
Nitrite Stress Induces Oxidative Stress and Leads to Muscle Quality Decreased in Wuchang Bream (Megalobrama amblycephala Yih) Juveniles


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Fish and Rearing Conditions
2.2. Nitrite Exposure and Sample Collection
2.3. Oxidative Stress Biomarkers
2.3.1. Physiological and Biochemical Indicators
2.3.2. Antioxidant Capacity
2.4. Determination of Related Indices of Muscle Metabolism
2.4.1. Metabolism Analysis
2.4.2. Measurement of pH
2.4.3. Textural Parameters
2.5. Statistical Analysis
3. Result
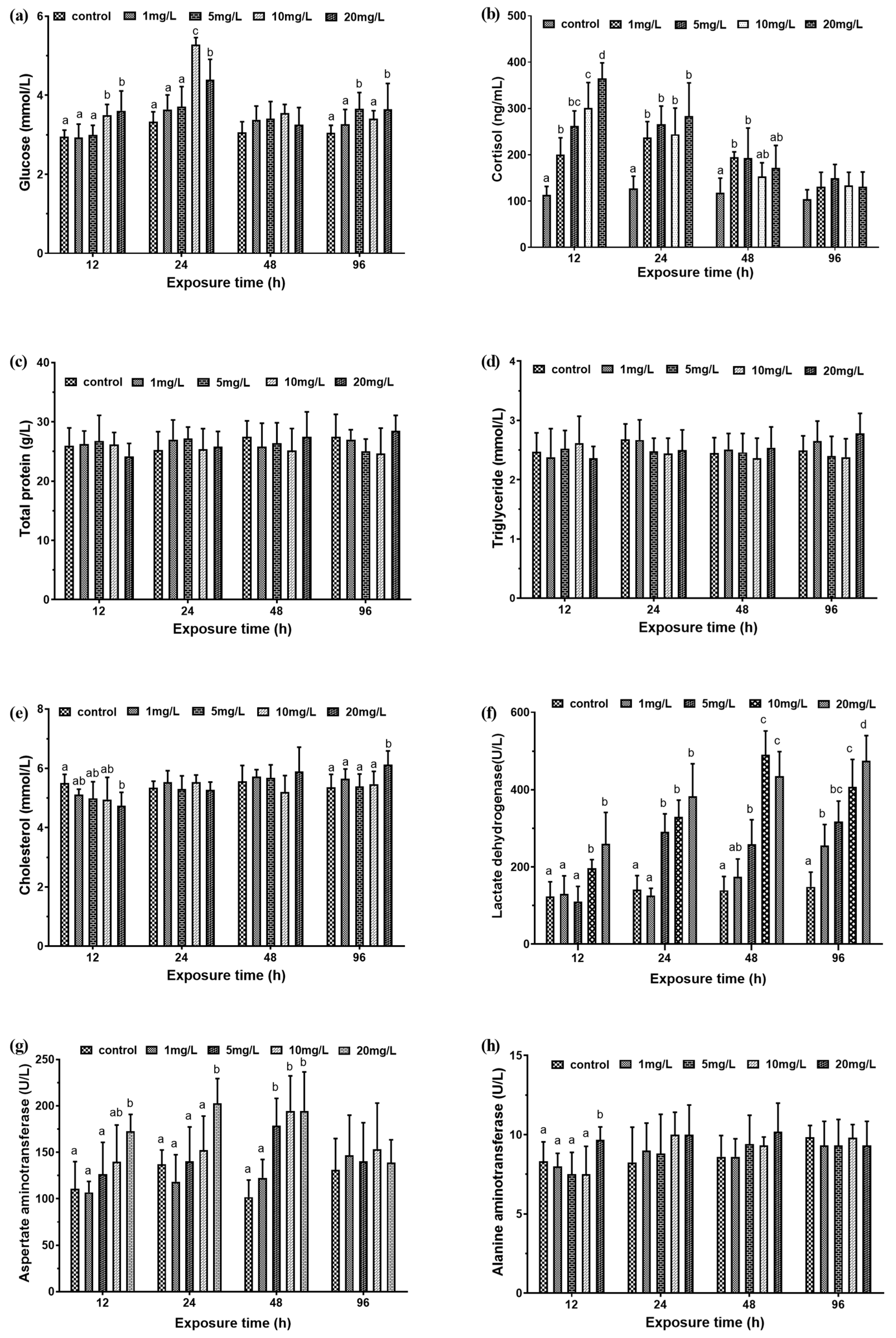
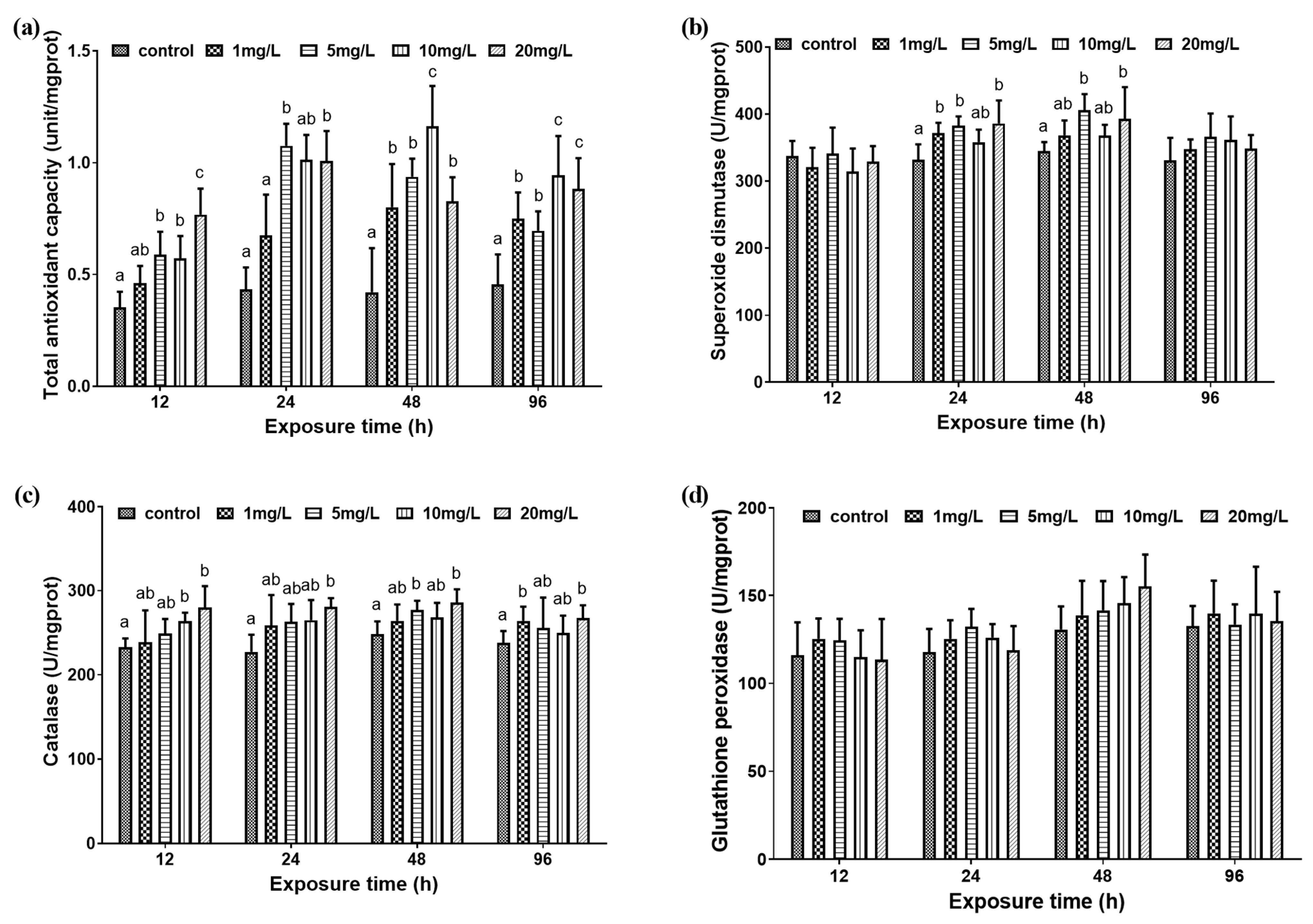
3.1. Biochemical Indicators and Antioxidant Capacity
3.1.1. Serum Biochemical Index
3.1.2. Liver Antioxidant Capacity
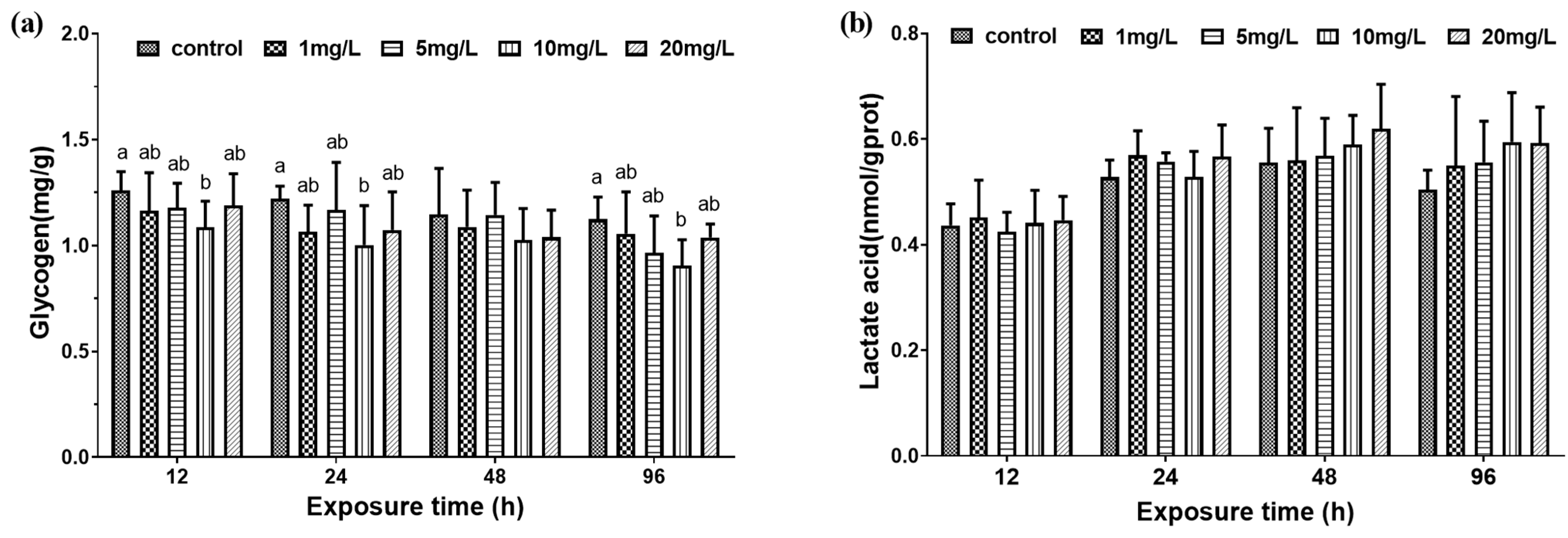
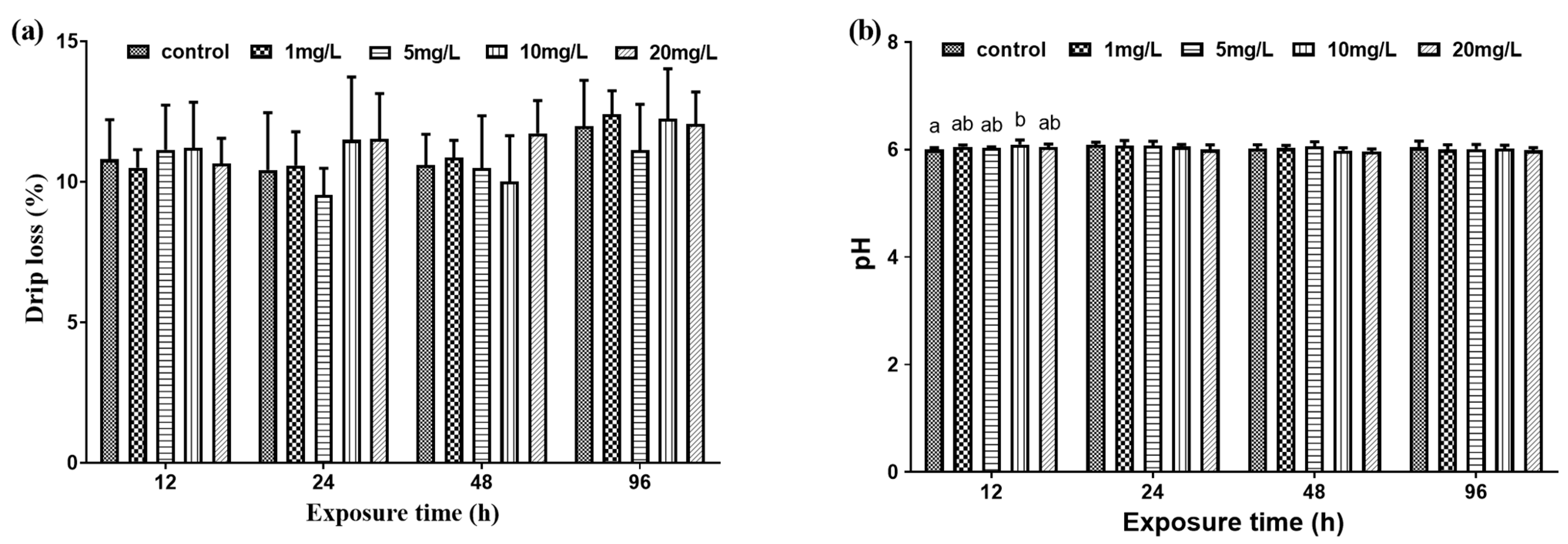
3.2. Muscle Metabolism
3.2.1. Muscle Physiological Characteristics
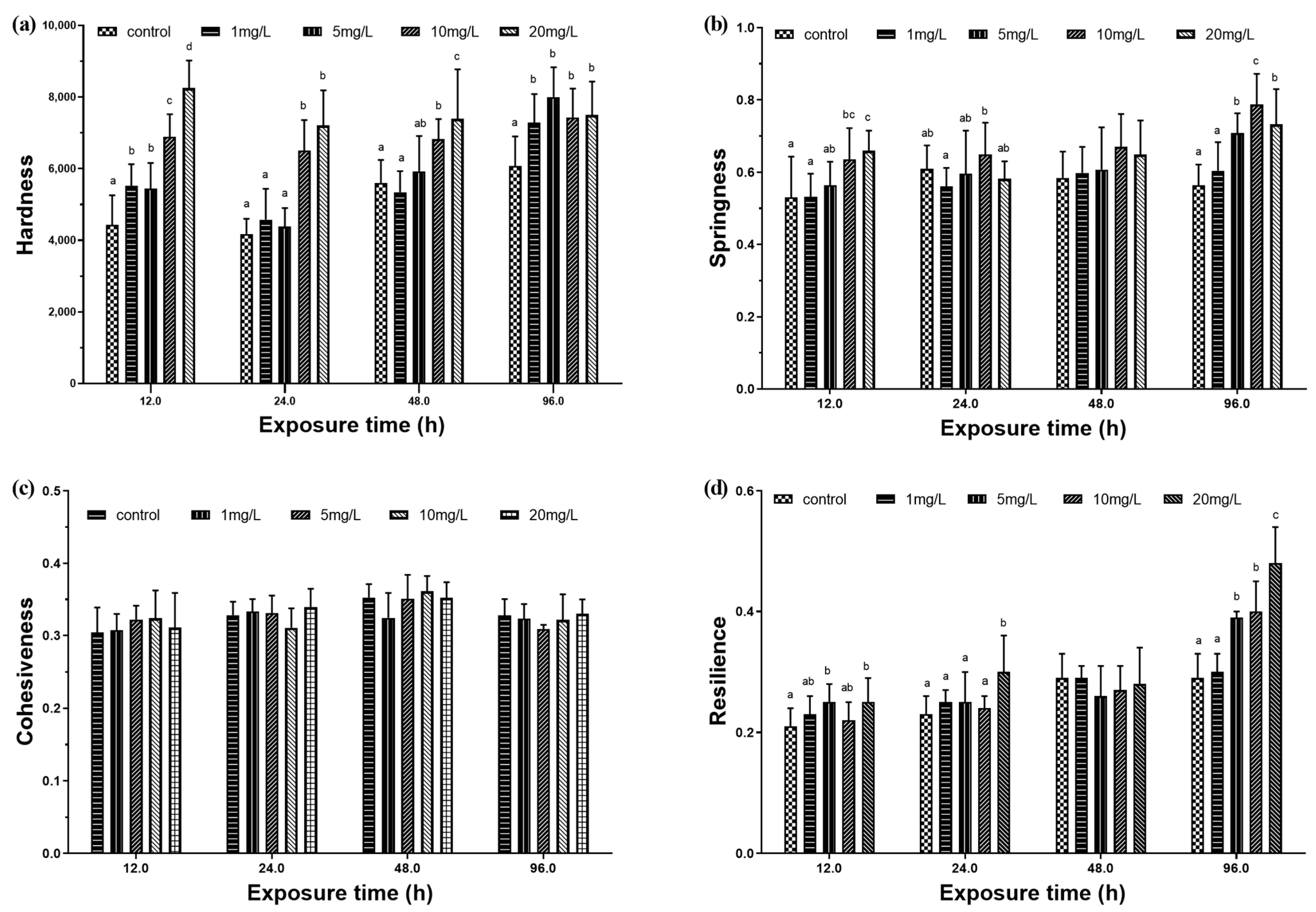
3.2.2. Muscle Physical Characteristics
4. Discussion
4.1. The Effect of Nitrite on Serum Biochemical Indices
4.2. The Effect of Nitrite on Liver Antioxidant Capacity
4.3. The Effect of Nitrite on Muscle Quality
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Das, P.; Ayyappan, S.; Das, B.; Jena, J. Nitrite toxicity in Indian major carps: Sublethal effect on selected enzymes in fingerlings of Catla catla, Labeo rohita and Cirrhinus mrigala. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2004, 138, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Avilez, I.M.; de Aguiar, L.H.; Hori, T.S.; Moraes, G. Metabolic responses of matrinxã, Brycon Amazonicus (Spix & Agassiz, 1829), exposed to environmental nitrite. Aquac. Res. 2012, 44, 596–603. [Google Scholar] [CrossRef]
- Kim, J.-H.; Kang, J.-C. The lead accumulation and hematological findings in juvenile rock fish Sebastes Schlegelii exposed to the dietary lead (II) concentrations. Ecotoxicol. Environ. Saf. 2015, 115, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-S.; Ha, T.-Y.; Kim, S.; Lee, S.-J.; Ahn, J. Red paprika (Capsicum annuum L.) and its main carotenoid capsanthin ameliorate impaired lipid metabolism in the liver and adipose tissue of high-fat diet-induced obese mice. J. Funct. Foods 2017, 31, 131–140. [Google Scholar] [CrossRef]
- Jensen, F.B. Nitrite disrupts multiple physiological functions in aquatic animals. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2003, 135, 9–24. [Google Scholar] [CrossRef]
- Xian, J.-A.; Wang, A.-L.; Chen, X.-D.; Gou, N.-N.; Miao, Y.-T.; Liao, S.-A.; Ye, C.-X. Cytotoxicity of nitrite on haemocytes of the tiger shrimp, Penaeus Monodon, using flow cytometric analysis. Aquaculture 2011, 317, 240–244. [Google Scholar] [CrossRef]
- Wang, J.; Tang, H.; Zhang, X.; Xue, X.; Zhu, X.; Chen, Y.; Yang, Z. Mitigation of nitrite toxicity by increased salinity is associated with multiple physiological responses: A case study using an economically important model species, the juvenile obscure puffer (Takifugu obscurus). Environ. Pollut. 2018, 232, 137–145. [Google Scholar] [CrossRef]
- Chen, W.; Liu, H.; Zhang, Q.; Dai, S. Effect of nitrite on growth and microcystins production of Microcystis Aeruginosa PCC7806. Environ. Boil. Fishes 2010, 23, 665–671. [Google Scholar] [CrossRef]
- Guo, H.; Xian, J.-A.; Wang, A.-L. Analysis of digital gene expression profiling in hemocytes of white shrimp Litopenaeus Vannamei under nitrite stress. Fish Shellfish. Immunol. 2016, 56, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, P.K.; Jensen, F.B. Recovery from nitrite-induced methaemoglobinaemia and potassium balance disturbances in carp. Fish Physiol. Biochem. 1997, 16, 1–10. [Google Scholar] [CrossRef]
- Ciji, A.; Sahu, N.P.; Pal, A.K.; Dasgupta, S.; Akhtar, M.S. Alterations in serum electrolytes, antioxidative enzymes and haematological parameters of Labeo Rohita on short-term exposure to sublethal dose of nitrite. Fish Physiol. Biochem. 2012, 38, 1355–1365. [Google Scholar] [CrossRef]
- Lin, Y.; Miao, L.-H.; Pan, W.-J.; Huang, X.; Dengu, J.M.; Zhang, W.-X.; Ge, X.-P.; Liu, B.; Ren, M.-C.; Zhou, Q.-L.; et al. Effect of nitrite exposure on the antioxidant enzymes and glutathione system in the liver of bighead carp, Aristichthys nobilis. Fish Shellfish. Immunol. 2018, 76, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Das, P.C.; Ayyappan, S.; Jena, J.K.; Das, B.K. Effect of sub-lethal nitrite on selected haematological parameters in fingerling Catla Catla (Hamilton). Aquac. Res. 2004, 35, 874–880. [Google Scholar] [CrossRef]
- da Costa, O.T.F.; Ferreira, D.J.D.S.; Mendonça, F.L.P.; Fernandes, M.N. Susceptibility of the Amazonian fish, Colossoma Macropomum (Serrasalminae), to short-term exposure to nitrite. Aquaculture 2004, 232, 627–636. [Google Scholar] [CrossRef]
- Kim, J.-H.; Kim, J.-Y.; Lim, L.-J.; Kim, S.K.; Choi, H.S.; Hur, Y.B. Effects of waterborne nitrite on hematological parameters and stress indicators in olive flounders, Paralichthys Olivaceus, raised in bio-floc and seawater. Chemosphere 2018, 209, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Grosell, M.; Jensen, F. Uptake and effects of nitrite in the marine teleost fish Platichthys flesus. Aquat. Toxicol. 2000, 50, 97–107. [Google Scholar] [CrossRef]
- Li, M.; Yu, N.; Qin, J.; Li, E.; Du, Z.; Chen, L. Effects of ammonia stress, dietary linseed oil and Edwardsiella Ictaluri challenge on juvenile darkbarbel catfish Pelteobagrus vachelli. Fish Shellfish. Immunol. 2014, 38, 158–165. [Google Scholar] [CrossRef]
- Xie, L.; Chen, S.; Yao, C.; Li, D.; Li, L.; Tang, R. Nitrite induces endoplasmic reticulum stress and associates apoptosis of liver cells in grass carp (Ctenopharyngodon Idella). Aquaculture 2019, 507, 275–281. [Google Scholar] [CrossRef]
- Kim, J.-H.; Kang, J.-C. The selenium accumulation and its effect on growth, and haematological parameters in red sea bream, Pagrus Major, exposed to waterborne selenium. Ecotoxicol. Environ. Saf. 2014, 104, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Li, J.; Tang, L.; Yang, Z. Responses of crucian carp Carassius Auratus to long-term exposure to nitrite and low dissolved oxygen levels. Biochem. Syst. Ecol. 2012, 44, 224–232. [Google Scholar] [CrossRef]
- Zhou, X.-X.; Wang, Y.-B.; Li, W.-F. Effect of feeding apidaecin on common carp (Cyprinus Carpio) growth performances and immune function. Aquaculture 2008, 279, 108–112. [Google Scholar] [CrossRef]
- Martinez, C.B.; Souza, M.M. Acute effects of nitrite on ion regulation in two neotropical fish species. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 133, 151–160. [Google Scholar] [CrossRef]
- Das, P.; Ayyappan, S.; Jena, J.; Das, B. Nitrite toxicity in Cirrhinus Mrigala (Ham.): Acute toxicity and sub-lethal effect on selected haematological parameters. Aquaculture 2004, 235, 633–644. [Google Scholar] [CrossRef]
- Hong, M.; Chen, L.; Sun, X.; Gu, S.; Zhang, L.; Chen, Y. Metabolic and immune responses in Chinese mitten-handed crab (Eriocheir Sinensis) juveniles exposed to elevated ambient ammonia. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 145, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shen, Z.; Qi, T.; Xi, R.; Liang, X.; Li, L.; Tang, R.; Li, D. Slight increases in salinity improve muscle quality of grass carp (Ctenopharyngodon idellus). Fishes 2021, 6, 7. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Tang, R.; He, X.; Li, L.; Takagi, Y.; Li, D. Improvement of muscle quality of grass carp (Ctenopharyngodon Idellus) with a bio-floating bed in culture ponds. Front. Physiol. 2019, 10, 683. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Lin, W.; Wang, L.; Zhang, D.; Wu, X.; Li, L.; Li, D.; Tang, R.; Yang, L.; Qiu, Y. The supplementation of dietary selenium yeast and green tea-derived polyphenols improves antioxidant capacity and immune response in juvenile Wuchang bream under ammonia stress. Aquac. Res. 2020, 51, 3790–3803. [Google Scholar] [CrossRef]
- Mommsen, T.P.; Vijayan, M.M.; Moon, T.W. Cortisol in teleosts: Dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol. Fish. 1999, 9, 211–268. [Google Scholar] [CrossRef]
- Montero, D.; Izquierdo, M.; Tort, L.; Robaina, L.; Vergara, J. High stocking density produces crowding stress altering some physiological and biochemical parameters in gilthead seabream, Sparus Aurata, juveniles. Fish Physiol. Biochem. 1999, 20, 53–60. [Google Scholar] [CrossRef]
- Kumaraguru, A.; Beamish, F. Effect of permethrin (NRDC-143) on the bioenergetics of rainbow trout, Salmo gairdneri. Aquat. Toxicol. 1986, 9, 47–58. [Google Scholar] [CrossRef]
- Cheng, S.-Y.; Shieh, L.-W.; Chen, J.-C. Changes in hemolymph oxyhemocyanin, acid–base balance, and electrolytes in Marsupenaeus japonicus under combined ammonia and nitrite stress. Aquat. Toxicol. 2013, 130, 132–138. [Google Scholar] [CrossRef]
- Chen, J.-C.; Cheng, S.-Y. Hemolymph PCO2, hemocyanin, protein levels and urea excretions of Penaeus Monodon exposed to ambient ammonia. Aquat. Toxicol. 1993, 27, 281–291. [Google Scholar] [CrossRef]
- Ciji, A.; Akhtar, M.S. Nitrite implications and its management strategies in aquaculture: A review. Rev. Aquac. 2019, 12, 878–908. [Google Scholar] [CrossRef]
- Olsen, Y.A.; Einarsdottir, I.E.; Nilssen, K.J. Metomidate anaesthesia in Atlantic salmon, Salmo Salar, prevents plasma cortisol increase during stress. Aquaculture 1995, 134, 155–168. [Google Scholar] [CrossRef]
- Das, P.C.; Ayyappan, S.; Jena, J.K.; Das, B.K. Acute toxicity of ammonia and its sub-lethal effects on selected haematological and enzymatic parameters of mrigal, Cirrhinus Mrigala (Hamilton). Aquac. Res. 2004, 35, 134–143. [Google Scholar] [CrossRef]
- Bogin, E.; Marom, M.; Levi, Y. Changes in serum, liver and kidneys of cisplatin-treated rats; effects of antioxidants. Clin. Chem. Lab. Med. 1994, 32, 843–852. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Ge, X.; Zhu, J.; Xuan, F.; Jiang, X. Identification and mRNA expression of antioxidant enzyme genes associated with the oxidative stress response in the Wuchang bream (Megalobrama Amblycephala Yih) in response to acute nitrite exposure. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 159, 69–77. [Google Scholar] [CrossRef]
- Zheng, J.; Mao, Y.; Su, Y.; Wang, J. Effects of nitrite stress on mRNA expression of antioxidant enzymes, immune-related genes and apoptosis-related proteins in Marsupenaeus japonicus. Fish Shellfish. Immunol. 2016, 58, 239–252. [Google Scholar] [CrossRef]
- Parihar, M.S.; Dubey, A.K.; Prakash, P. Responses of superoxide dismutase, glutathione peroxidase and reduced gluta-thione antioxidant defenses in gills of the freshwater catfish (Heteropneustes Fossills) to short-term elevated temperature. J. Therm. Biol. 1997, 22, 151–156. [Google Scholar] [CrossRef]
- Wei, W.; Cuijuan, N.; Ying, G.; Li, L. The effects of environmental endocrine disruptors on fertilization and embryonic development of the fish Liza haematocheila. Comp. Biochem. Physiol. Part C 2008, 4, 466. [Google Scholar] [CrossRef]
- Chatterjee, N.; Pal, A.; Manush, S.; Das, T.; Mukherjee, S. Thermal tolerance and oxygen consumption of Labeo Rohita and Cyprinus Carpio early fingerlings acclimated to three different temperatures. J. Therm. Biol. 2004, 29, 265–270. [Google Scholar] [CrossRef]
- Wang, W.-N.; Wang, A.-L.; Zhang, Y.-J.; Li, Z.-H.; Wang, J.-X.; Sun, R.-Y. Effects of nitrite on lethal and immune response of Macrobrachium nipponense. Aquaculture 2004, 232, 679–686. [Google Scholar] [CrossRef]
- de Lima, R.L.; Braun, N.; Kochhann, D.; Lazzari, R.; Neto, J.R.; Moraes, B.S.; Loro, V.L.; Baldisserotto, B. Survival, growth and metabolic parameters of silver catfish, Rhamdia Quelen, juveniles exposed to different waterborne nitrite levels. Neotrop. Ichthyol. 2011, 9, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Hemre, G.-I.; Mommsen, T.; Krogdahl, Å. Carbohydrates in fish nutrition: Effects on growth, glucose metabolism and hepatic enzymes. Aquac. Nutr. 2002, 8, 175–194. [Google Scholar] [CrossRef]
- Neyrão, I.M.; Biller, J.D.; Takahashi, L.S.; Urbinati, E.C. Modulation of immunity and hepatic antioxidant defense by corticosteroids in pacu (Piaractus Mesopotamicus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2021, 260, 111025. [Google Scholar] [CrossRef] [PubMed]
- Johnston, A.I. Muscle development and growth: Potential implications for flesh quality in fish. Aquaculture 1999, 177, 99–115. [Google Scholar] [CrossRef]
- He, H.-J.; Wu, D.; Sun, D.-W. Rapid and non-destructive determination of drip loss and pH distribution in farmed Atlantic salmon (Salmo Salar) fillets using visible and near-infrared (Vis–NIR) hyperspectral imaging. Food Chem. 2014, 156, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Luciano, G.; Monahan, F.; Vasta, V.; Biondi, L.; Lanza, M.; Priolo, A. Dietary tannins improve lamb meat colour stability. Meat Sci. 2009, 81, 120–125. [Google Scholar] [CrossRef]
- Ofstad, R.; Egelandsdal, B.; Kidman, S.; Myklebust, R.; Olsen, R.L.; Hermansson, A.-M. liquid loss as effected by post mortem ultrastructural changes in fish muscle: Cod (Gadus Morhua L) and salmon (Salmo Salar). J. Sci. Food Agric. 1996, 71, 301–312. [Google Scholar] [CrossRef]
- Wang, P.A.; Vang, B.; Pedersen, A.M.; Martinez, I.; Olsen, R.L. Post-Mortem degradation of myosin heavy chain in intact fish muscle: Effects of pH and enzyme inhibitors. Food Chem. 2011, 124, 1090–1095. [Google Scholar] [CrossRef] [Green Version]
- Ando, M.; Nakamura, H.; Harada, R.; Yamane, A. Effect of super chilling storage on maintenance of freshness of Kuruma Prawn. Food Sci. Technol. Res. 2004, 10, 25–31. [Google Scholar] [CrossRef]
- Suárez, M.; Martínez, T.; Abellán, E.; Arizcun, M.; Jimenez, M.C.H.; Hidalgo, M.; Cardenete, G. The effects of the diet on flesh quality of farmed dentex (Dentex Dentex). Aquaculture 2009, 288, 106–113. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Z.; Qi, C.; Lin, C.; Tang, R. Nitrite Stress Induces Oxidative Stress and Leads to Muscle Quality Decreased in Wuchang Bream (Megalobrama amblycephala Yih) Juveniles. Water 2022, 14, 160. https://doi.org/10.3390/w14020160
Hu Z, Qi C, Lin C, Tang R. Nitrite Stress Induces Oxidative Stress and Leads to Muscle Quality Decreased in Wuchang Bream (Megalobrama amblycephala Yih) Juveniles. Water. 2022; 14(2):160. https://doi.org/10.3390/w14020160
Chicago/Turabian StyleHu, Zhenyi, Chenglong Qi, Chenzhi Lin, and Rong Tang. 2022. "Nitrite Stress Induces Oxidative Stress and Leads to Muscle Quality Decreased in Wuchang Bream (Megalobrama amblycephala Yih) Juveniles" Water 14, no. 2: 160. https://doi.org/10.3390/w14020160
APA StyleHu, Z., Qi, C., Lin, C., & Tang, R. (2022). Nitrite Stress Induces Oxidative Stress and Leads to Muscle Quality Decreased in Wuchang Bream (Megalobrama amblycephala Yih) Juveniles. Water, 14(2), 160. https://doi.org/10.3390/w14020160