
Cryptophyte and Photosynthetic Picoeukaryote Abundances in the Bransfield Strait during Austral Summer


 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
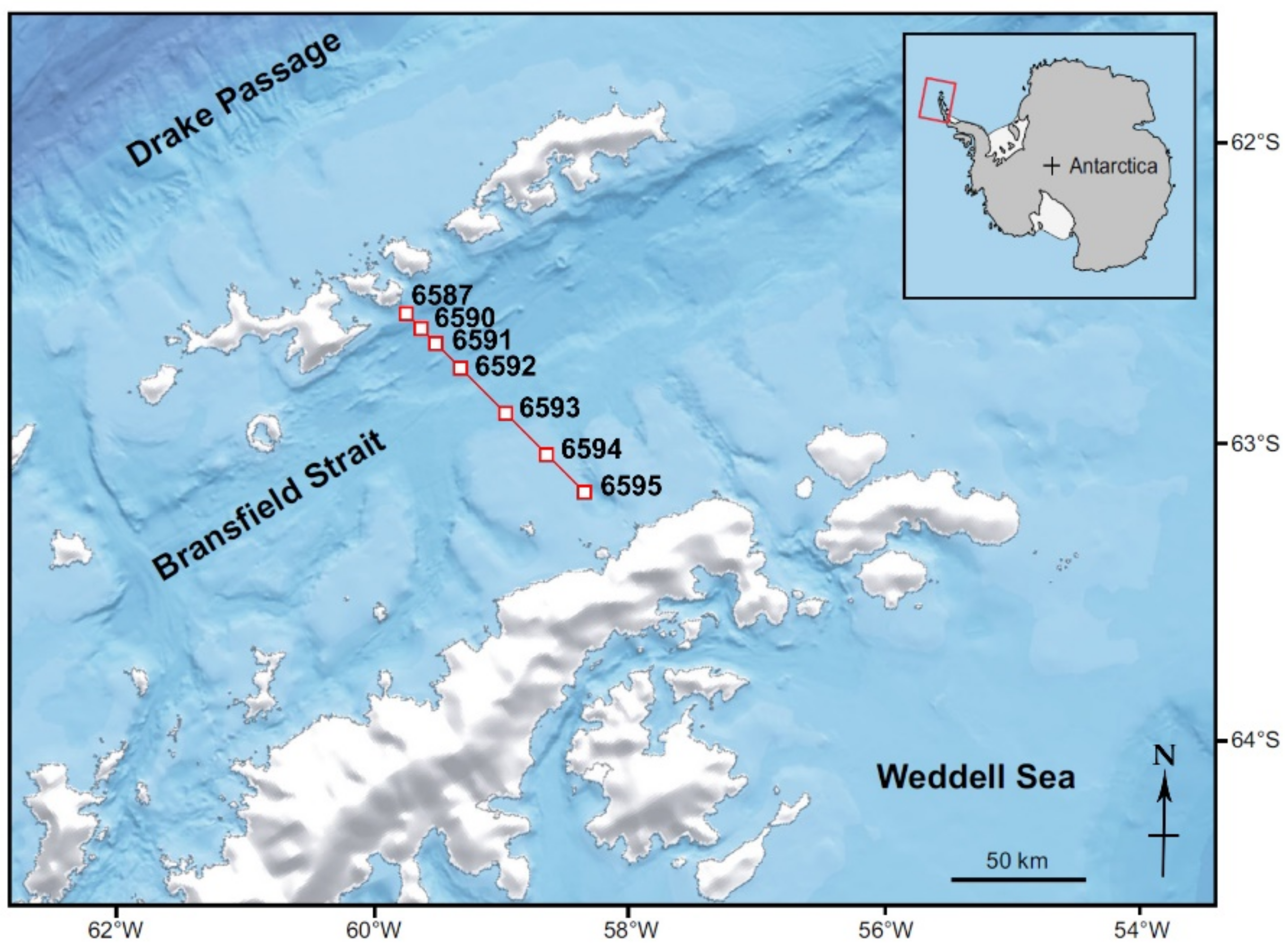
2.1. Sampling Sites and Methods
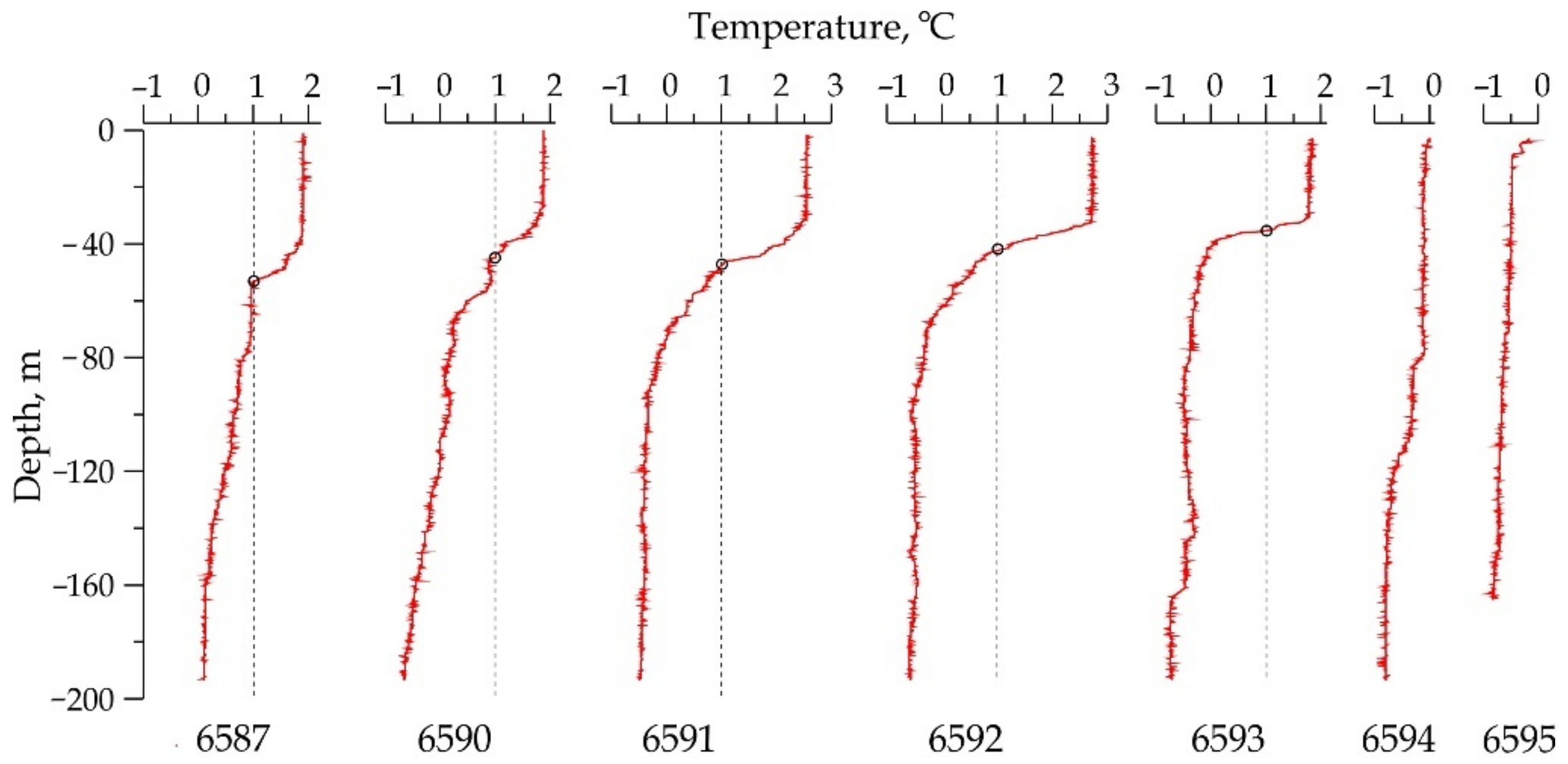
2.2. Physical and Chemical Oceanographic Data
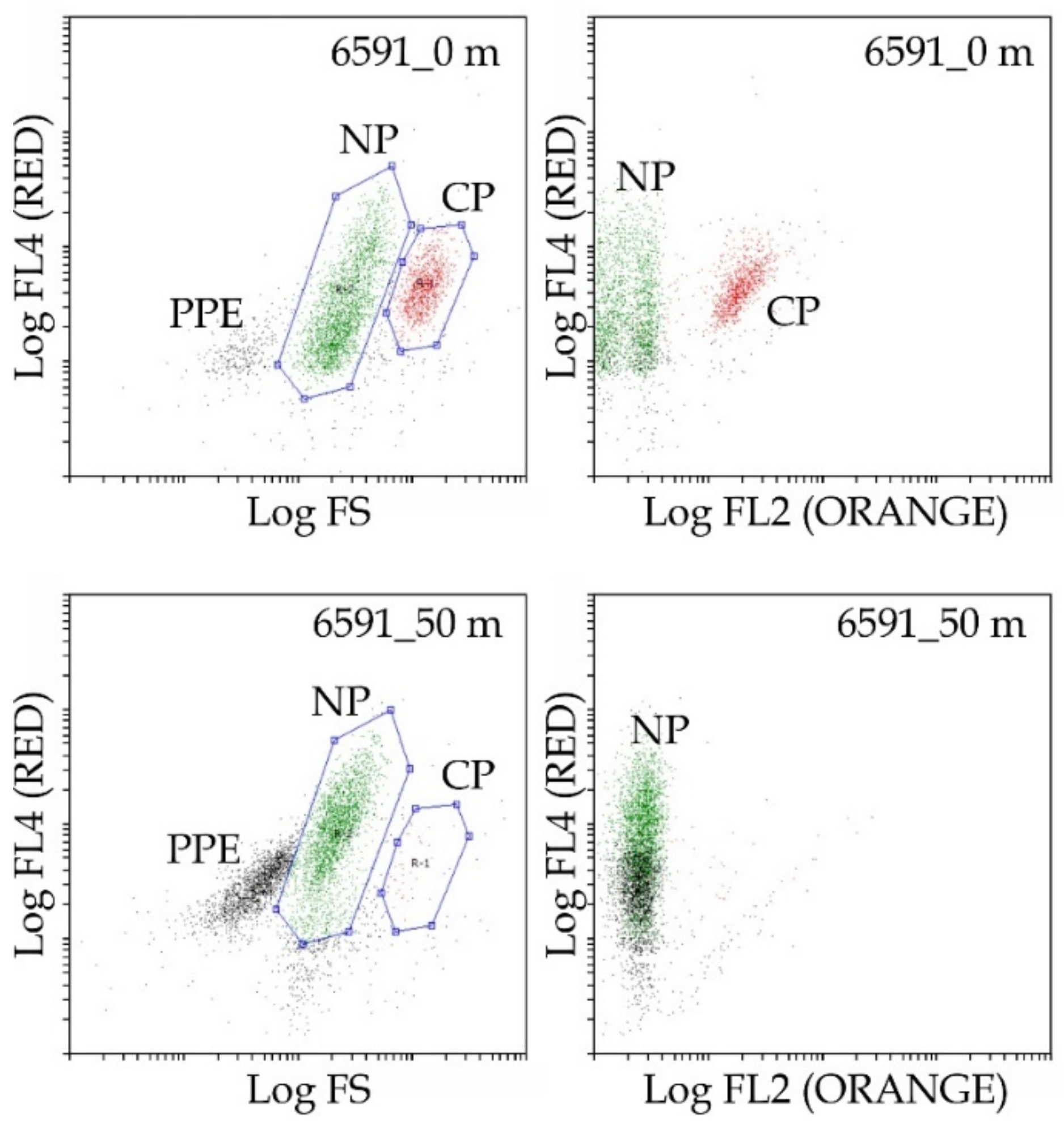
2.3. Flow Cytometry
2.4. Statistics
3. Results
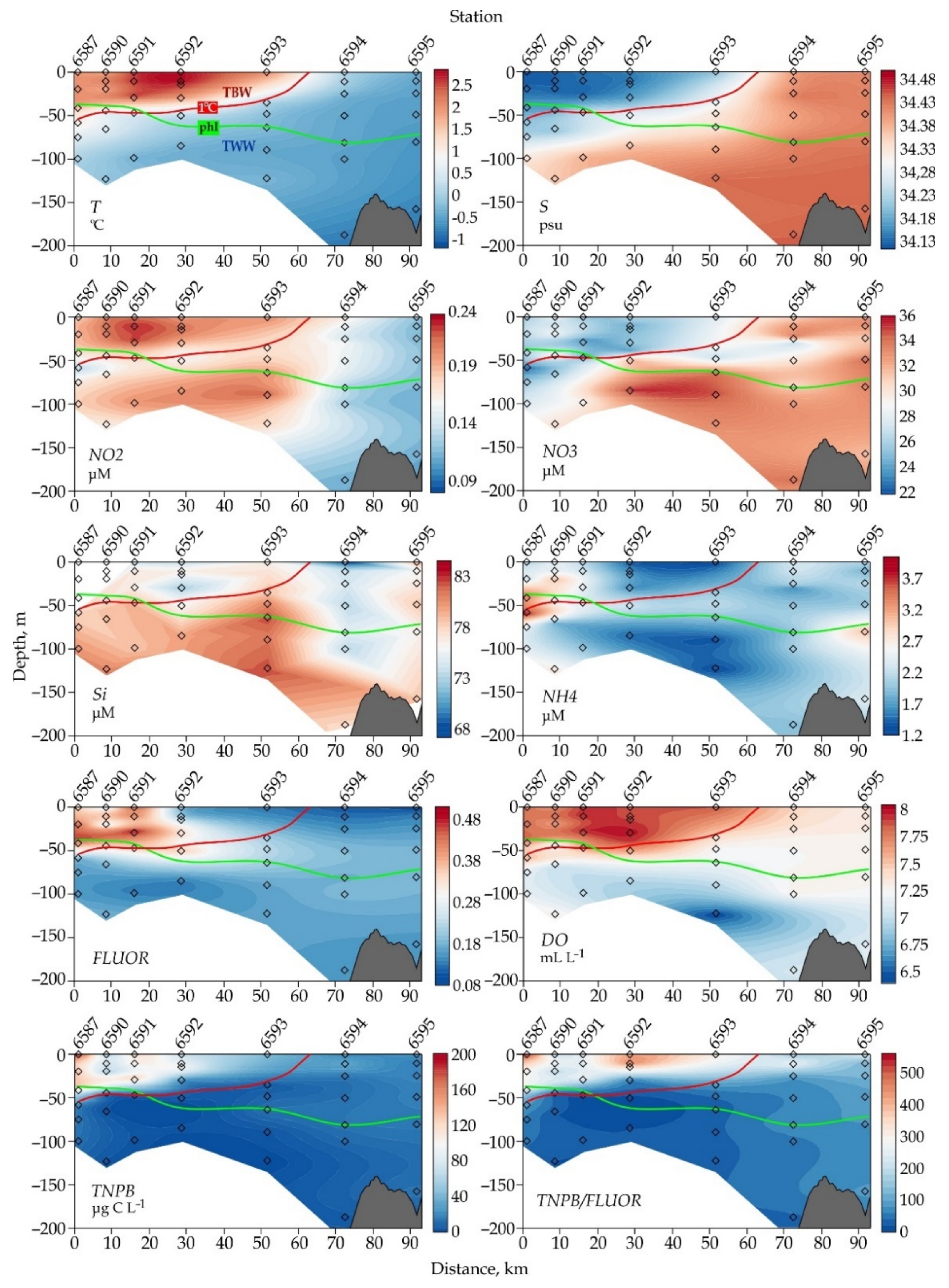
3.1. Description of Oceanographic Features
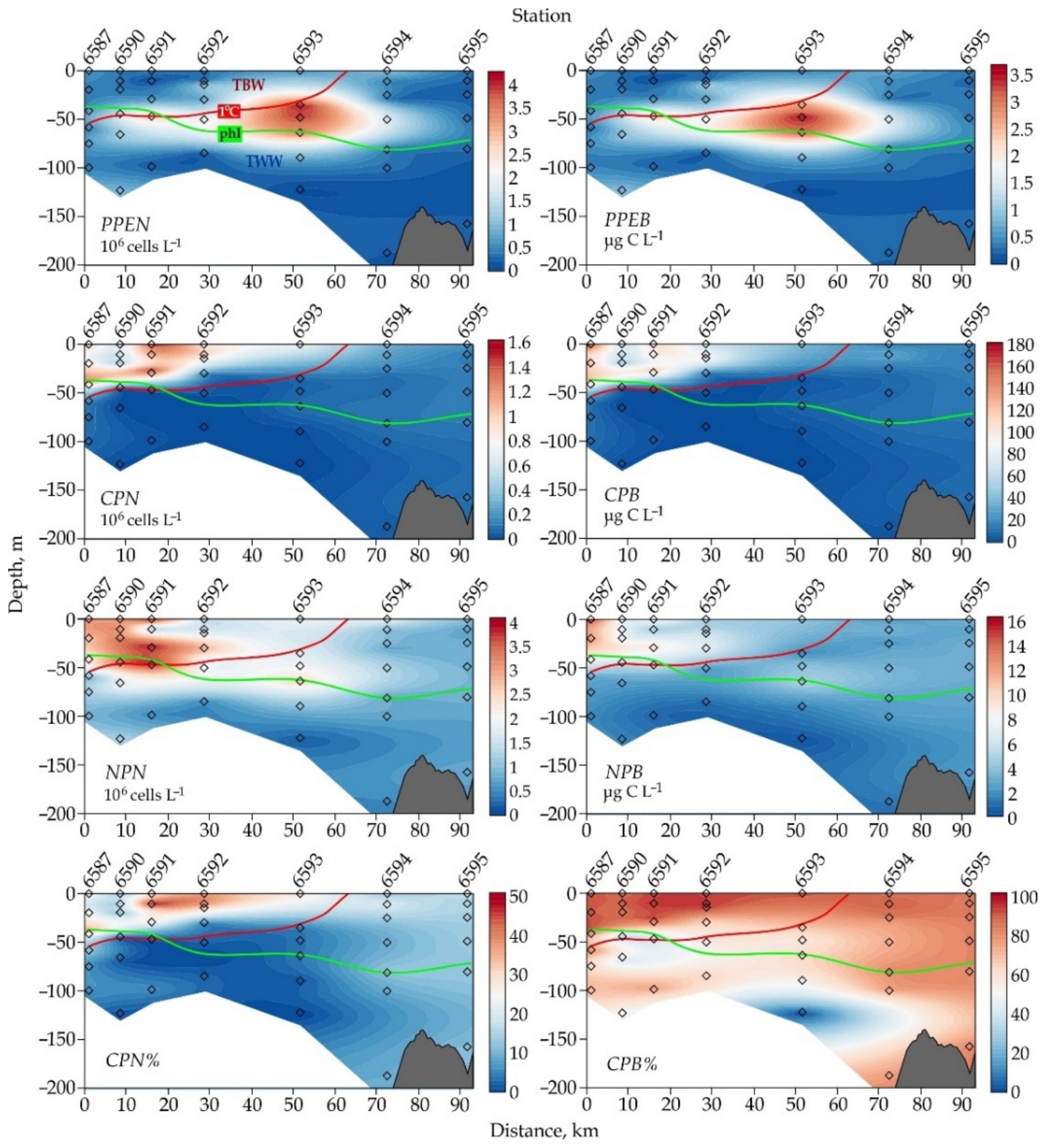
3.2. Picophytoplankton
3.3. Nanophytoplankton
4. Discussion
4.1. Picoeukaryotes
4.2. Cryptophyta
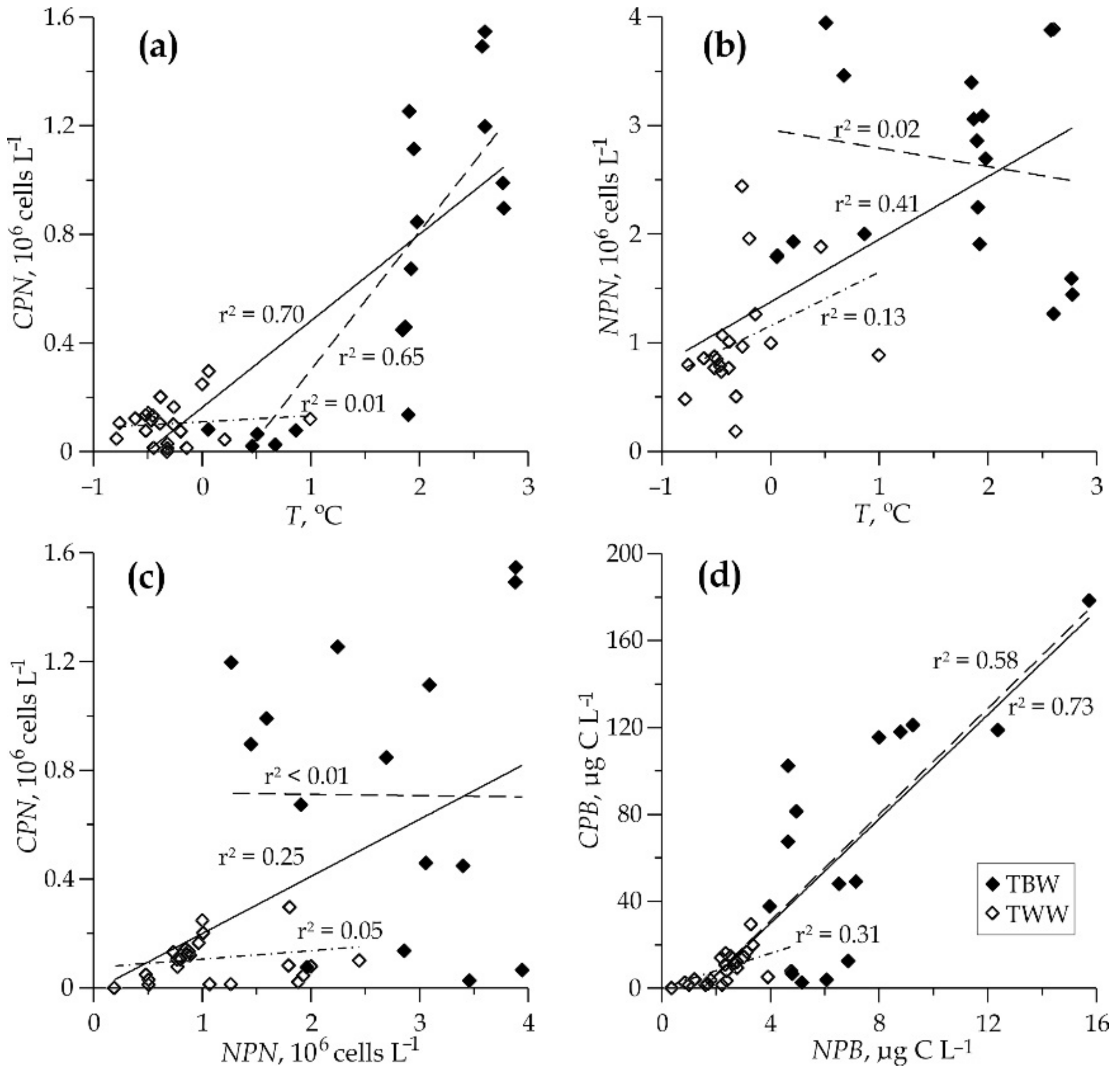
4.3. Linkage between Nanophytoplankton and Environmental Variables
4.4. Comparing Nanophytoplankton from TBW and TWW
4.5. Cryptophyta Distribution Pattern in the Bransfield Strait
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rodriguez, F.; Varela, M.; Zapata, M. Phytoplankton assemblages in the Gerlache and Bransfield Straits (Antarctic Peninsula) determined by light microscopy and CHEMTAX analysis of HPLC pigment data. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 723–747. [Google Scholar] [CrossRef]
- Garibotti, I.A.; Vernet, M.; Ferrario, M.E.; Smith, R.C.; Ross, R.M.; Quetin, L.B. Phytoplankton spatial distribution patterns along the western Antarctic Peninsula (Southern Ocean). Mar. Ecol. Prog. Ser. 2003, 261, 21–39. [Google Scholar] [CrossRef] [Green Version]
- Garibotti, I.A.; Vernet, M.; Ferrario, M.E. Annually recurrent phytoplanktonic assemblages during summer in the seasonal ice zone west of the Antarctic Peninsula (Southern Ocean). Deep Sea Res. Part I Oceanogr. Res. Pap. 2005, 52, 1823–1841. [Google Scholar] [CrossRef]
- Kozlowski, W.A.; Deutschman, D.; Garibotti, I.; Trees, C.; Vernet, M. An evaluation of the application of CHEMTAX to Antarctic coastal pigment data. Deep Sea Res. Part I Oceanogr. Res. Pap. 2011, 58, 350–364. [Google Scholar] [CrossRef]
- Mendes, C.R.B.; de Souza, M.S.; Garcia, V.M.T.; Leal, M.C.; Brotas, V.; Garcia, C.A.E. Dynamics of phytoplankton communities during late summer around the tip of the Antarctic Peninsula. Deep Sea Res. Part I Oceanogr. Res. Pap. 2012, 65, 1–14. [Google Scholar] [CrossRef]
- Prézelin, B.B.; Hofmann, E.E.; Mengelt, C.; Klinck, J.M. The linkage between Upper Circumpolar Deep Water (UCDW) and phytoplankton assemblages on the west Antarctic Peninsula continental shelf. J. Mar. Res. 2000, 58, 165–202. [Google Scholar] [CrossRef] [Green Version]
- Moline, M.A.; Prezelin, B.B. Long-term monitoring and analyses of physical factors regulating variability in coastal Antarctic phytoplankton biomass, in situ productivity and taxonomic composition over subseasonal, seasonal and interannual time scales. Mar. Ecol. Prog. Ser. 1996, 145, 143–160. [Google Scholar] [CrossRef]
- Moline, M.A.; Claustre, H.; Frazer, T.K.; Schofield, O.; Vernet, M. Alteration of the food web along the Antarctic Peninsula in response to a regional warming trend. Glob. Chang. Biol. 2004, 10, 1973–1980. [Google Scholar] [CrossRef]
- Gonçalves-Araujo, R.; de Souza, M.S.; Tavano, V.M.; Garcia, C.A.E. Influence of oceanographic features on spatial and interannual variability of phytoplankton in the Bransfield Strait, Antarctica. J. Mar. Syst. 2015, 142, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mendes, C.R.B.; Tavano, V.M.; Leal, M.C.; de Souza, M.S.; Brotas, V.; Garcia, C.A.E. Shifts in the dominance between diatoms and cryptophytes during three late summers in the Bransfield Strait (Antarctic Peninsula). Polar Biol. 2013, 36, 537–547. [Google Scholar] [CrossRef]
- Haberman, K.L.; Ross, R.M.; Quetin, L.B. Diet of the Antarctic krill (Euphausia superba Dana): II Selective grazing in mixed phytoplankton assemblages. J. Exper. Mar. Biol. Ecol. 2003, 283, 97–113. [Google Scholar] [CrossRef]
- Moline, M.A.; Claustre, H.; Frazer, T.K.; Grzymski, J.; Vernet, M. Changes in phytoplankton assemblages along the Antarctic Peninsula and potential implications for the Antarctic food web. In Antarctic Ecosystems: Models for Wider Ecological Understanding; Davidson, W., Howard-Williams, C., Broady, P., Eds.; University of Canterbury: Christchurch, New Zealand, 2001. [Google Scholar]
- McClatchie, S.; Boyd, C.M. Morphological study of sieve efficiencies and mandibular surfaces in the Antarctic krill, Euphausia superba. Can. J. Fish. Aquat. Sci. 1983, 40, 955–967. [Google Scholar] [CrossRef]
- Boyd, C.M.; Heyraud, M.; Boyd, C.N. Feeding of the Antarctic krill Euphausia superba. J. Crustacean Biol. 1984, 4, 123–141. [Google Scholar] [CrossRef]
- Montes-Hugo, M.; Doney, S.C.; Ducklow, H.W.; Fraser, W.; Martinson, D.; Stammerjohn, S.E.; Schofield, O. Recent changes in phytoplankton communities associated with rapid regional climate change along the western Antarctic Peninsula. Science 2009, 323, 1470–1473. [Google Scholar] [CrossRef] [PubMed]
- Loeb, V.; Siegel, V.; Holm-Hansen, O.; Hewitt, R.; Fraser, W.; Trivelpiece, W.; Trivelpiece, S. Effects of sea-ice extent and krill or salp dominance on the Antarctic food web. Nature 1997, 387, 897–900. [Google Scholar] [CrossRef]
- Castro, C.G.; Ríos, A.F.; Doval, M.D.; Pérez, F.F. Nutrient utilisation and chlorophyll distribution in the Atlantic sector of the Southern Ocean during Austral summer 1995–96. Deep Sea Res. 2002, 49, 623–641. [Google Scholar] [CrossRef]
- Mendes, C.R.B.; Tavano, V.M.; Dotto, T.S.; Kerr, R.; De Souza, M.S.; Garcia, C.A.E.; Secchi, E.R. New insights on the dominance of cryptophytes in Antarctic coastal waters: A case study in Gerlache Strait. Deep Sea Res. Part II Top. Stud. Oceanogr. 2018, 149, 161–170. [Google Scholar] [CrossRef]
- Polukhin, A.A.; Morozov, E.G.; Tishchenko, P.P.; Frey, D.I.; Artemiev, V.A.; Borisenko, G.V.; Vidnichuk, A.V.; Marina, E.N.; Medvedev, E.V.; Popov, O.S.; et al. Water structure in the Bransfield Strait (Antarctica) in January 2020: Hydrophysical, optical and hydrochemical features. Oceanology 2021, 61, 632–644. [Google Scholar] [CrossRef]
- Tokarczyk, R. Classification of water masses in the Bransfield Strait and southern part of the Drake Passage using a method of statistical multidimensional analysis. Pol. Res. 1987, 8, 333–366. [Google Scholar]
- Garcıa, M.; Castro, C.; Rıos, A. Water masses and distribution of physico-chemical properties in the Western Bransfield Strait and Gerlache Strait during Austral summer 1995/96. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 585–602. [Google Scholar] [CrossRef]
- Bordovsky, O.K.; Chernyakova, A.M. Modern Methods of Hydrochemical Studies of the Ocean; IO RAS: Moscow, Russia, 1992; pp. 1–200. (In Russian) [Google Scholar]
- Solorzano, L. Determination of ammonia in natural waters by phenol-hypochlorite method. Limnol. Oceanogr. 1969, 14, 799–801. [Google Scholar]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis, 2nd ed.; Bulletin of Fisheries Research Board of Canada: Ottawa, ON, Canada, 1972; pp. 1–328. [Google Scholar]
- Grasshoff, K.; Kremling, K.; Ehrhardt, M. Methods of Seawater Analysis; John Wiley & Sons: New York, NY, USA, 2009; pp. 1–599. [Google Scholar]
- Parsons, T.R. A Manual of Chemical & Biological Methods for Seawater Analysis; Elsevier: Amsterdam, The Netherlands, 2013; pp. 1–188. [Google Scholar]
- Hoef-Emden, K.; Marin, B.; Melkonian, M. Nuclear and nucleomorph SSU rDNA phylogeny in the Cryptophyta and the evolution of cryptophyte diversity. J. Mol. Evol. 2002, 55, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Six, C.; Thomas, J.C.; Brahamsha, B.; Lemoine, Y.; Partensky, F. Photophysiology of the marine cyanobacteria Synechococcus sp. WH8102, a new model organism. Aquat. Microb. Ecol. 2004, 35, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Marie, D.; Simon, N.; Vaulot, D. Phytoplankton cell counting by flow cytometry. In Algal Culturing Techniques; Andersen, R.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 253–267. [Google Scholar]
- Olson, R.J.; Zettler, E.R.; DuRand, M.D. Phytoplankton analysis using flow cytometry. In Handbook of Methods in Aquatic Microbial Ecology; CRC Press: Boca Raton, FL, USA, 1993; pp. 175–186. [Google Scholar]
- Alvarez, E.; Nogueira, E.; López-Urrutia, Á. In vivo single-cell fluorescence and size scaling of phytoplankton chlorophyll content. Appl. Environ. Microbiol. 2017, 83, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worden, A.; Nolan, J.; Palenik, B. Assessing the dynamics and ecology of marine picophytoplankton: The importance of the eukaryotic component. Limnol. Oceanogr. 2004, 49, 168–179. [Google Scholar] [CrossRef]
- Garrison, D.; Gowing, M.; Hughes, M. Microbial food web structure in the Arabian Sea: A US JGOFS study. Deep Sea Res. Part II 2000, 47, 1387–1422. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Paleontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Zhou, M.; Niiler, P.P.; Zhu, Y.; Dorland, R.D. The western boundary current in the Bransfield Strait, Antarctica. Deep Sea Res. I 2006, 53, 1244–1252. [Google Scholar] [CrossRef]
- Niiler, P.P.; Amos, A.; Hu, J.-H. Water masses and 200 m relative geostrophic circulation in the western Bransfield Strait region. Deep Sea Res. 1991, 38, 943–959. [Google Scholar] [CrossRef]
- Garcia, M.A.; López, O.; Sospedra, J.; Espino, M.; Gracia, V.; Morrison., G.; Arcilla, A.S. Mesoscale variability in the Bransfield Strait region (Antarctica) during Austral summer. Ann. Geophys. 1994, 12, 856–867. [Google Scholar] [CrossRef]
- Sangrà, P.; Gordo, C.; Hernández-Arencibia, M.; Marrero-Díaz, A.; Rodríguez-Santana, A.; Stegner, A.; Pichon, T. The Bransfield current system. Deep Sea Res. I 2011, 58, 390–402. [Google Scholar] [CrossRef]
- Jiang, M.; Charette, M.A.; Measures, C.I.; Zhu, Y.; Zhou, M. Seasonal cycle of circulation in the Antarctic Peninsula and the off-shelf transport of shelf waters into southern Drake Passage and Scotia Sea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2013, 90, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Morozov, E.G. Currents in Bransfield Strait. Dokl. Earth Sci. 2007, 415, 984–986. [Google Scholar] [CrossRef]
- Holm-Hansen, O.; Mitchell, B.G. Spatial and temporal distribution of phytoplankton and primary production in the western Bransfield Strait region. Deep Sea Res. II 1991, 39, 961–980. [Google Scholar] [CrossRef]
- Dierssen, H.M.; Smith, R.C. Case 2 Antarctic coastal waters: The bio-optical properties of surface meltwater. In Proceedings of the Ocean Optics XV, Kailua-Kona, HI, USA, 16–20 October 2000. [Google Scholar]
- McMinn, A.; Hodgson, D. Summer phytoplankton succession in Ellis Fjord, eastern Antarctica. J. Plankton Res. 1993, 15, 925–938. [Google Scholar] [CrossRef]
- Bouman, H.A.; Ulloa, O.; Barlow, R.; Li, W.K.; Platt, T.; Zwirglmaier, K.; Sathyendranath, S. Water-column stratification governs the community structure of subtropical marine picophytoplankton. Environ. Microbiol. Rep. 2011, 3, 473–482. [Google Scholar] [CrossRef]
- Buitenhuis, E.T.; Li, W.K.; Vaulot, D.; Lomas, M.W.; Landry, M.R.; Partensky, F.; McManus, G.B. Picophytoplankton biomass distribution in the global ocean. Earth Syst. Sci. Data 2012, 4, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Agawin, N.S.; Agustí, S.; Duarte, C.M. Abundance of Antarctic picophytoplankton and their response to light and nutrient manipulation. Aquat. Microb. Ecol. 2002, 29, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Weber, L.H.; El-Sayed, S.Z. Contribution of the net-, nano and picoplankton standing crop and primary productivity in the Southern Ocean. J. Plankton Res. 1987, 9, 973–994. [Google Scholar] [CrossRef]
- Lin, L.; He, J.; Zhao, Y.; Zhang, F.; Cai, M. Flow cytometry investigation of picoplankton across latitudes and along the circum Antarctic Ocean. Acta Oceanol. Sin. 2012, 31, 134–142. [Google Scholar] [CrossRef]
- Morris, I.; Glover, H. Physiology of photosynthesis by marine coccoid cyanobacteria—Some ecological implications. Limnol. Oceanogr. 1981, 26, 957–961. [Google Scholar] [CrossRef]
- Sakshaug, E.; Holm-Hansen, O. Photoadaptation in Antarctic phytopfankton: Variations in growth rate, chemical composition and P versus I curves. J. Plankton Res. 1986, 8, 459–473. [Google Scholar] [CrossRef]
- Mitchell, B.G.; Holm-Hansen, O. Observations of modeling of the Antarctic phytoplankton crop in relation to mixing depth. Deep Sea Res. Part A Oceanogr. Res. Pap. 1991, 38, 981–1007. [Google Scholar] [CrossRef]
- Vaillancourt, R.D.; Brown, C.W.; Guillard, R.R.L.; Balch, W.M. Light scattering properties of marine phytoplankton: Relationships to cell size, chemical composition and taxonomy. J. Plankton Res. 2004, 26, 191–212. [Google Scholar] [CrossRef]
- Eriksen, N.T.; Poulsen, B.R.; Iversen, J.L. Dual sparging laboratory-scale photobioreactor for continuous production of microalgae. J. Appl. Phycol. 1998, 10, 377–382. [Google Scholar] [CrossRef]
- Jacques, G.; Panouse, M. Biomass and composition of size fractionated phytoplankton in the Weddell-Scotia Confluence area. Polar Biol. 1991, 5, 315–328. [Google Scholar] [CrossRef]
- Kopczynska, E.E. Distribution of microflagellates and diatoms in the sea-ice zone between Elephant Island and the South Orkney Islands (December 1988–January 1989). Pol. Polar Res. 1991, 12, 515–528. [Google Scholar]
- Buma, A.G.J.; Gieskes, W.W.C.; Thomsen, H.A. Abundance of Cryptophyceae and chlorophyll b-containing organisms in the Weddell-Scotia Confluence area in the spring of 1988. In Weddell Sea Ecology; Hempel, G., Ed.; Springer: Berlin/Heidelberg, Germany, 1992; pp. 43–52. [Google Scholar]
- Kang, S.-H.; Lee, S. Antarctic phytoplankton assemblage in the western Bransfield Strait region, February 1993: Composition, biomass, and mesoscale distributions. Mar. Ecol. Prog. Ser. 1995, 129, 253–267. [Google Scholar] [CrossRef] [Green Version]
- Holm-Hansen, O.; Mitchell, B.G.; Hewes, C.D.; Karl, D.M. Phytoplankton blooms in the vicinity of Palmer Station, Antarctica. Polar Biol. 1989, 10, 49–57. [Google Scholar] [CrossRef]
- Smith, W.O., Jr.; Sakshaug, E. Polar Phytoplankton. In Polar Oceanography, Part B; Smith, W.O., Jr., Ed.; Academic Press: London, UK, 1990; pp. 477–525. [Google Scholar]
- García-Muñoz, C.; Lubián, L.M.; García, C.M.; Marrero-Díaz, Á.; Sangrà, P.; Vernet, M. A mesoscale study of phytoplankton assemblages around the South Shetland Islands (Antarctica). Polar Biol. 2013, 36, 1107–1123. [Google Scholar] [CrossRef] [Green Version]
- Mura, M.P.; Satta, M.P.; Agustí, S. Water-mass influences on summer Antarctic phytoplankton biomass and community structure. Polar Biol. 1995, 15, 15–20. [Google Scholar] [CrossRef]
- Sangrà, P.; García-Muñoz, C.; García, C.M.; Marrero-Díaz, Á.; Sobrino, C.; Mouriño-Carballido, B.; Hernández-Arencibia, M. Coupling between upper ocean layer variability and size-fractionated phytoplankton in a non-nutrient-limited environment. Mar. Ecol. Prog. Ser. 2014, 499, 35–46. [Google Scholar] [CrossRef]
- Hewes, C.D.; Reiss, C.S.; Holm-Hansen, O. A quantitative analysis of sources for summertime phytoplankton variability over 18 years in the South Shetland Islands (Antarctica) region. Deep Sea Res. Part I Oceanogr. Res. Pap. 2009, 56, 1230–1241. [Google Scholar] [CrossRef]
- Holm-Hansen, O.; Hewes, C.D.; Villafane, V.E.; Helbling, E.W.; Silva, N.; Amos, T. Distribution of phytoplankton and nutrients in relation to different water masses in the area around Elephant Island, Antarctica. Polar Biol. 1997, 18, 145–153. [Google Scholar] [CrossRef]
- Schofield, O.; Saba, G.; Coleman, K.; Carvalho, F.; Couto, N.; Ducklow, H.; Montes-Hugo, M. Decadal variability in coastal phytoplankton community composition in a changing West Antarctic Peninsula. Deep Sea Res. Part I Oceanogr. Res. Pap. 2017, 124, 42–54. [Google Scholar] [CrossRef]
- Moreau, S.; Ferreyra, G.A.; Mercier, B.; Lemarchand, K.; Lionard, M.; Roy, S.; Mostajir, B.; Van Hardenberg, B.; Demers, S. Variability of the microbial community in the western Antarctic Peninsula from late fall to spring during a low ice cover year. Polar Biol. 2010, 33, 1599–1614. [Google Scholar] [CrossRef]
- Ferreyra, G.A.; Tomo, A.P. Variacion estational de las diatomeas planctonicas en Puerto Paraiso-I. Contrib. Del Inst. Antart. Argentio 1979, 264, 149–184. [Google Scholar]
- Whitaker, T.M. Primary production of phytoplankton off Signy Island, South Orkneys, the Antarctic. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1982, 214, 169–189. [Google Scholar]
- Krebs, W.N. Ecology of neritic marine diatoms, Arthur Harbor, Antarctica. Micropaleontology 1983, 29, 267–297. [Google Scholar] [CrossRef]
- Kopczynska, E.E. Dominance of microflagellates over diatoms in the Antarctic areas of deep vertical mixing and krill concentrations. J. Plankton Res. 1992, 14, 1031–1054. [Google Scholar] [CrossRef]
- Walsh, J.J.; Dieterle, D.A.; Lenes, J. A numerical analysis of carbon dynamics of the Southern Ocean phytoplankton community: The roles of light and grazing in effecting both sequestration of atmospheric CO2 and food availability to larval krill. Deep Sea Res. 2001, 48, 1–48. [Google Scholar] [CrossRef]
- Ainley, D.G.; Ballard, G.; Emslie, S.D. Adélie penguins and environmental change. Science 2003, 300, 429–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Definition, Units | Min | Max | Mean ± SD |
|---|---|---|---|---|
| T | Water temperature, °C | −0.79 | 2.77 | 0.59 ± 1.19 |
| S | Water salinity, psu | 34.1 | 34.4 | 34.3 ± 0.1 |
| PO4 | Phosphates, µM | 1.56 | 4.37 | 3.09 ± 0.58 |
| Si | Silicates, µM | 67.9 | 112.3 | 81.3 ± 10.1 |
| NO3 | Nitrates, µM | 22.6 | 35.4 | 29.4 ± 3.7 |
| NO2 | Nitrites, µM | 0.11 | 0.23 | 0.17 ± 0.04 |
| NH4 | Ammonium, µM | 1.32 | 3.81 | 2.13 ± 0.58 |
| DO | Dissolved oxygen, mL L−1 | 6.44 | 7.98 | 7.44 ± 0.36 |
| FLUOR | Chl-a fluorescence, r. u. | 0.10 | 0.49 | 0.24 ± 0.12 |
| CPN | Cryptophyte abundance, 106 cells L−1 | 0.00 | 1.55 | 0.35 ± 0.46 |
| CP-ESD | Mean CP ESD, µm | 6.70 | 11.59 | 9.42 ± 1.77 |
| CPB | CP biomass, µg C L−1 | 0 | 179 | 33 ± 45 |
| CP-CHL | Intracellular Chl-a content in CP, r. u. cell −1 | 0 | 163 | 69 ± 39 |
| CP-PE | Intracellular phycoerythrin content in CP, r. u. cell −1 | 0 | 206 | 31 ± 39 |
| NPN | NP abundance, 106 cells L−1 | 0.19 | 3.95 | 1.71 ± 1.07 |
| NP-ESD | Mean NP ESD, µm | 2.14 | 3.67 | 2.88 ± 0.38 |
| NPB | NP biomass, µg C L−1 | 0.4 | 15.7 | 4.3 ± 3.2 |
| NP-CHL | Intracellular Chl-a content in NP, r. u. cell −1 | 8 | 220 | 74 ± 52 |
| TNPB | Total NP+CP biomass, µg C L−1 | 0.4 | 194.2 | 37.3 ± 47.7 |
| CPN% | Proportion of CP in the total abundance, % | 0 | 49 | 14 ± 12 |
| CPB% | Proportion of CP in the total biomass, % | 0 | 96 | 74 ± 21 |
| PPEN | PPE abundance, 106 cells L−1 | 0.06 | 4.18 | 0.87 ± 1.03 |
| PPE-ESD | Mean PPE ESD, µm | 1.72 | 2.09 | 1.90 ± 0.11 |
| PPEB | PPE biomass, µg C L−1 | 0.04 | 3.50 | 0.72 ± 0.83 |
| Variable | T | S | PO4 | Si | NO3 | NO2 | NH4 | DO | FLUOR |
|---|---|---|---|---|---|---|---|---|---|
| CPN | 0.59 | −0.53 | −0.22 | −0.22 | −0.45 | 0.29 | 0.20 | 0.78 | 0.37 |
| CP-ESD | −0.24 | 0.02 | −0.14 | 0.28 | 0.29 | −0.18 | 0.27 | −0.15 | −0.11 |
| CPB | 0.54 | −0.52 | −0.23 | −0.16 | −0.40 | 0.25 | 0.21 | 0.74 | 0.36 |
| CP-CHL | −0.12 | −0.06 | −0.32 | 0.27 | 0.09 | −0.41 | 0.62 | 0.04 | −0.04 |
| CP-PE | 0.10 | −0.06 | −0.01 | −0.13 | 0.03 | 0.00 | 0.28 | 0.11 | −0.03 |
| NPN | 0.74 | −0.81 | 0.32 | 0.24 | −0.59 | 0.56 | 0.32 | 0.60 | 0.73 |
| NP-ESD | −0.02 | 0.01 | −0.32 | −0.17 | −0.10 | −0.26 | 0.08 | 0.31 | 0.06 |
| NPB | 0.68 | −0.75 | 0.12 | 0.10 | −0.63 | 0.42 | 0.25 | 0.73 | 0.73 |
| NP-CHL | 0.01 | −0.14 | 0.04 | 0.44 | −0.16 | −0.12 | 0.34 | −0.15 | 0.12 |
| TNPB | 0.59 | −0.57 | −0.19 | −0.14 | −0.45 | 0.27 | 0.22 | 0.77 | 0.42 |
| CPN% | 0.43 | −0.33 | −0.31 | −0.32 | −0.30 | 0.15 | 0.07 | 0.65 | 0.15 |
| CPB% | 0.47 | −0.41 | −0.30 | −0.25 | −0.30 | 0.25 | 0.15 | 0.65 | 0.19 |
| PPEN | 0.35 | −0.39 | 0.41 | 0.13 | −0.30 | 0.24 | 0.17 | 0.21 | 0.45 |
| PPE-ESD | −0.85 | 0.78 | −0.28 | −0.04 | 0.62 | −0.68 | −0.04 | −0.62 | −0.47 |
| PPEB | 0.06 | −0.11 | 0.34 | 0.09 | −0.09 | −0.03 | 0.17 | 0.02 | 0.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mukhanov, V.; Sakhon, E.; Polukhin, A.; Artemiev, V.; Morozov, E.; Tsai, A.-Y. Cryptophyte and Photosynthetic Picoeukaryote Abundances in the Bransfield Strait during Austral Summer. Water 2022, 14, 185. https://doi.org/10.3390/w14020185
Mukhanov V, Sakhon E, Polukhin A, Artemiev V, Morozov E, Tsai A-Y. Cryptophyte and Photosynthetic Picoeukaryote Abundances in the Bransfield Strait during Austral Summer. Water. 2022; 14(2):185. https://doi.org/10.3390/w14020185
Chicago/Turabian StyleMukhanov, Vladimir, Evgeny Sakhon, Alexander Polukhin, Vladimir Artemiev, Eugene Morozov, and An-Yi Tsai. 2022. "Cryptophyte and Photosynthetic Picoeukaryote Abundances in the Bransfield Strait during Austral Summer" Water 14, no. 2: 185. https://doi.org/10.3390/w14020185
APA StyleMukhanov, V., Sakhon, E., Polukhin, A., Artemiev, V., Morozov, E., & Tsai, A. -Y. (2022). Cryptophyte and Photosynthetic Picoeukaryote Abundances in the Bransfield Strait during Austral Summer. Water, 14(2), 185. https://doi.org/10.3390/w14020185