
Lotic Environment Affects Morphological Characteristics and Energy Metabolism of Juvenile Grass Carp Ctenopharyngodon idella



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Experimental Conditions
2.2. Experimental Protocol
2.3. Sampling
2.4. Analytical Procedures
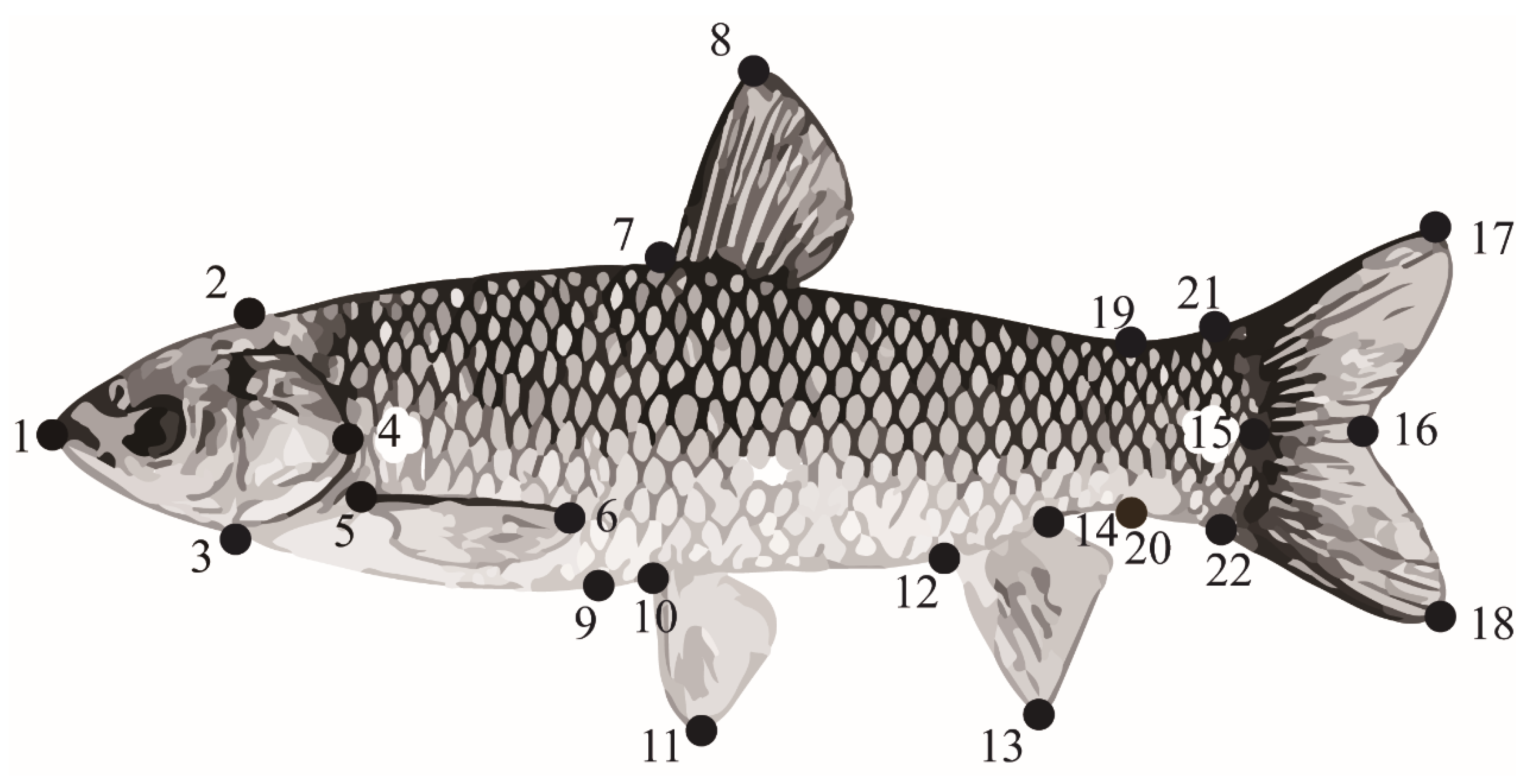
2.4.1. Measurement of Morphology
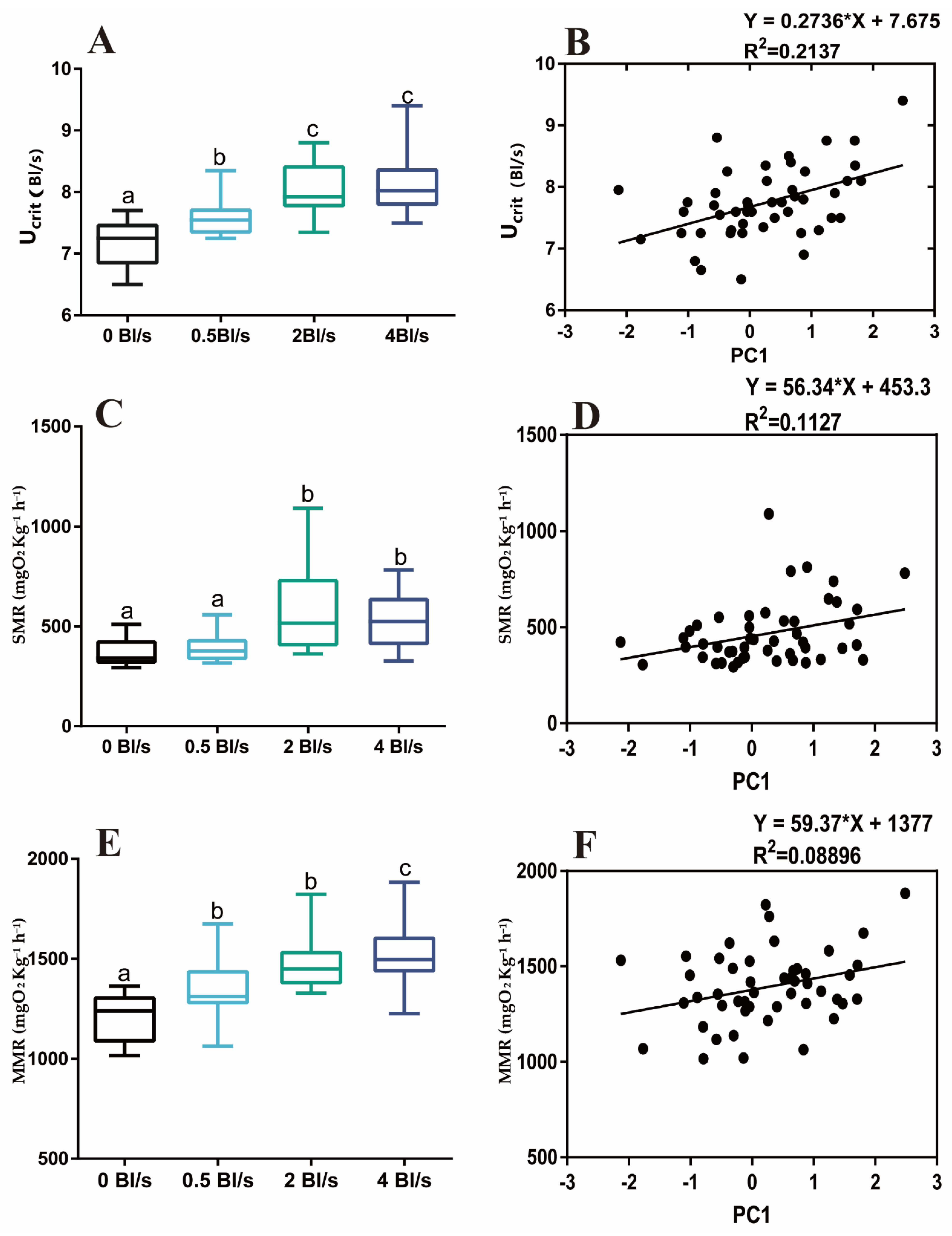
2.4.2. Measurement of Aerobic Swimming Ability
2.4.3. Measurements of Oxygen Consumption Rate
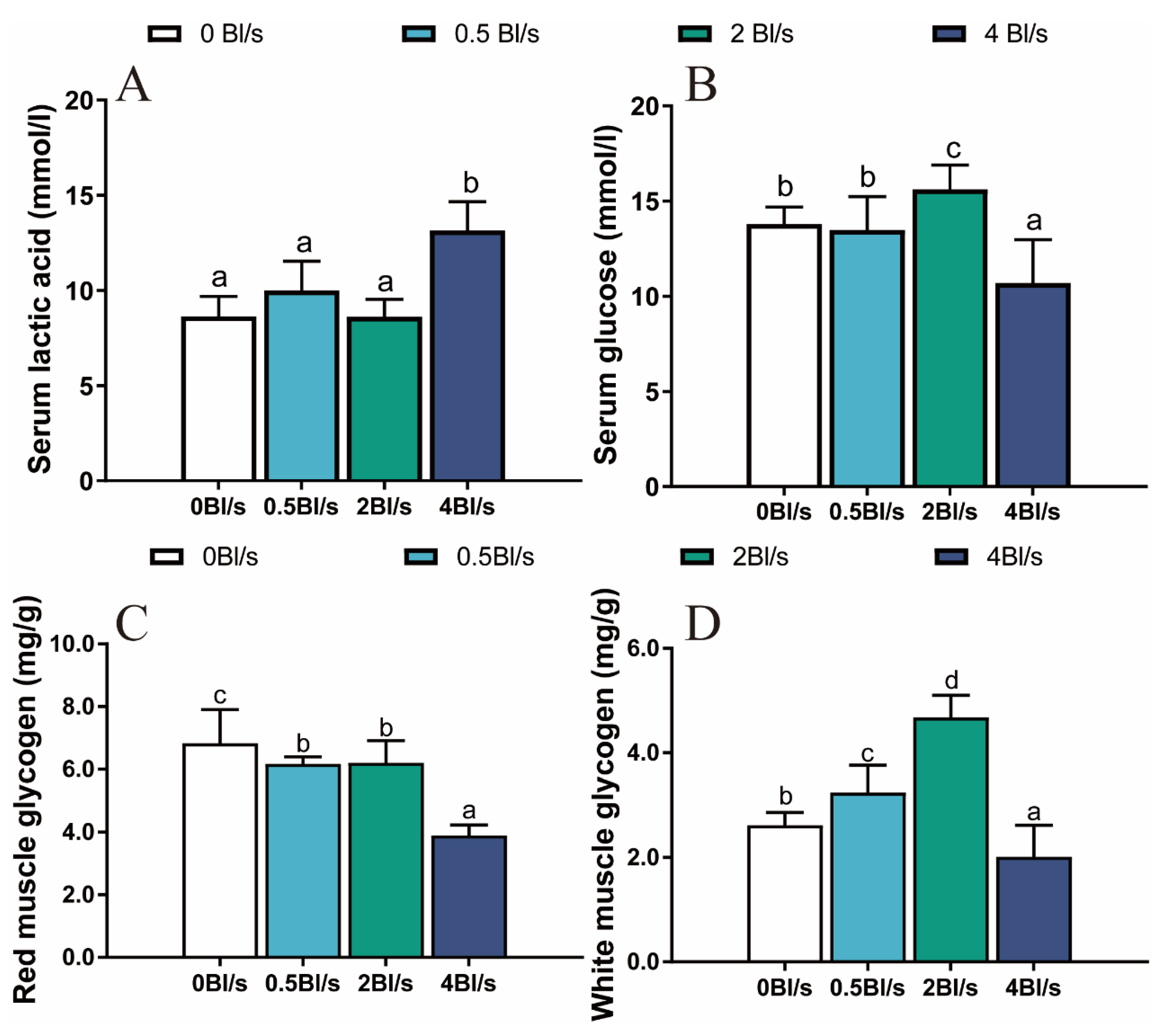
2.4.4. Determination of Contents of Lactic Acid and Glucose in Serum
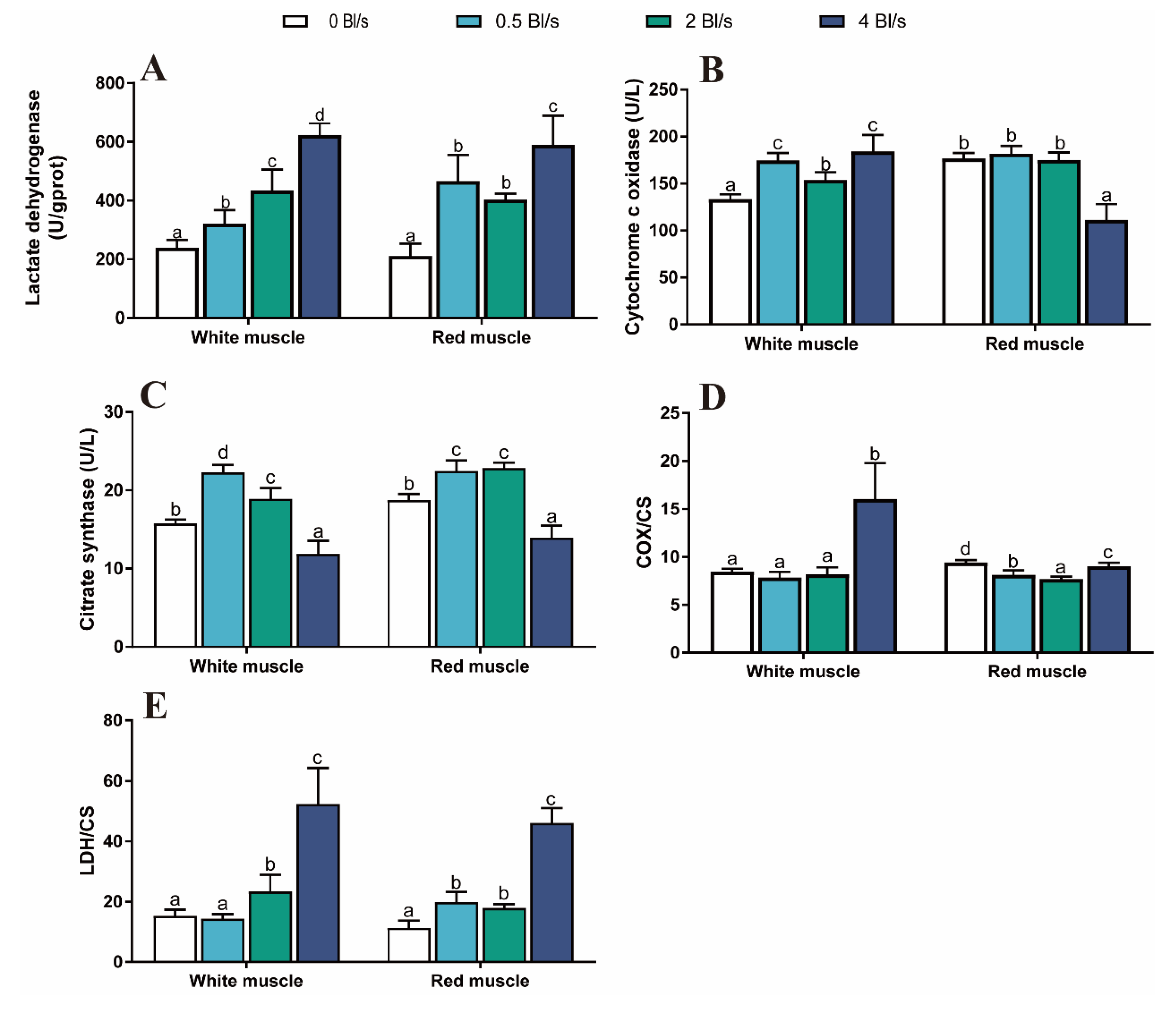
2.4.5. Determination of Glycogen and Enzyme Activity of Muscle
2.4.6. Gene Expression Level of AMPK Pathway in Red and White Muscle
2.5. Statistics
3. Results
3.1. Growth Performance
3.2. Morphological Characteristics, Aerobic Swimming Performance, and Oxygen Consumption Rate
3.3. Serum Biochemical Parameters and Muscle Glycogen Content
3.4. Muscle Enzyme Activity
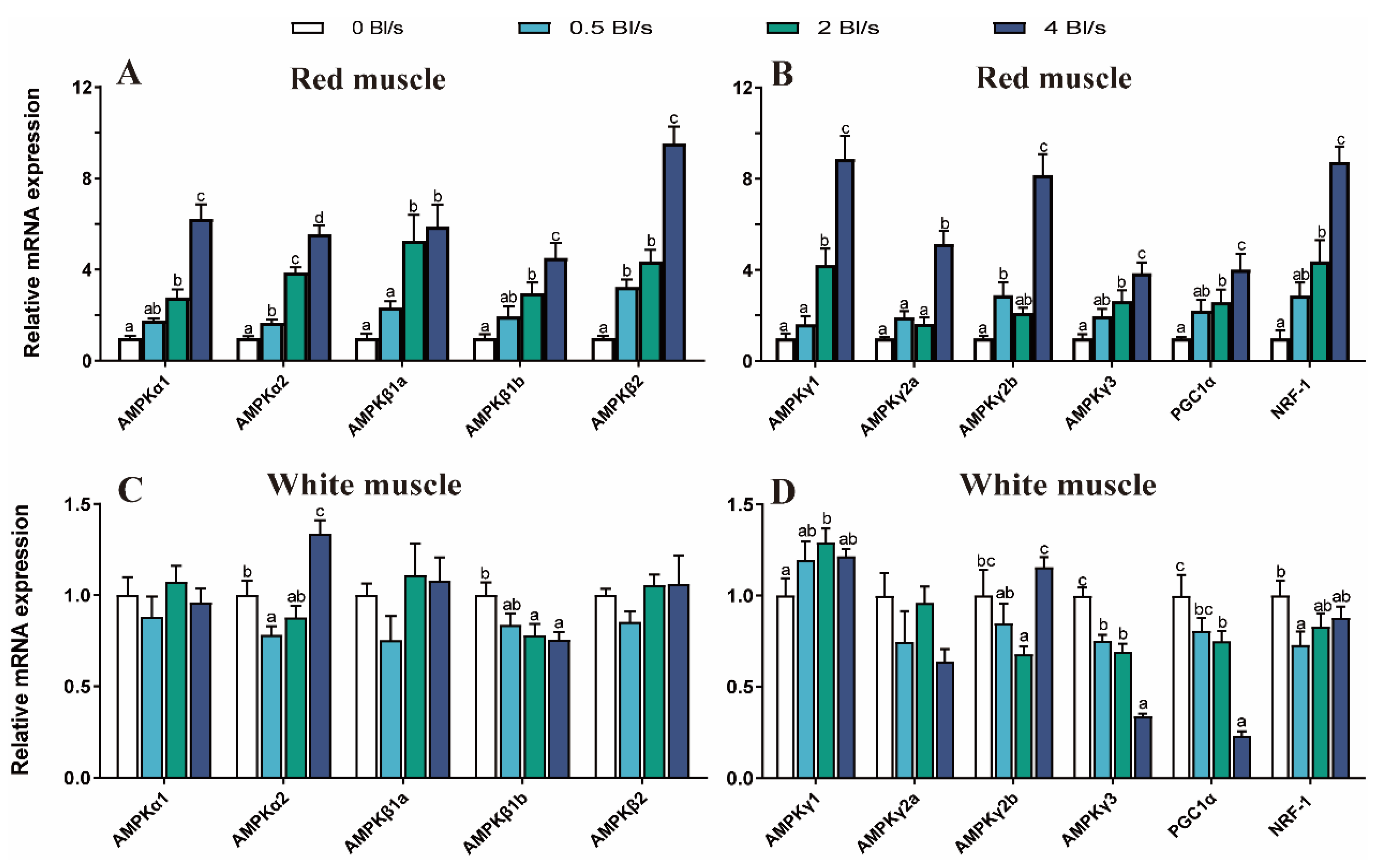
3.5. AMPK-PGC-1α Signaling Pathway Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix A.1. Differences in Morphological Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | Water Velocity | |||
|---|---|---|---|---|
| 0 Bl/s | 0.5 Bl/s | 2 Bl/s | 4 Bl/s | |
| 1–2/body length | 23.82 ± 1.50 | 23.69 ± 1.11 | 23.38 ± 1.13 | 23.90 ± 1.16 |
| 1–3/body length | 20.58 ± 0.97 | 19.95 ± 1.48 | 20.36 ± 1.21 | 19.90 ± 1.14 |
| 1–4/body length | 25.47 ± 1.40 | 24.90 ± 1.09 | 25.07 ± 0.99 | 25.13 ± 1.12 |
| 1–16/body length | 109.35 ± 1.09 b | 108.53 ± 0.88 ab | 108.46 ± 1.43 a | 107.75 ± 1.57 a |
| 1–17/body length | 117.89 ± 1.59 | 117.49 ± 1.50 | 116.91 ± 1.76 | 117.15 ± 2.42 |
| 1–18/body length | 120.01 ± 1.79 b | 119.77 ± 1.54 b | 118.91 ± 1.22 ab | 117.90 ± 2.30 a |
| 2–3/body length | 18.94 ± 0.68 a | 19.33 ± 1.02 a | 19.10 ± 0.93 a | 20.06 ± 0.90 b |
| 2–7/body length | 32.62 ± 2.49 | 32.60 ± 1.38 | 32.75 ± 1.85 | 32.31 ± 1.92 |
| 5–6/body length | 15.32 ± 1.84 a | 16.21 ± 1.08 ab | 16.98 ± 1.49 b c | 17.81 ± 1.00 c |
| 7–8/body length | 20.93 ± 1.94 ab | 19.53 ± 2.24 a | 20.62 ± 2.18 ab | 21.54 ± 1.89 b |
| 7–9/body length | 23.22 ± 0.87 a | 24.45 ± 1.52 b | 24.72 ± 1.44 b | 26.52 ± 2.16 c |
| 10–11/body length | 10.92 ± 1.15 a | 11.09 ± 1.37 a | 11.72 ± 1.29 ab | 12.44 ± 1.28 b |
| 12–13/body length | 15.94 ± 2.40 b | 14.21 ± 1.60 a | 14.75 ± 1.73 ab | 15.18 ± 1.65 ab |
| 14–22/body length | 12.48 ± 1.59 | 11.36 ± 2.01 | 12.03 ± 1.54 | 12.12 ± 1.48 |
| 17–18/body length | 17.30 ± 4.22 a | 20.48 ± 3.48 b | 18.56 ± 5.36 ab | 19.73 ± 3.81 ab |
| 17–21/body length | 21.49 ± 1.64 | 22.15 ± 1.37 | 22.07 ± 1.39 | 22.13 ± 1.98 |
| 18–22/body length | 23.25 ± 1.42 ab | 23.47 ± 1.23 b | 22.93 ± 1.30 ab | 22.48 ± 1.48 a |
| 19–20/body length | 10.25 ± 0.55 a | 11.20 ± 0.81 b | 11.00 ± 0.64 b | 11.40 ± 0.96 b |
| 19–21/body length | 32.87 ± 2.40 a | 33.56 ± 2.19 a | 32.28 ± 2.65 a | 35.34 ± 1.75 b |
Appendix A.2. Principal Component Analysis
| Morphometric Characteristics | Principal Component 1 | Principal Component 2 | Principal Component 3 | Principal Component 4 | Principal Component 5 | Principal Component 6 |
|---|---|---|---|---|---|---|
| 1–2/body length % | 0.126 | −0.066 | 0.835 | −0.023 | 0.196 | −0.077 |
| 1–3/body length % | −0.229 | −0.077 | 0.714 | 0.011 | −0.197 | −0.042 |
| 1–4/body length % | −0.035 | −0.130 | 0.904 | 0.101 | 0.067 | 0.093 |
| 1–16/body length % | −0.178 | 0.270 | 0.007 | 0.724 | −0.091 | −0.004 |
| 1–17/body length % | −0.055 | 0.220 | 0.035 | 0.755 | 0.024 | −0.046 |
| 1–18/body length % | −0.179 | 0.774 | 0.017 | 0.367 | −0.007 | −0.144 |
| 2–3/body length % | 0.616 | −0.070 | 0.323 | −0.364 | 0.022 | 0.045 |
| 2–7/body length % | 0.035 | −0.032 | −0.088 | 0.424 | −0.003 | −0.778 |
| 5–6/body length % | 0.832 | −0.026 | −0.051 | 0.177 | −0.043 | 0.018 |
| 7–8/body length % | 0.108 | 0.075 | −0.091 | 0.279 | 0.182 | 0.757 |
| 7–9/body length % | 0.843 | −0.119 | −0.054 | −0.214 | 0.095 | −0.007 |
| 10–11/body length % | 0.334 | 0.002 | −0.057 | 0.236 | 0.587 | 0.070 |
| 12–13/body length % | −0.425 | 0.244 | 0.111 | −0.147 | 0.580 | −0.117 |
| 14–22/body length % | −0.323 | −0.013 | −0.068 | 0.026 | 0.543 | 0.310 |
| 17–18/body length % | 0.252 | 0.456 | −0.247 | −0.508 | 0.028 | −0.257 |
| 17–21/body length % | 0.014 | 0.821 | −0.146 | 0.058 | 0.153 | 0.104 |
| 18–22/body length % | −0.065 | 0.851 | −0.123 | 0.162 | −0.155 | 0.139 |
| 19–20/body length % | 0.747 | 0.058 | −0.196 | −0.334 | 0.088 | −0.006 |
| 19–21/body length % | 0.164 | −0.078 | 0.093 | −0.151 | 0.651 | 0.040 |
| Characteristic value | 2.981 | 2.449 | 2.318 | 2.208 | 1.591 | 1.433 |
| Contribution rate | 15.687545 | 12.88992 | 12.19804 | 11.61951 | 8.373102 | 7.541147 |
| Cumulative Contribution rate | 15.687545 | 28.57747 | 40.7755 | 52.39501 | 60.76812 | 68.30926 |
Appendix A.3. Aerobic Swimming Performance and Morphological Characteristics
| Parameter | Principal Component | R2 | p | Df | F |
|---|---|---|---|---|---|
| Ucrit | PC1 | 0.2137 | 0.0009 | 1.000, 46.00 | 12.5 |
| PC2 | 0.002314 | 0.7454 | 1.000, 46.00 | 0.1067 | |
| PC3 | 0.00704 | 0.5707 | 1.000, 46.00 | 0.3261 | |
| PC4 | 0.05543 | 0.1072 | 1.000, 46.00 | 2.7 | |
| PC5 | 0.06287 | 0.0856 | 1.000, 46.00 | 3.086 | |
| PC6 | 0.02617 | 0.2720 | 1.000, 46.0 | 1.236 |
References
- Li, X.; Zhang, Y.; Li, X.; Zheng, H.; Peng, J.; Fu, S. Sustained Exercise-Trained Juvenile Black Carp (Mylopharyngodon piceus) at a Moderate Water Velocity Exhibit Improved Aerobic Swimming Performance and Increased Postprandial Metabolic Responses. Biol. Open 2018, 7, bio032425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yogata, H.; Oku, H. The Effects of Swimming Exercise on Growth and Whole-Body Protein and Fat Contents of Fed and Unfed Fingerling Yellowtail. Fish. Sci. 2000, 66, 1100–1105. [Google Scholar] [CrossRef] [Green Version]
- Gallaugher, P.E.; Thorarensen, H.; Kiessling, A.; Farrell, A.P. Effects of High Intensity Exercise Training on Cardiovascular Function, Oxygen Uptake, Internal Oxygen Transport and Osmotic Balance in Chinook Salmon (Oncorhynchus tshawytscha) during Critical Speed Swimming. J. Exp. Biol. 2001, 204, 2861–2872. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.I.; Johnston, I.A. The Role of Myostatin and the Calcineurin-Signalling Pathway in Regulating Muscle Mass in Response to Exercise Training in the Rainbow Trout Oncorhynchus mykiss Walbaum. J. Exp. Biol. 2005, 208, 2083–2090. [Google Scholar] [CrossRef] [Green Version]
- Davison, W. The Effects of Exercise Training on Teleost Fish, a Review of Recent Literature. Comp. Biochem. Physiol. Part A Physiol. 1997, 117, 67–75. [Google Scholar] [CrossRef]
- Plaut, I. Critical Swimming Speed: Its Ecological Relevance. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 131, 41–50. [Google Scholar] [CrossRef]
- Ohlberger, J.; Staaks, G.; Hölker, F. Swimming Efficiency and the Influence of Morphology on Swimming Costs in Fishes. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2006, 176, 17–25. [Google Scholar] [CrossRef]
- Binning, S.A.; Roche, D.G.; Fulton, C.J. Localised Intraspecific Variation in the Swimming Phenotype of a Coral Reef Fish across Different Wave Exposures. Oecologia 2014, 174, 623–630. [Google Scholar] [CrossRef]
- Nelson, J.A.; Atzori, F.; Gastrich, K.R. Repeatability and Phenotypic Plasticity of Fish Swimming Performance across a Gradient of Urbanization. Environ. Biol. Fishes 2015, 98, 1431–1447. [Google Scholar] [CrossRef]
- Killen, S.S.; Atkinson, D.; Glazier, D.S. The Intraspecific Scaling of Metabolic Rate with Body Mass in Fishes Depends on Lifestyle and Temperature. Ecol. Lett. 2010, 13, 184–193. [Google Scholar] [CrossRef]
- Auer, S.K.; Salin, K.; Rudolf, A.M.; Anderson, G.J.; Metcalfe, N.B. The Optimal Combination of Standard Metabolic Rate and Aerobic Scope for Somatic Growth Depends on Food Availability. Funct. Ecol. 2015, 29, 479–486. [Google Scholar] [CrossRef]
- Castro, V.; Grisdale-Helland, B.; Jørgensen, S.M.; Helgerud, J.; Claireaux, G.; Farrell, A.P.; Krasnov, A.; Helland, S.J.; Takle, H. Disease Resistance Is Related to Inherent Swimming Performance in Atlantic Salmon. BMC Physiol. 2013, 13, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, J.R.; Trembath, C.; Pether, S.; Bruce, M.; Walker, S.P.; Herbert, N.A. Accommodating the Cost of Growth and Swim Ming in Fish—The Applicability of Exercise-Induced Growth to Juvenile Hapuku (Polyprion oxygeneios). Front. Physiol. 2014, 5, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaki, S.S.; Mabuchi, K.; Takamura, N.; Nishida, M.; Washitani, I. Behavioural and Morphological Differences between Feral and Domesticated Strains of Common Carp Cyprinus carpio. J. Fish Biol. 2009, 75, 1206–1220. [Google Scholar] [CrossRef]
- Killen, S.S.; Marras, S.; Metcalfe, N.B.; McKenzie, D.J.; Domenici, P. Environmental Stressors Alter Relationships between Physiology and Behaviour. Trends Ecol. Evol. 2013, 28, 651–658. [Google Scholar] [CrossRef]
- Rosenfeld, J.; Richards, J.; Allen, D.; Van Leeuwen, T.; Monnet, G. Adaptive Trade-Offs in Fish Energetics and Physiology: Insights from Adaptive Differentiation among Juvenile Salmonids. Can. J. Fish. Aquat. Sci. 2020, 77, 1243–1255. [Google Scholar] [CrossRef]
- Weber, J.M.; Choi, K.; Gonzalez, A.; Omlin, T. Metabolic Fuel Kinetics in Fish: Swimming, Hypoxia and Muscle Membranes. J. Exp. Biol. 2016, 219, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Ross, F.A.; MacKintosh, C.; Hardie, D.G. AMP-Activated Protein Kinase: A Cellular Energy Sensor That Comes in 12 Flavours. FEBS J. 2016, 283, 2987–3001. [Google Scholar] [CrossRef]
- Magnoni, L.J.; Vraskou, Y.; Palstra, A.P.; Planas, J.V. AMP-Activated Protein Kinase Plays an Important Evolutionary Conserved Role in the Regulation of Glucose Metabolism in Fish Skeletal Muscle Cells. PLoS ONE 2012, 7, e31219. [Google Scholar] [CrossRef] [Green Version]
- McClelland, G.B. Muscle Remodeling and the Exercise Physiology of Fish. Exerc. Sport Sci. Rev. 2012, 40, 165–173. [Google Scholar] [CrossRef]
- Jibb, L.A.; Richards, J.G. AMP-Activated Protein Kinase Activity during Metabolic Rate Depression in the Hypoxic Goldfish, Carassius Auratus. J. Exp. Biol. 2008, 211, 3111–3122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morash, A.J.; Vanderveken, M.; McClelland, G.B. Muscle Metabolic Remodeling in Response to Endurance Exercise in Salmonids. Front. Physiol. 2014, 5, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Nie, M.; Wang, L.; Xiong, Y.; Wang, F.; Wang, L.; Xiao, P.; Wu, Z.; Liu, Y.; You, F. Energy Response and Modulation of AMPK Pathway of the Olive Flounder Paralichthys Olivaceus in Low-Temperature Challenged. Aquaculture 2018, 484, 205–213. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, N.; Liu, X.; Onxayvieng, K.; Liu, L.; Tang, R.; Li, D. Exercise Training Accelerates UPS- and MTOR-Mediated Protein Turnover of Grass Carp Ctenopharyngodon idella. Aquaculture 2021, 545, 737252. [Google Scholar] [CrossRef]
- Lu, Y.; Wu, H.; Deng, L.-J.; Li, T.-C.; Yang, K.; Fu, S.-J.; Song, Z.-B. Improved Aerobic and Anaerobic Swimming Performance after Exercise Training and Detraining in Schizothorax Wangchiachii: Implications for Fisheries Releases. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 245, 110698. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Shao, F.; Ding, S.H.; Fu, S.J.; Zhang, Y.G. Interspecific Differences and Ecological Correlations of Energy Metabolism Traits in Freshwater Fishes. Funct. Ecol. 2020, 34, 616–630. [Google Scholar] [CrossRef]
- Roche, D.G.; Binning, S.A.; Bosiger, Y.; Johansen, J.L.; Rummer, J.L. Finding the Best Estimates of Metabolic Rates in a Coral Reef Fish. J. Exp. Biol. 2013, 216, 2103–2110. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.J.; Cao, Z.D.; Peng, J.L. Effect of Feeding and Fasting on Excess Post-Exercise Oxygen Consumption in Juvenile Southern Catfish (Silurus meridionalis Chen). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 146, 435–439. [Google Scholar] [CrossRef]
- Qin, N.; Li, D.; Hong, H.; Zhang, Y.; Zhu, B.; Luo, Y. Effects of Different Stunning Methods on the Flesh Quality of Grass Carp (Ctenopharyngodon idellus) Fillets Stored at 4 °C. Food Chem. 2016, 201, 131–138. [Google Scholar] [CrossRef]
- Zeng, K.W.; Liao, L.X.; Zhao, M.B.; Song, F.J.; Yu, Q.; Jiang, Y.; Tu, P.F. Protosappanin B Protects PC12 Cells against Oxygen-Glucose Deprivation-Induced Neuronal Death by Maintaining Mitochondrial Homeostasis via Induction of Ubiquitin-Dependent P53 Protein Degradation. Eur. J. Pharmacol. 2015, 751, 13–23. [Google Scholar] [CrossRef]
- Liang, X.; Fang, T.; Yan, F.; Xu, W.; Onxayvieng, K.; Tang, R.; Li, L.; Zhang, X.; Chi, W.; Li, D. Glutamate and Glutamine Transporter Genes in Grass Carp (Ctenopharyngodon idellus): Molecular Cloning, Characterization and Expression in Response to Different Stocking Densities. Aquac. Res. 2020, 51, 4122–4134. [Google Scholar] [CrossRef]
- Peoples, B.K.; Midway, S.R. Fishing Pressure and Species Traits Affect Stream Fish Invasions both Directly and Indirectly. Divers. Distrib. 2018, 24, 1158–1168. [Google Scholar] [CrossRef] [Green Version]
- Domenici, P.; Turesson, H.; Brodersen, J.; Brönmark, C. Predator-Induced Morphology Enhances Escape Locomotion in Crucian Carp. Proc. R. Soc. B Biol. Sci. 2008, 275, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.A. Dynamics of Pectoral Fin Rowing in a Fish with an Extreme Rowing Stroke: The Threespine Stickleback (Gasterosteus aculeatus). J. Exp. Biol. 2004, 207, 1925–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seebacher, F.; Webster, M.M.; James, R.S.; Tallis, J.; Ward, A.J.W. Morphological Differences between Habitats Are Associated with Physiological and Behavioural Trade-Offs in Stickleback (Gasterosteus aculeatus). R. Soc. Open Sci. 2016, 3, 160316. [Google Scholar] [CrossRef] [Green Version]
- Calazans, A.M.; Martinez, P.A.; Jacobina, U.P. Lentic and Lotic Environments Affect Morphological Diversity in Characiformes Fishes in the Neotropical São Francisco River Basin, Brazil. Environ. Biol. Fishes 2021, 104, 977–987. [Google Scholar] [CrossRef]
- Roff, D.A.; Fairbairn, D.J. The Evolution of Trade-Offs: Where Are We? J. Evol. Biol. 2007, 20, 433–447. [Google Scholar] [CrossRef]
- Pettersson, L.B.; Hedenström, A. Energetics, Cost Reduction and Functional Consequences of Fish Morphology. Proc. R. Soc. B Biol. Sci. 2000, 267, 759–764. [Google Scholar] [CrossRef] [Green Version]
- Li, X.M.; Yu, L.J.; Wang, C.; Zeng, L.Q.; Cao, Z.D.; Fu, S.J.; Zhang, Y.G. The Effect of Aerobic Exercise Training on Growth Performance, Digestive Enzyme Activities and Postprandial Metabolic Response in Juvenile Qingbo (Spinibarbus sinensis). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 166, 8–16. [Google Scholar] [CrossRef]
- Shrivastava, J.; Rašković, B.; Blust, R.; De Boeck, G. Exercise Improves Growth, Alters Physiological Performance and Gene Expression in Common Carp (Cyprinus carpio). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2018, 226, 38–48. [Google Scholar] [CrossRef]
- Li, X.M.; Liu, L.; Yuan, J.M.; Xiao, Y.Y.; Fu, S.J.; Zhang, Y.G. The Effect of Aerobic Exercise and Starvation on Growth Performance and Postprandial Metabolic Response in Juvenile Southern Catfish (Silurus meridionalis). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 193, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Guderley, H.; Pörtner, H.O. Metabolic Power Budgeting and Adaptive Strategies in Zoology: Examples from Scallops and Fish. Can. J. Zool. 2010, 88, 753–763. [Google Scholar] [CrossRef]
- Changes, B.; Marqueze, A.; Fagundes, M.; Rossini, M.; Variani, C.; Balbinoti, F.; Tietbo, H.; Rosemberg, D.B.; Jose, L.; Barcellos, G. Just Keep Swimming: Neuroendocrine, Metabolic, and Behavioral Changes After a Forced Swimming Test in Zebrafish. Zebrafish 2017, 14, 51–59. [Google Scholar] [CrossRef]
- Moves, C.D.; West, T.G. Exercise Metabolism of Fish. Biochem. Mol. Biol. Fishes 1995, 4, 367–392. [Google Scholar] [CrossRef]
- Parsons, S.A.; Wilkins, B.J.; Bueno, O.F.; Molkentin, J.D. Altered Skeletal Muscle Phenotypes in Calcineurin Aα and Aβ Gene-Targeted Mice. Mol. Cell. Biol. 2003, 23, 4331–4343. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A Nutrient and Energy Sensor That Maintains Energy Homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Pengam, M.; Amérand, A.; Simon, B.; Guernec, A.; Inizan, M.; Moisan, C. How Do Exercise Training Variables Stimulate Processes Related to Mitochondrial Biogenesis in Slow and Fast Trout Muscle Fibres? Exp. Physiol. 2021, 106, 938–957. [Google Scholar] [CrossRef]
- Pengam, M.; Moisan, C.; Simon, B.; Guernec, A.; Inizan, M.; Amérand, A. Training Protocols Differently Affect AMPK–PGC-1α Signaling Pathway and Redox State in Trout Muscle. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 243, 110673. [Google Scholar] [CrossRef]
- Craig, P.M.; Moyes, C.D.; LeMoine, C.M.R. Sensing and Responding to Energetic Stress: Evolution of the AMPK Network. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2018, 224, 156–169. [Google Scholar] [CrossRef]
- Conde-Sieira, M.; Capelli, V.; Álvarez-Otero, R.; Comesaña, S.; Liñares-Pose, L.; Velasco, C.; López, M.; Soengas, J.L. Differential Role of Hypothalamic AMPKα Isoforms in Fish: An Evolutive Perspective. Mol. Neurobiol. 2019, 56, 5051–5066. [Google Scholar] [CrossRef]
- Liu, G.; Wu, Y.; Qin, X.; Shi, X.; Wang, X. The Effect of Aerobic Exercise Training on Growth Performance, Innate Immune Response and Disease Resistance in Juvenile Schizothorax Prenanti. Aquaculture 2018, 486, 18–25. [Google Scholar] [CrossRef]
- Magnoni, L.J.; Crespo, D.; Ibarz, A.; Blasco, J.; Fernández-Borràs, J.; Planas, J.V. Effects of Sustained Swimming on the Red and White Muscle Transcriptome of Rainbow Trout (Oncorhynchus mykiss) Fed a Carbohydrate-Rich Diet. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 166, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Hood, D.A.; Irrcher, I.; Ljubicic, V.; Joseph, A.M. Coordination of Metabolic Plasticity in Skeletal Muscle. J. Exp. Biol. 2006, 209, 2265–2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyons, C.N.; Mathieu-Costello, O.; Moyes, C.D. Regulation of Skeletal Muscle Mitochondrial Content during Aging. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2006, 61, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClelland, G.B.; Craig, P.M.; Dhekney, K.; Dipardo, S. Temperature- and Exercise-Induced Gene Expression and Metabolic Enzyme Changes in Skeletal Muscle of Adult Zebrafish (Danio rerio). J. Physiol. 2006, 577, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Anttila, K.; Jäntti, M.; Mänttäri, S. Effects of Training on Lipid Metabolism in Swimming Muscles of Sea Trout (Salmo trutta). J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2010, 180, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Blasco, J.; Moya, A.; Millán-Cubillo, A.; Vélez, E.J.; Capilla, E.; Pérez-Sánchez, J.; Gutiérrez, J.; Fernández- Borrás, J. Growth-Promoting Effects of Sustained Swimming in Fingerlings of Gilthead Sea Bream (Sparus aurata L.). J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2015, 185, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Martin-Perez, M.; Fernandez-Borras, J.; Ibarz, A.; Millan-Cubillo, A.; Felip, O.; De Oliveira, E.; Blasco, J. New Insights into Fish Swimming: A Proteomic and Isotopic Approach in Gilthead Sea Bream. J. Proteome Res. 2012, 11, 3533–3547. [Google Scholar] [CrossRef]





| Gene | Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|
| AMPKα1 | TAGTGGCCGAGACTCAACCG | TCTGGCTACGGATTCCCAGG |
| AMPKα2 | AGTTCTACCTGGCCTCCAGT | GAGAGCATCCAGAGGGCATC |
| AMPKβ1a | TGGACAGCCCAGAAGATGC | TGGGTCGATCTAAAGTGAGGA |
| AMPKβ1b | TGGAGCTCGACCCAAAATCC | AACACAGTGGGCCTTTCCTC |
| AMPKβ2 | GAGCCCGGCAAAATGATGG | AGCCTGAGGACTCGTCTTT |
| AMPKγ1 | AACACACTCTACATCCTGACAC | CCAGCGTTTTAGACAGAAAAGCA |
| AMPKγ2a | TTCAAGTCCAGCTGTTTGGT | GACGTCTCCAGCCAATAGCA |
| AMPKγ2b | CACGAGACCCTGGAGACCAT | TGCCCATTCTGCATCTACCTG |
| AMPKγ3 | CCCTCACGAAACCCTCGAAA | CAGAGAGCGAGACGATTCCC |
| PGC-1α | TCGGCCCCAGTCTCATTCTA | ATACACCACTCGCCTCTCCT |
| NRF-1 | GTAGTGCAGACGATCAGTAATCC | GTCACTGTGGAGTAATTCACCTGG |
| β-actin | TGAAATTGCCGCACTGGTTG | CTGAGCCTCGTCACCAACAT |
| 18sRNA | GGCGCGCAAATTACCCATTT | TCCCGAGATCCAACTACAAGC |
| Parameters | Water Velocity | |||
|---|---|---|---|---|
| 0 Bl s−1 | 0.5 Bl s−1 | 2 Bl s−1 | 4 Bl s−1 | |
| Body weight (g) | 23.08 ± 5.53 a | 31.89 ± 13.78 b | 31.72 ± 14.67 b | 41.24 ± 18.91 c |
| Body length (cm) | 10.38 ± 0.68 a | 11.45 ± 1.38 b | 11.78 ± 1.42 b | 12.77 ± 1.36 c |
| Condition factor (CF) | 1.95 ± 0.11 a | 2.07 ± 0.18 b | 2.08 ± 0.15 b | 2.21 ± 0.21 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, P.; Liu, J.; Wang, Y.; Li, D. Lotic Environment Affects Morphological Characteristics and Energy Metabolism of Juvenile Grass Carp Ctenopharyngodon idella. Water 2022, 14, 1019. https://doi.org/10.3390/w14071019
Zhu P, Liu J, Wang Y, Li D. Lotic Environment Affects Morphological Characteristics and Energy Metabolism of Juvenile Grass Carp Ctenopharyngodon idella. Water. 2022; 14(7):1019. https://doi.org/10.3390/w14071019
Chicago/Turabian StyleZhu, Pengjin, Jieya Liu, Yin Wang, and Dapeng Li. 2022. "Lotic Environment Affects Morphological Characteristics and Energy Metabolism of Juvenile Grass Carp Ctenopharyngodon idella" Water 14, no. 7: 1019. https://doi.org/10.3390/w14071019
APA StyleZhu, P., Liu, J., Wang, Y., & Li, D. (2022). Lotic Environment Affects Morphological Characteristics and Energy Metabolism of Juvenile Grass Carp Ctenopharyngodon idella. Water, 14(7), 1019. https://doi.org/10.3390/w14071019