


Fate of Saxitoxins in Lake Water: Preliminary Testing of Degradation by Microbes and Sunlight

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling and Location
2.2. Analysis of Intracellular Pools and Dissolved, Extracellular Pools of STXs in the Lake Water
2.3. Quantification of STX-Producing Cyanobacteria in the Lake Waters
2.4. Bacterial Degradation of STXs in the Lake Water
2.5. Isolation of STX-Degrading Bacteria from Lake Water Samples
2.6. Effect of Natural and Simulated Solar Conditions on STX Levels Using SaWa® Bags
3. Results
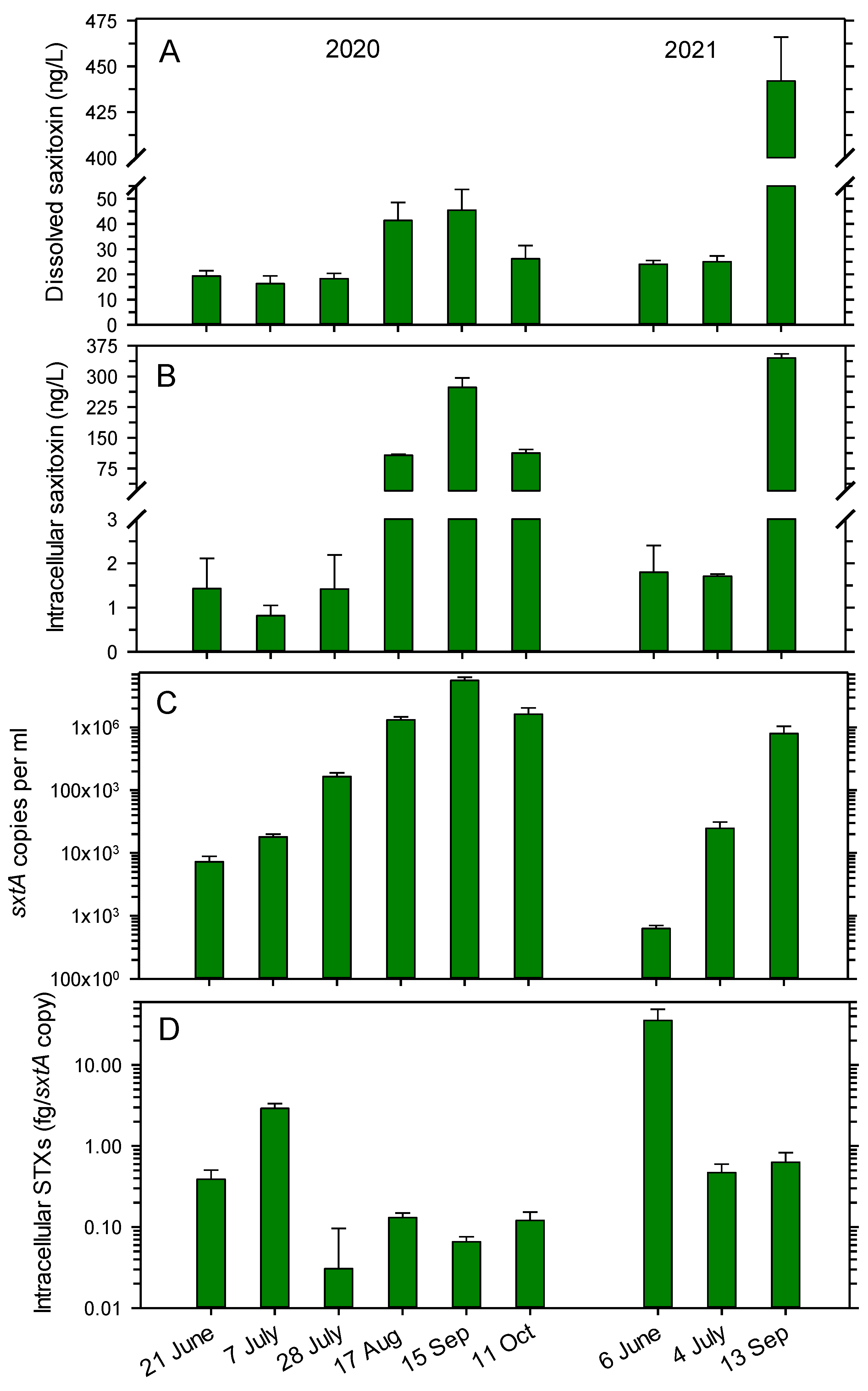
3.1. Occurrence of STX in the Lake Water and Number of sxtA Copies
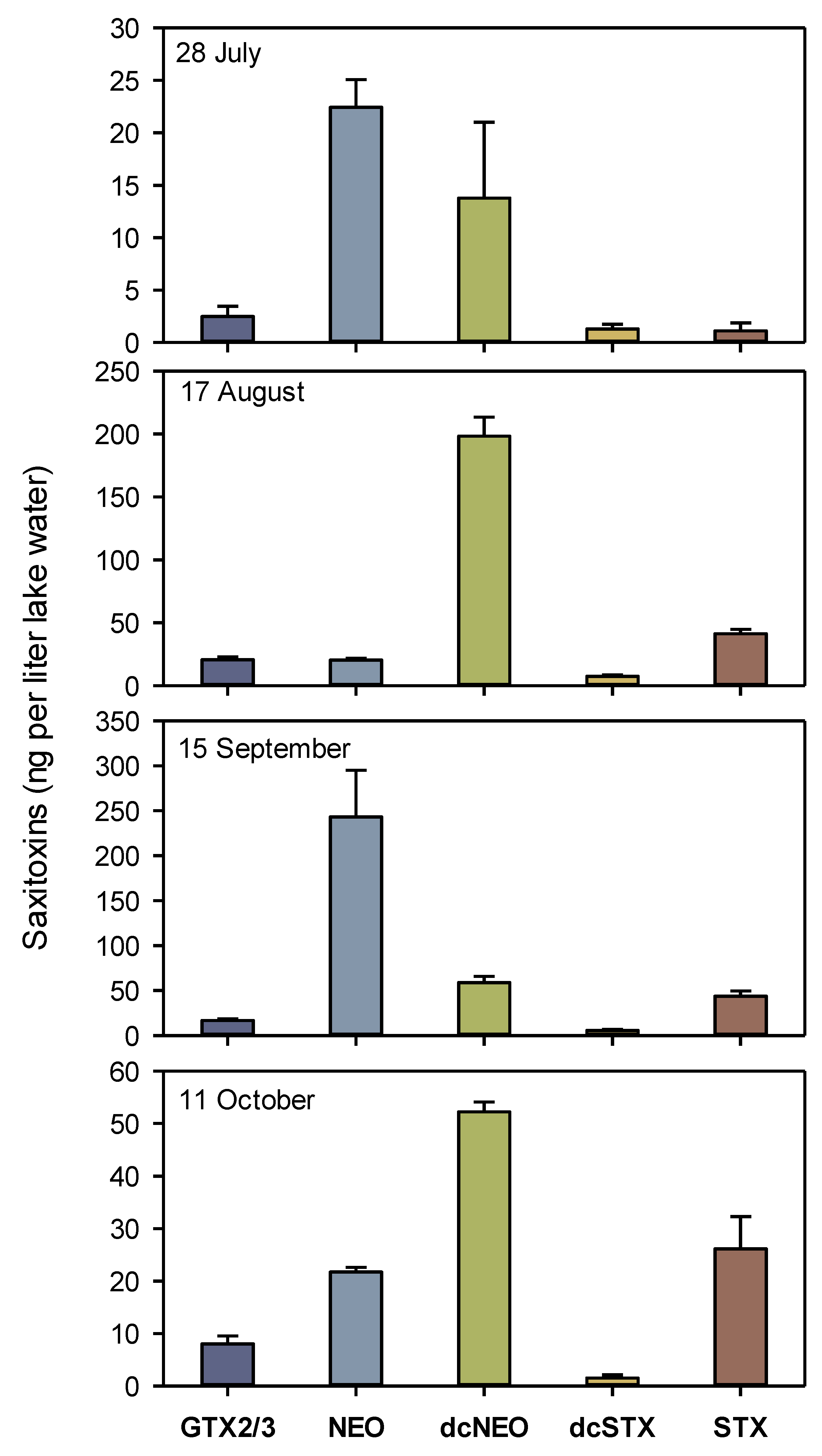
3.2. Concentrations of STX Analogs in Lake Water in 2020
3.3. Bacterial Degradation of STX in the Lake Water
3.4. Sunlight-Induced Effects on STX Concentrations
3.5. Microbial Degradation of STX Analogs in the Lake Water
4. Discussion
4.1. Concentrations of Dissolved STX and STX Analogs
4.2. Abundance of STX Producers and Intracellular Pools of STXs
4.3. Degradation of STX by Lake Water Microorganisms
4.4. Effects of Natural and Simulated Solar Radiation on STX and Its Biosynthesis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akbar, M.A.; Yusof, N.Y.M.; Tahir, N.I.; Ahmad, A.; Usup, G.; Sahrani, F.K.; Bunawan, H. Biosynthesis of saxitoxin in marine dinoflagellates: An omics perspective. Mar. Drugs 2020, 18, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negri, A.P.; Jones, G.J.; Hindmarsh, M. Sheep mortality associated with paralytic shellfish poisons from the cyanobacterium Anabaena circinalis. Toxicon 1995, 33, 1321–1329. [Google Scholar] [CrossRef]
- Smith, Z.J.; Martin, R.M.; Wei, B.; Wilhelm, S.W.; Boyer, G.L. Spatial and temporal variation in paralytic shellfish toxin production by benthic Microseira (Lyngbya) wollei in a freshwater New York lake. Toxins 2019, 11, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, L.S.; Dörr, F.; Dörr, F.A.; Frascareli, D.; Melo, D.S.; Gontijo, E.S.J.; Friese, K.; Pinto, E.; Rosa, A.H.; Pompêo, M.M.; et al. Permanent occurrence of Raphidiopsis raciborskii and cyanotoxins in a subtropical reservoir polluted by domestic effluents (Itupararanga reservoir, São Paulo, Brazil). Environ. Sci. Pollut. Res. 2022, 29, 18653–18664. [Google Scholar] [CrossRef] [PubMed]
- Bridgeman, T.B.; Penamon, W.A. Lyngbya wollei in Western Lake Erie. J. Great Lakes Res. 2010, 36, 167–171. [Google Scholar] [CrossRef]
- Yilmaz, M.; Foss, A.J.; Selwood, A.I.; Özen, M.; Boundy, M. Paralytic shellfish toxin producing Aphanizomenon gracile strains isolated from Lake Iznik, Turkey. Toxicon 2018, 148, 132–142. [Google Scholar] [CrossRef]
- Casali, S.P.; dos Santos, A.C.A.; Falco, P.B.; Calijuri, M.C. Influence of environmental variables on saxitoxin yields by Cylindrospermopsis raciborskii in a mesotrophic subtropical reservoir. J. Water Health 2017, 15, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Podduturi, R.; Schlüter, L.; Liu, T.; Osti, J.A.S.; Moraes, M.A.B.; Jørgensen, N.O.G. Monitoring of saxitoxin production in lakes in Denmark by molecular, chromatographic and microscopic approaches. Harmful Algae 2021, 101, 101966. [Google Scholar] [CrossRef]
- Ledreux, A.; Thomazeau, S.; Catherine, A.; Duval, C.; Yéprémian, C.; Marie, A.; Bernard, C. Evidence for saxitoxins production by the cyanobacterium Aphanizomenon gracile in a French recreational water body. Harmful Algae 2010, 10, 88–97. [Google Scholar] [CrossRef]
- Rapala, J.; Robertson, A.; Negri, A.P.; Berg, K.A.; Tuomi, P.; Lyra, C.; Erkomaa, K.; Lahti, K.; Hoppu, K.; Lepistö, L. First report of saxitoxin in Finnish lakes and possible associated effects on human health. Environ Toxicol 2005, 20, 331–340. [Google Scholar] [CrossRef]
- Cirés, S.; Delgado, A.; González-Pleiter, M.; Quesada, A. Temperature influences the production and transport of saxitoxin and the expression of sxt genes in the cyanobacterium Aphanizomenon gracile. Toxins 2017, 9, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization—WHO. Cyanobacterial toxins: Saxitoxins. In Background Document for Development of WHO for Drinking-Water Quality and Guidelines for Safe Recreational Water Environments; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Clemente, Z.; Busato, R.H.; Ribeiro, C.A.O.; Cestari, M.M.; Ramsdorf, W.A.; Magalhães, V.F.; Wosiack, A.C.; Silva de Assis, H.C. Analyses of paralytic shellfish toxins and biomarkers in a Southern Brazilian reservoir. Toxicon 2010, 55, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Calado, S.L.M.; Wojciechowski, J.; Santos, G.S.; Magalhães, V.F.; Padial, A.A.; Cestari, M.M.; Assis, H.C.S. Neurotoxins in a water supply reservoir: An alert to environmental and human health. Toxicon 2017, 126, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Hegde, K.; Brar, S.K.; Cledon, M.; Kermanshahi pour, A. Physico-chemical treatment for the degradation of cyanotoxins with emphasis on drinking water treatment—How far have we come? J. Environ. Chem. Eng. 2018, 6, 5369–5388. [Google Scholar] [CrossRef]
- Ho, L.; Tang, T.; Monis, P.T.; Hoefel, D. Biodegradation of multiple cyanobacterial metabolites in drinking water supplies. Chemosphere 2012, 87, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.J.; Negri, A.P. Persistence and Degradation of cyanobacterial paralytic shellfish poisons (PSPs) in freshwaters. Water Res. 1997, 31, 525–533. [Google Scholar] [CrossRef]
- Kayal, N.; Newcombe, G.; Ho, L. Investigating the fate of saxitoxins in biologically active water treatment plant filters. Environ. Toxicol. 2008, 23, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.A.; Grant, F.; Ferguson, C.M.J.; Gallacher, S. Biotransformations of paralytic shellfish toxins by bacteria isolated from bivalve molluscs. Appl. Environ. Microbiol. 2001, 67, 2345–2353. [Google Scholar] [CrossRef] [Green Version]
- Burns, J.M.; Hall, S.; Ferry, J.L. The adsorption of saxitoxin to clays and sediments in fresh and saline waters. Water Res. 2009, 43, 1899–1904. [Google Scholar] [CrossRef]
- Chae, S.; Noeiaghaei, T.; Oh, Y.; Kim, I.S.; Park, J.S. Effective removal of emerging dissolved cyanotoxins from water using hybrid photocatalytic composites. Water Res. 2019, 149, 421–431. [Google Scholar] [CrossRef]
- Beamud, G.; Vico, P.; Haakonsson, S.; de la Escalera, G.M.; Piccini, C.; Brena, B.M.; Pirez, M.; Bonilla, S. Influence of UV-B radiation on the fitness and toxin expression of the cyanobacterium Cylindrospermopsis raciborskii. Hydrobiologia 2016, 763, 161–172. [Google Scholar] [CrossRef]
- Christensen, V.G.; Khan, E. Freshwater neurotoxins and concerns for human, animal, and ecosystem health: A review of anatoxin-a and saxitoxin. Sci. Total Environ. 2020, 736, 139515. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority—EFSA. Scientific opinion of the panel on contaminants in the food chain on a request from the European Commission on marine biotoxins in shellfish—Saxitoxin group. EFSA J. 2009, 1019, 1–76. [Google Scholar]
- Yu, H.; Lim, K.S.; Song, K.C.; Lee, K.J.; Lee, M.A.; Kim, J.H. Comparison of MBA and HPLC post-column oxidation methods for the quantification of paralytic shellfish poisoning toxins. Fish. Aquat. Sci. 2013, 16, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Malz, S.; Grell, M.N.; Thrane, C.; Maier, F.J.; Rosager, P.; Felk, A.; Albertsen, K.S.; Salomon, S.; Bohn, L.; Schäfer, W.; et al. Identification of a gene cluster responsible for the biosynthesis of aurofusarin in the Fusarium graminearum species complex. Fungal Genet. Biol. 2005, 42, 420–433. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization—WHO. Solar Disinfection Batch Systems (With or without Pre-Filtration And/Or Disinfection Addition); World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- He, Q.; Wang, W.; Xu, Q.; Liu, Z.; Teng, J.; Yan, H.; Liu, X. Microcystins in Water: Detection, Microbial Degradation Strategies, and Mechanisms. Int. J. Environ. Res. Public Health 2022, 19, 13175. [Google Scholar] [CrossRef]
- Kurtz, T.; Zeng, T.; Rosario-Ortiz, F.L. Photodegradation of cyanotoxins in surface waters. Water Res. 2021, 192, 116804. [Google Scholar] [CrossRef]
- Loftin, K.A.; Graham, J.L.; Hilborn, E.D.; Lehmann, S.C.; Meyer, M.T.; Dietze, J.E.; Griffith, C.B. Cyanotoxins in inland lakes of the United States: Occurrence and potential recreational health risks in the EPA National Lakes Assessment 2007. Harmful Algae 2016, 56, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Moraes, M.A.B.; Rodrigues, R.A.M.; Schlüter, L.; Podduturi, R.; Jørgensen, N.O.G.; Calijuri, M.C. Influence of environmental factors on occurrence of cyanobacteria and abundance of saxitoxin-producing cyanobacteria in a subtropical drinking water reservoir in Brazil. Water 2021, 13, 1716. [Google Scholar] [CrossRef]
- Vo Duy, S.; Munoz, G.; Dinh, Q.T.; Zhang, Y.; Simon, D.F.; Sauvé, S. Fast screening of saxitoxin, neosaxitoxin, and decarbamoyl analogues in fresh and brackish surface waters by on-line enrichment coupled to HILIC-HRMS. Talanta 2022, 241, 123267. [Google Scholar] [CrossRef]
- Al-Tebrineh, J.; Mihali, T.K.; Pomati, F.; Neilan, B.A. Detection of saxitoxin-producing cyanobacteria and Anabaena circinalis in environmental water blooms by quantitative PCR. Appl. Environ. Microbiol. 2010, 76, 7836–7842. [Google Scholar] [CrossRef] [PubMed]
- Manganelli, M. Blooms of toxic microorganisms in aquatic environments: Marine microalgae and freshwater cyanobacteria. A brief review with a particular focus on the Italian situation. Rend. Lincei 2016, 27, 135–143. [Google Scholar] [CrossRef]
- Miotto, M.C.; Costa, L.D.M.; Brentano, D.B.; Nader, C.; Souza, L.D.S.; Gressler, P.D.; Laudares-Silva, R.; Yunes, J.S.; Barufi, J.B.; Rörig, L.R. Ecophysiological characterization and toxin profile of two strains of Cylindrospermopsis raciborskii isolated from a subtropical lagoon in Southern Brazil. Hydrobiologia 2017, 802, 97–113. [Google Scholar] [CrossRef]
- Ramos, T.K.; Costa, L.D.F.; Yunes, J.S.; Resgalla, C., Jr.; Barufi, J.B.; Bastos, E.d.O.; Horta, P.A., Jr.; Rörig, L.R. Saxitoxins from the freshwater cyanobacterium Raphidiopsis raciborskii can contaminate marine mussels. Harmful Algae 2021, 103, 102004. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.; Hoefel, D.; Grasset, C.; Palazot, S.; Newcombe, G.; Saint, C.P.; Brookes, J.D. Removal of cyanobacterial metabolites through wastewater treatment plant filters. Water Sci. Technol. 2012, 65, 1244–1251. [Google Scholar] [CrossRef]
- Tang, T.; Hoefel, D.; Mosisch, T.; Ho, L. Assessing the fate and biodegradation of cyanobacterial metabolites in Australian waters. Water Pr. Technol. 2012, 7, wpt2012064. [Google Scholar] [CrossRef]
- Donovan, C.J.; Ku, J.C.; Quilliam, M.A.; Gill, T.A. Bacterial degradation of paralytic shellfish toxins. Toxicon 2008, 52, 91–100. [Google Scholar] [CrossRef]
- Donovan, C.J.; Garduño, R.A.; Kalmokoff, M.; Ku, J.C.; Quilliam, M.A.; Gill, T.A. Pseudoalteromonas bacteria are capable of degrading paralytic shellfish toxins. Appl. Environ. Microbiol. 2009, 75, 6919–6923. [Google Scholar] [CrossRef] [Green Version]
- Kurtz, T.D. The Photodegradation of saxitoxins in surface waters. MSc Thesis, University of Colorado, Boulder, CO, USA, 2021. [Google Scholar]
- Gill, T.A. Thermal degradation of paralytic shellfish poisoning toxins in scallop digestive glands. Food Res. Int. 1999, 32, 49–57. [Google Scholar] [CrossRef]
- Noyma, N.P.; Silva, T.P.; Chiarini-Garcia, H.; Amado, A.M.; Roland, F.; Melo, R.C.N. Potential effects of UV radiation on photosynthetic structures of the bloom-forming cyanobacterium Cylindrospermopsis raciborskii CYRF-01. Front. Microbiol. 2015, 6, 1202. [Google Scholar] [CrossRef] [Green Version]
- Cordeiro-Araújo, M.K.; Lorenzi, A.S.; Chia, M.A.; Mota, E.C.; Bittencourt-Oliveira, M.C. Insights into the impact of increasing temperature, light intensity, and UV-B exposure on the circadian rhythm of microcystin production and release, and the expression of mcy genes in the cyanobacterium Microcystis aeruginosa. J. Appl. Phycol. 2022, 34, 231–242. [Google Scholar] [CrossRef]
- Mullenders, L.H.F. Solar UV damage to cellular DNA: From mechanisms to biological effects. Photochem. Photobiol. Sci. 2018, 17, 1842–1852. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Sinha, R.P.; Moh, S.H.; Lee, T.K.; Kottuparambil, S.; Kim, Y.J.; Rhee, J.S.; Choi, E.M.; Brown, M.T.; Häder, D.P.; et al. Ultraviolet radiation and cyanobacteria. J. Photochem. Photobiol. B 2014, 141, 154–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theitler, D.J.; Nasser, A.; Gerchman, Y.; Kribus, A.; Mamane, H. Synergistic effect of heat and solar UV on DNA damage and water disinfection of E. coli and bacteriophage MS2. J. Water Health 2012, 10, 49–57. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Date | Detection of SXT in Water (W) and Phytoplankton (P) | Detection of SXT Analogs in Water | Degradation of SXT by Lake Water Bacteria | Degradation of SXT by Solar Exposure | Quantification of SXT Producers by qPCR |
|---|---|---|---|---|---|
| 21 June 2020 | HPLC (W + P) | Yes | |||
| 7 July 2020 | HPLC (W + P) | Yes | |||
| 28 July 2020 | HPLC (W + P) | HPLC | Yes | Yes | |
| 17 August 2020 | HPLC (W + P) | Yes | Yes | ||
| 15 September 2020 | HPLC (W + P) | HPLC | Yes | Yes | |
| 11 October 2020 | HPLC (W + P) | HPLC | Yes | ||
| 6 June 2021 | ELISA (W) | Yes | Yes | ||
| 4 July 2021 | ELISA (W) | Yes | Yes | ||
| 13 September 2021 | ELISA (W) | Yes | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jørgensen, N.O.G.; Podduturi, R.; Michelsen, C.F.; Jepsen, T.; Moraes, M.d.A.B. Fate of Saxitoxins in Lake Water: Preliminary Testing of Degradation by Microbes and Sunlight. Water 2022, 14, 3556. https://doi.org/10.3390/w14213556
Jørgensen NOG, Podduturi R, Michelsen CF, Jepsen T, Moraes MdAB. Fate of Saxitoxins in Lake Water: Preliminary Testing of Degradation by Microbes and Sunlight. Water. 2022; 14(21):3556. https://doi.org/10.3390/w14213556
Chicago/Turabian StyleJørgensen, Niels O. G., Raju Podduturi, Charlotte Frydenlund Michelsen, Thea Jepsen, and Munique de Almeida Bispo Moraes. 2022. "Fate of Saxitoxins in Lake Water: Preliminary Testing of Degradation by Microbes and Sunlight" Water 14, no. 21: 3556. https://doi.org/10.3390/w14213556
APA StyleJørgensen, N. O. G., Podduturi, R., Michelsen, C. F., Jepsen, T., & Moraes, M. d. A. B. (2022). Fate of Saxitoxins in Lake Water: Preliminary Testing of Degradation by Microbes and Sunlight. Water, 14(21), 3556. https://doi.org/10.3390/w14213556