M.; Li, M.; Wang, Y. Surprisingly Low Particle Concentrations of Nanoplastics Impair the Gut Health of Medaka. Environ. Sci. Nano 2022. in peer review. [Google Scholar]
- Wang, X.; Huang, W.; Wei, S.; Shang, Y.; Gu, H.; Wu, F.; Lan, Z.; Hu, M.; Shi, H.; Wang, Y. Microplastics Impair Digestive Performance but Show Little Effects on Antioxidant Activity in Mussels under Low PH Conditions. Environ. Pollut. 2020, 258, 113691. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Wu, F.; Jiang, X.; Wang, T.; Hu, M.; Chen, J.; Huang, W.; Bao, Y.; Wang, Y. Nano-TiO2 Impairs Digestive Enzyme Activities of Marine Mussels under Ocean Acidification. Chemosphere 2019, 237, 124561. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Li, H.; Wang, X.; Li, X.; Liu, Y.; Sun, J.; Liu, C. A Review of the Immune Molecules in the Sea Cucumber. Fish Shellfish Immunol. 2015, 44, 1–11. [Google Scholar] [CrossRef]
- Jiang, J.; Zhou, Z.; Dong, Y.; Jiang, B.; Chen, Z.; Yang, A.; Wang, B.; Guan, X.; Gao, S.; Sun, H. The in vitro Effects of Divalent Metal Ions on the Activities of Immune-Related Enzymes in Coelomic Fluid from the Sea Cucumber Apostichopus japonicus. Aquac. Res. 2016, 47, 1269–1276. [Google Scholar] [CrossRef]
- Wang, G.; Wang, T.; Zhang, X.; Chen, J.; Feng, C.; Yun, S.; Cheng, Y.; Cheng, F.; Cao, J. Sex-Specific Effects of Fluoride and Lead Exposures on Histology, Antioxidant Physiology, and Immune System in the Liver of Zebrafish (Danio rerio). Ecotoxicology 2022, 31, 396–414. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Rong, J.; Guan, X.; Zha, S.; Shi, W.; Han, Y.; Du, X.; Wu, F.; Huang, W.; Liu, G. Immunotoxicity of Microplastics and Two Persistent Organic Pollutants Alone or in Combination to a Bivalve Species. Environ. Pollut. 2020, 258, 113845. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yu, P.; Cai, M.; Wu, D.; Zhang, M.; Chen, M.; Zhao, Y. Effects of Microplastics on the Innate Immunity and Intestinal Microflora of Juvenile Eriocheir Sinensis. Sci. Total Environ. 2019, 685, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Giri, S.S.; Jun, J.W.; Kim, H.J.; Kim, S.W.; Yun, S.; Park, S.C. Effects of Algal Toxin Okadaic Acid on the Non-Specific Immune and Antioxidant Response of Bay Scallop (Argopecten irradians). Fish Shellfish Immunol. 2017, 65, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Lin, T.; Zhan, W. Variations of Enzyme Activities in the Haemocytes of Scallop Chlamys Farreri after Infection with the Acute Virus Necrobiotic Virus (AVNV). Fish Shellfish Immunol. 2008, 25, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Canesi, L.; Ciacci, C.; Bergami, E.; Monopoli, M.P.; Dawson, K.A.; Papa, S.; Canonico, B.; Corsi, I. Evidence for Immunomodulation and Apoptotic Processes Induced by Cationic Polystyrene Nanoparticles in the Hemocytes of the Marine Bivalve Mytilus. Mar. Environ. Res. 2015, 111, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Jin, S.R.; Chen, Z.Z.; Gao, J.Z.; Liu, Y.N.; Liu, J.H.; Feng, X.S. Single and Combined Effects of Microplastics and Cadmium on the Cadmium Accumulation, Antioxidant Defence and Innate Immunity of the Discus Fish (Symphysodon Aequifasciatus). Environ. Pollut. 2018, 243, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.B.; Wang, G.; Chan, S.F. Effects of Artificial Infection of Litopenaeus Vannamei by Micrococcus Ysodeikticus and WSSV on the Activity of Immunity Related Enzymes. Fish Shellfish Immunol. 2015, 46, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Palić, D. Micro- and Nano-Plastics Activation of Oxidative and Inflammatory Adverse Outcome Pathways. Redox Biol. 2020, 37, 101620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shen, H.; Wang, X.; Wu, J.; Xue, Y. Effects of Chronic Exposure of 2,4-Dichlorophenol on the Antioxidant System in Liver of Freshwater Fish Carassius auratus. Chemosphere 2004, 55, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, D.R. Contaminant-Stimulated Reactive Oxygen Species Production and Oxidative Damage in Aquatic Organisms. Mar. Pollut. Bull. 2001, 42, 656–666. [Google Scholar] [CrossRef]
- Egea, J.; Fabregat, I.; Frapart, Y.M.; Ghezzi, P.; Görlach, A.; Kietzmann, T.; Kubaichuk, K.; Knaus, U.G.; Lopez, M.G.; Olaso-Gonzalez, G.; et al. European Contribution to the Study of ROS: A Summary of the Findings and Prospects for the Future from the COST Action BM1203 (EU-ROS). Redox Biol. 2017, 13, 94–162. [Google Scholar] [CrossRef] [PubMed]
- Ajima, M.N.O.; Pandey, P.K.; Kumar, K.; Poojary, N. Neurotoxic Effects, Molecular Responses and Oxidative Stress Biomarkers in Nile Tilapia, Oreochromis niloticus (Linnaeus, 1758) Exposed to Verapamil. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 196, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Dorantes-Aranda, J.J.; Seger, A.; Mardones, J.I.; Nichols, P.D.; Hallegraeff, G.M. Progress in Understanding Algal Bloom-Mediated Fish Kills: The Role of Superoxide Radicals, Phycotoxins and Fatty Acids. PLoS ONE 2015, 10, e0133549. [Google Scholar]
- de Andrade, L.L.; do Espirito Santo Pereira, A.; Fernandes Fraceto, L.; Bueno dos Reis Martinez, C. Can Atrazine Loaded Nanocapsules Reduce the Toxic Effects of This Herbicide on the Fish Prochilodus Lineatus? A Multibiomarker Approach. Sci. Total Environ. 2019, 663, 548–559. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Yang, Y.; Tao, Y.; Jiang, Y.; Chen, B.; Zhu, X.; Cai, Z.; Li, B. A Mechanism Study on Toxicity of Graphene Oxide to Daphnia Magna: Direct Link between Bioaccumulation and Oxidative Stress. Environ. Pollut. 2018, 234, 953–959. [Google Scholar] [CrossRef]
- Company, R.; Serafim, A.; Cosson, R.P.; Fiala-Médioni, A.; Camus, L.; Colaço, A.; Serrão-Santos, R.; Bebianno, M.J. Antioxidant Biochemical Responses to Long-Term Copper Exposure in Bathymodiolus azoricus from Menez-Gwen Hydrothermal Vent. Sci. Total Environ. 2008, 389, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Ullah, S.; Li, Z.; Hasan, Z.; Khan, S.U.; Fahad, S. Malathion Induced Oxidative Stress Leads to Histopathological and Biochemical Toxicity in the Liver of Rohu (Labeo rohita, Hamilton) at Acute Concentration. Ecotoxicol. Environ. Saf. 2018, 161, 270–280. [Google Scholar] [CrossRef]
- Bucke, D.; Venthaak, D.; Lang, T.; Mellergaard, S. Times No. 19 Common Diseases and Parasites of Fish in the North Atlantic: Training Guide for Identification; International Council for the Exploration of the Sea (ICES): Copenhagen, Denmark, 1996. [Google Scholar]
- Ullah, S.; Begum, M.; Dhama, K.; Ahmad, S.; Hassan, S.; Alam, I. Malathion Induced DNA Damage in Freshwater Fish, Labeo Rohita (Hamilton, 1822) Using Alkaline Single Cell Gel Electrophoresis. Asian J. Anim. Vet. Adv. 2016, 11, 98–105. [Google Scholar] [CrossRef]
- Ahmed, M.K.; Habibullah-Al-Mamun, M.; Parvin, E.; Akter, M.S.; Khan, M.S. Arsenic Induced Toxicity and Histopathological Changes in Gill and Liver Tissue of Freshwater Fish, Tilapia (Oreochromis mossambicus). Exp. Toxicol. Pathol. 2013, 65, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Menard, A.; Drobne, D.; Jemec, A. Ecotoxicity of Nanosized TiO2. Review of in Vivo Data. Environ. Pollut. 2011, 159, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Anderson, D.; Dhawan, A. Zinc Oxide Nanoparticles Induce Oxidative DNA Damage and ROS-Triggered Mitochondria Mediated Apoptosis in Human Liver Cells (HepG2). Apoptosis 2012, 17, 852–870. [Google Scholar] [CrossRef] [PubMed]
- Decuzzi, P.; Godin, B.; Tanaka, T.; Lee, S.Y.; Chiappini, C.; Liu, X.; Ferrari, M. Size and Shape Effects in the Biodistribution of Intravascularly Injected Particles. J. Control. Release 2010, 141, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Devi, G.P.; Ahmed, K.B.A.; Varsha, M.K.N.S.; Shrijha, B.S.; Lal, K.K.S.; Anbazhagan, V.; Thiagarajan, R. Sulfidation of Silver Nanoparticle Reduces Its Toxicity in Zebrafish. Aquat. Toxicol. 2015, 158, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Rossbach, L.M.; Shaw, B.J.; Piegza, D.; Vevers, W.F.; Atfield, A.J.; Handy, R.D. Sub-Lethal Effects of Waterborne Exposure to Copper Nanoparticles Compared to Copper Sulphate on the Shore Crab (Carcinus maenas). Aquat. Toxicol. 2017, 191, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Ben Ameur, W.; de Lapuente, J.; El Megdiche, Y.; Barhoumi, B.; Trabelsi, S.; Camps, L.; Serret, J.; Ramos-López, D.; Gonzalez-Linares, J.; Driss, M.R.; et al. Oxidative Stress, Genotoxicity and Histopathology Biomarker Responses in Mullet (Mugil Cephalus) and Sea Bass (Dicentrarchus labrax) Liver from Bizerte Lagoon (Tunisia). Mar. Pollut. Bull. 2012, 64, 241–251. [Google Scholar] [CrossRef]
- Brandts, I.; Cánovas, M.; Tvarijonaviciute, A.; Llorca, M.; Vega, A.; Farré, M.; Pastor, J.; Roher, N.; Teles, M. Nanoplastics Are Bioaccumulated in Fish Liver and Muscle and Cause DNA Damage after a Chronic Exposure. Environ. Res. 2022, 212, 113433. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Zhao, L.; Xu, H.; Xu, E.G.; Li, M.; Wang, Y. Long-Term Exposure to Polystyrene Nanoplastics Impairs the Liver Health of Medaka. Water 2022, 14, 2767. https://doi.org/10.3390/w14172767
Zhou Y, Zhao L, Xu H, Xu EG, Li M, Wang Y. Long-Term Exposure to Polystyrene Nanoplastics Impairs the Liver Health of Medaka. Water. 2022; 14(17):2767. https://doi.org/10.3390/w14172767
Chicago/Turabian StyleZhou, Yinfeng, Li Zhao, Haijing Xu, Elvis Genbo Xu, Mingyou Li, and Youji Wang. 2022. "Long-Term Exposure to Polystyrene Nanoplastics Impairs the Liver Health of Medaka" Water 14, no. 17: 2767. https://doi.org/10.3390/w14172767
APA StyleZhou, Y., Zhao, L., Xu, H., Xu, E. G., Li, M., & Wang, Y. (2022). Long-Term Exposure to Polystyrene Nanoplastics Impairs the Liver Health of Medaka. Water, 14(17), 2767. https://doi.org/10.3390/w14172767