
Toxic Effects and Mechanism of Pefloxacin on the Early Life Stage of Zebrafish (Danio rerio)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental
2.1. Apparatus and Materials
2.2. Zebrafish Husbandry and Embryo Exposure
2.3. PFLX Determination
2.4. The Interaction between PFLX and Embryonic Membrane
2.5. Embryo Behavior and Toxicity Experiment
2.6. Determination of Exposure Concentration
2.7. Larva Behavior and Toxicity Experiment
2.8. Data Analysis
3. Results and Discussion
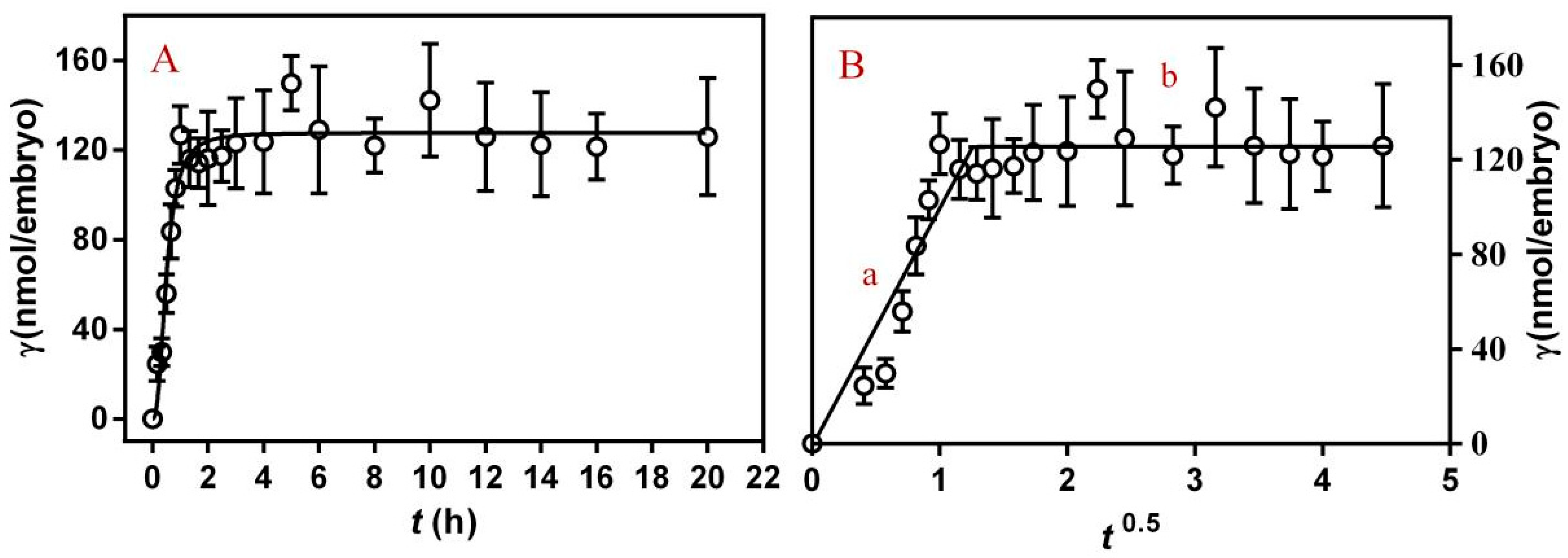
3.1. The Interaction between PFLX and Embryonic Membrane
3.2. Effects of PFLX Exposure on Embryonic Membranes
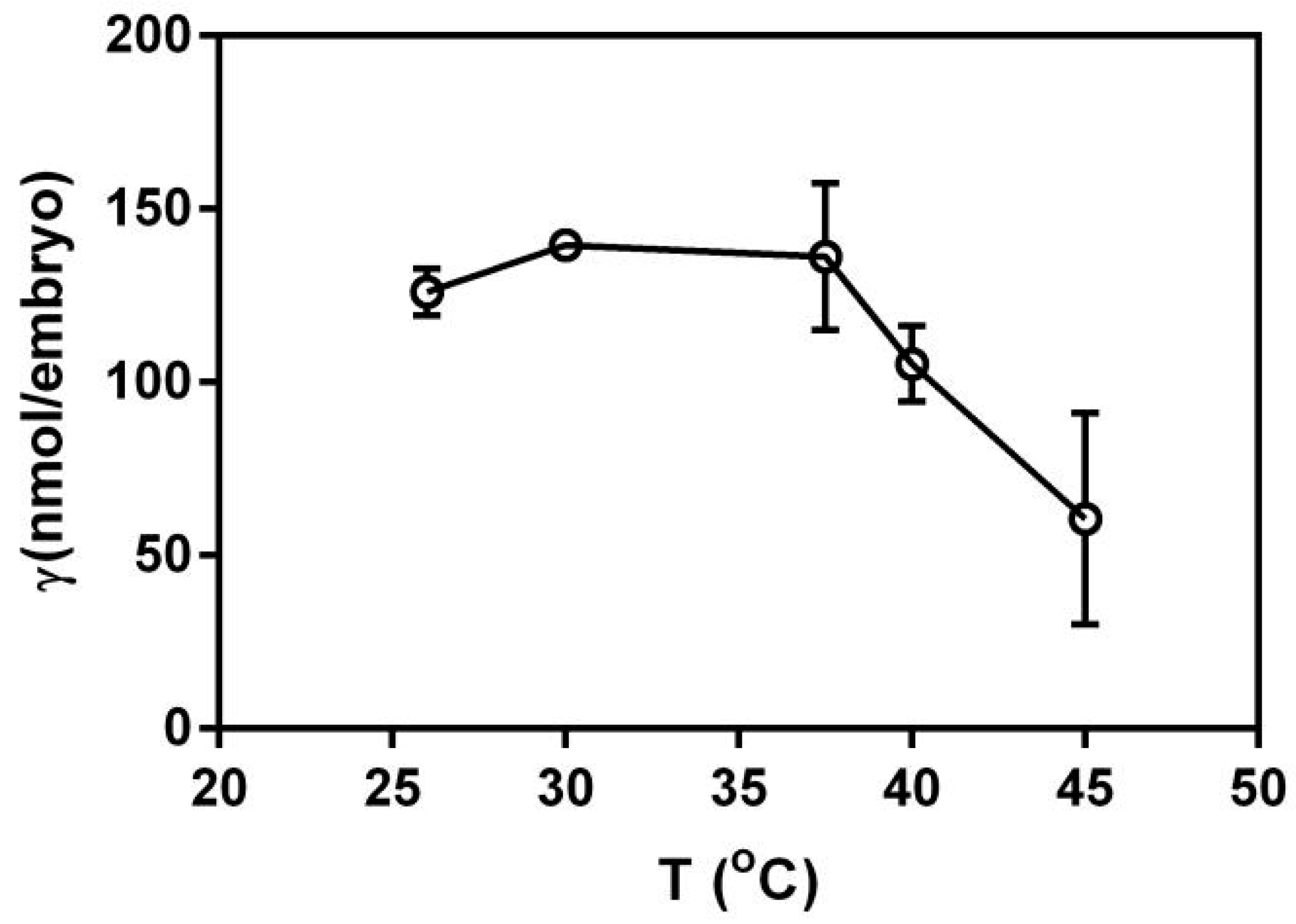
3.3. Effect of Temperature on the Action of PFLX and Embryo
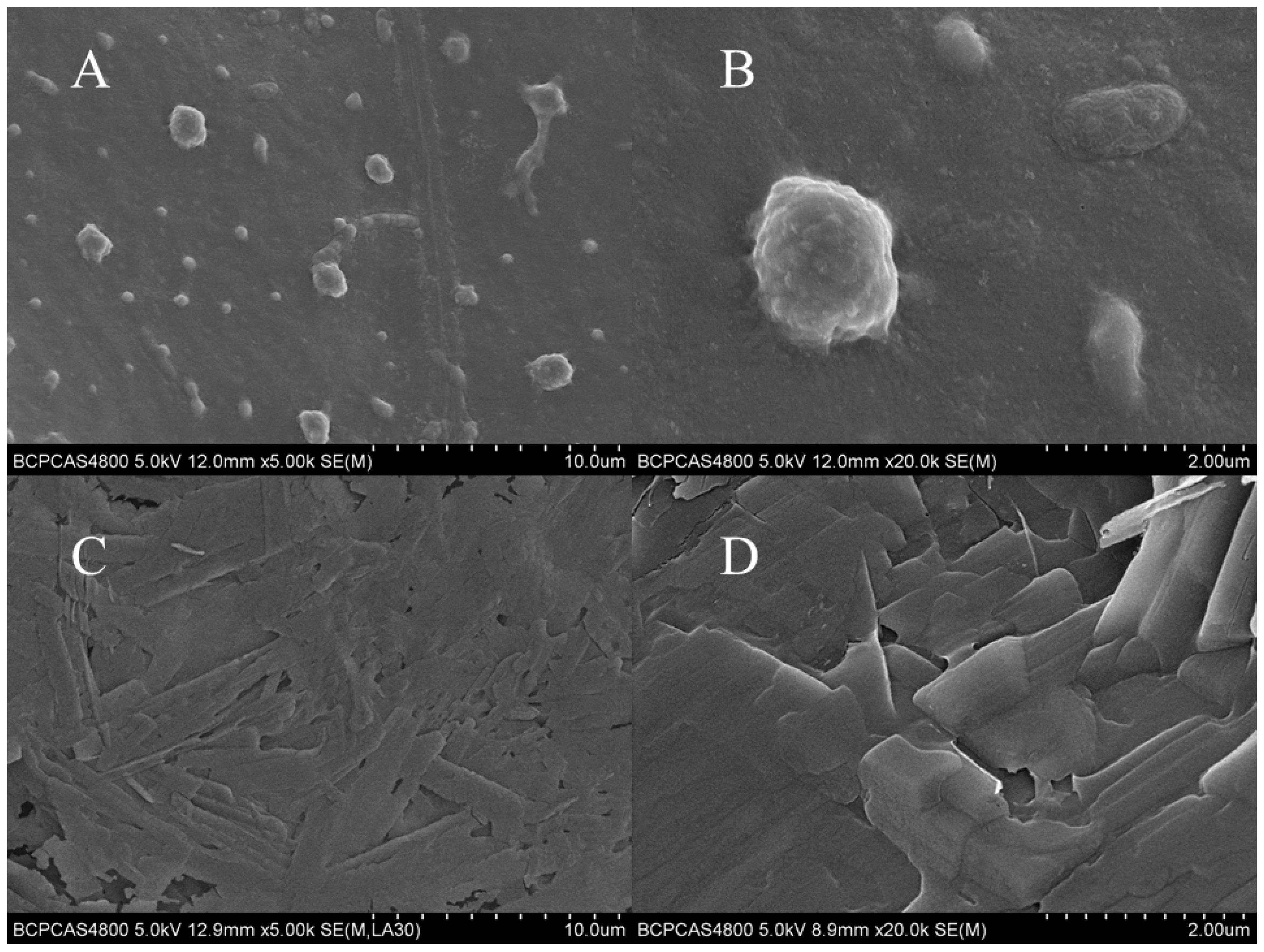
3.4. SEM Analysis
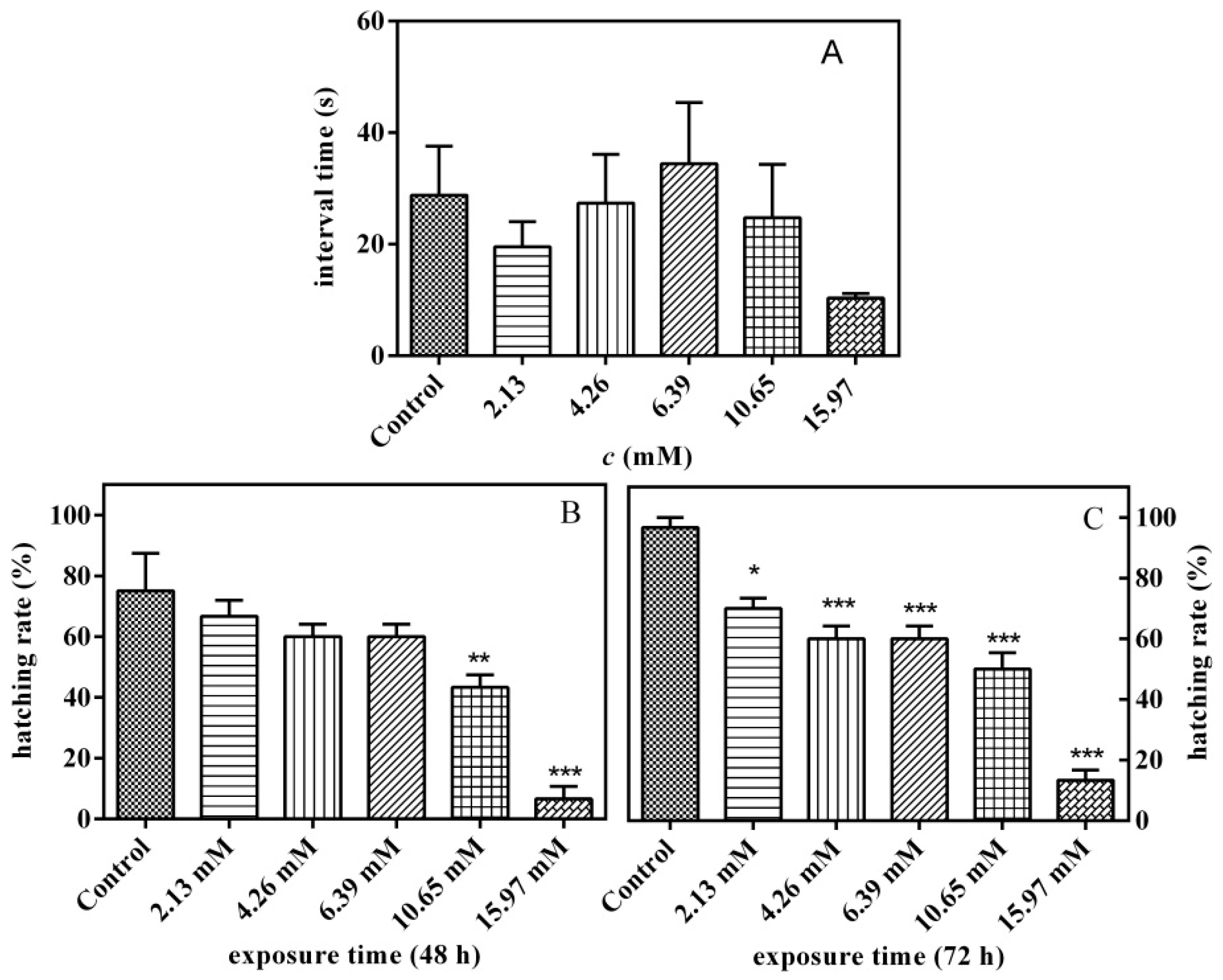
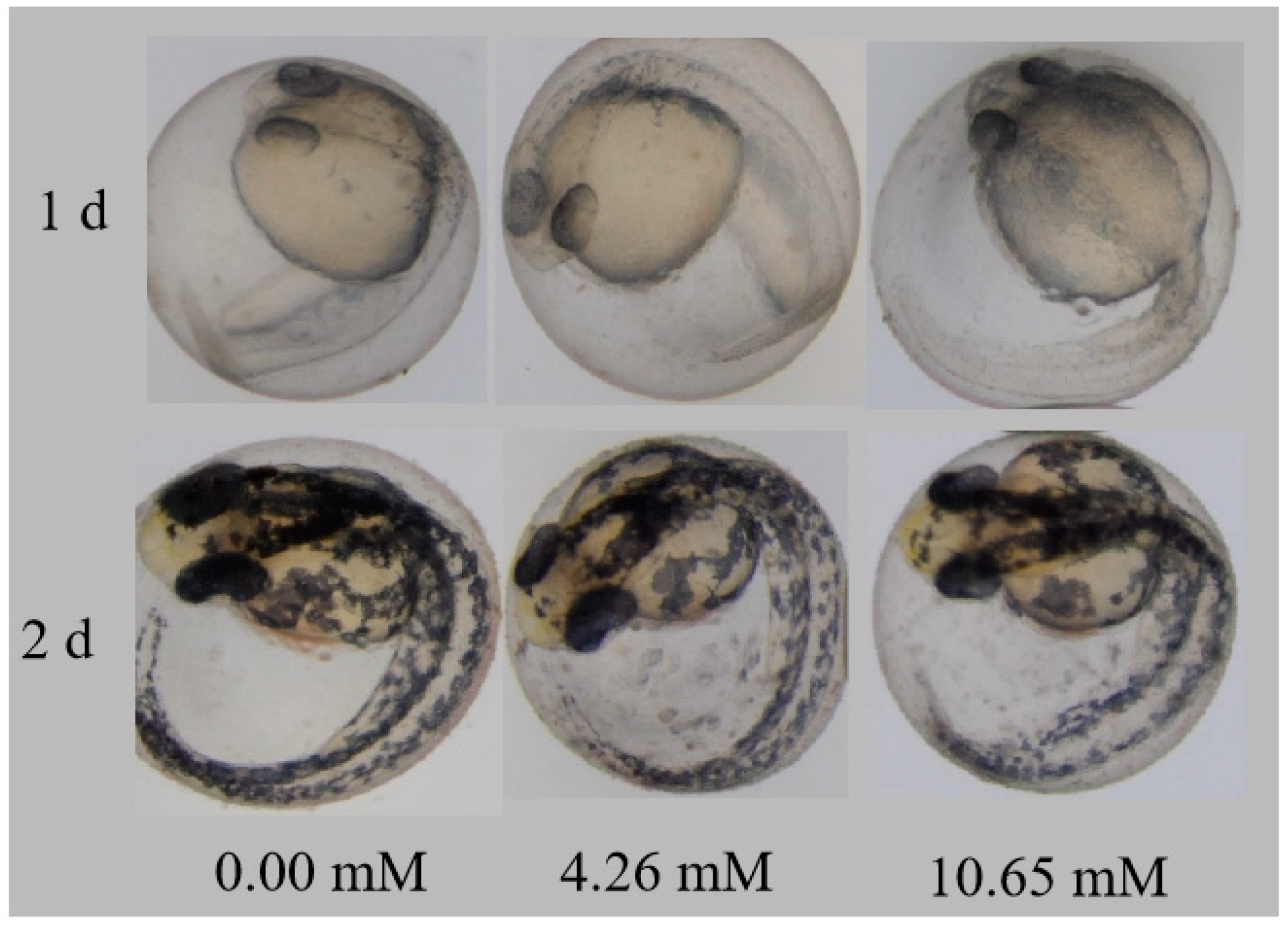
3.5. Effects of PFLX Exposure on Embryonic Autonomic Movement and Hatchability
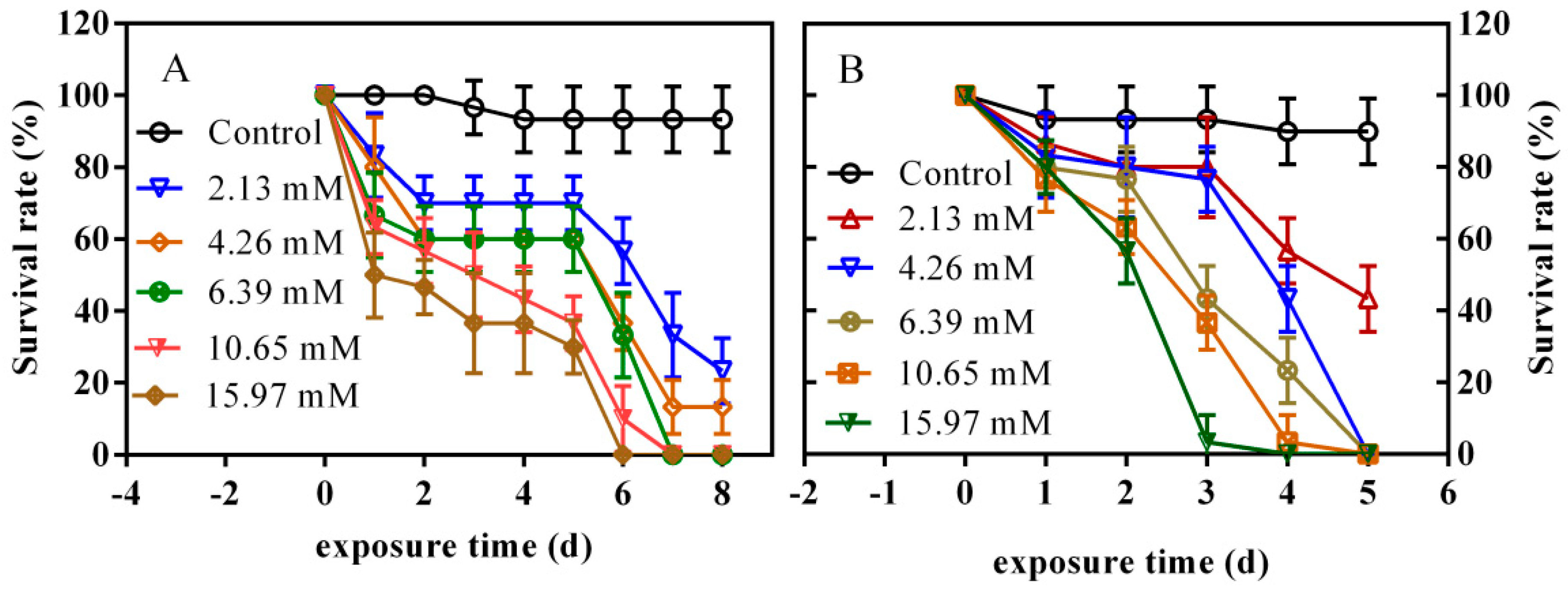
3.6. Effects of PFLX Exposure on the Survival Rate of Zebrafish Embryos and Larvae
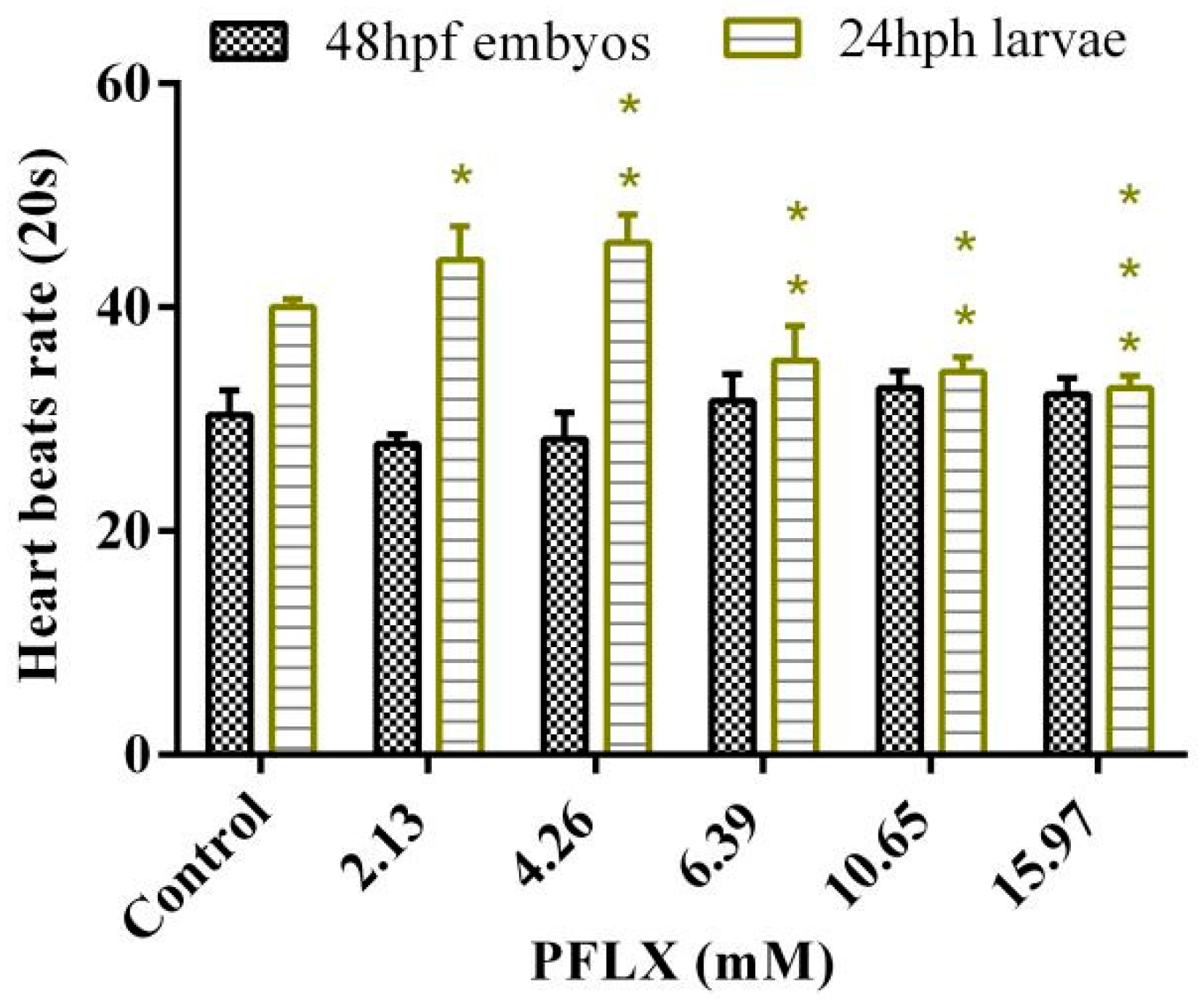
3.7. Effects of PFLX Exposure on the Heart Rate of Zebrafish Embryo and Larva
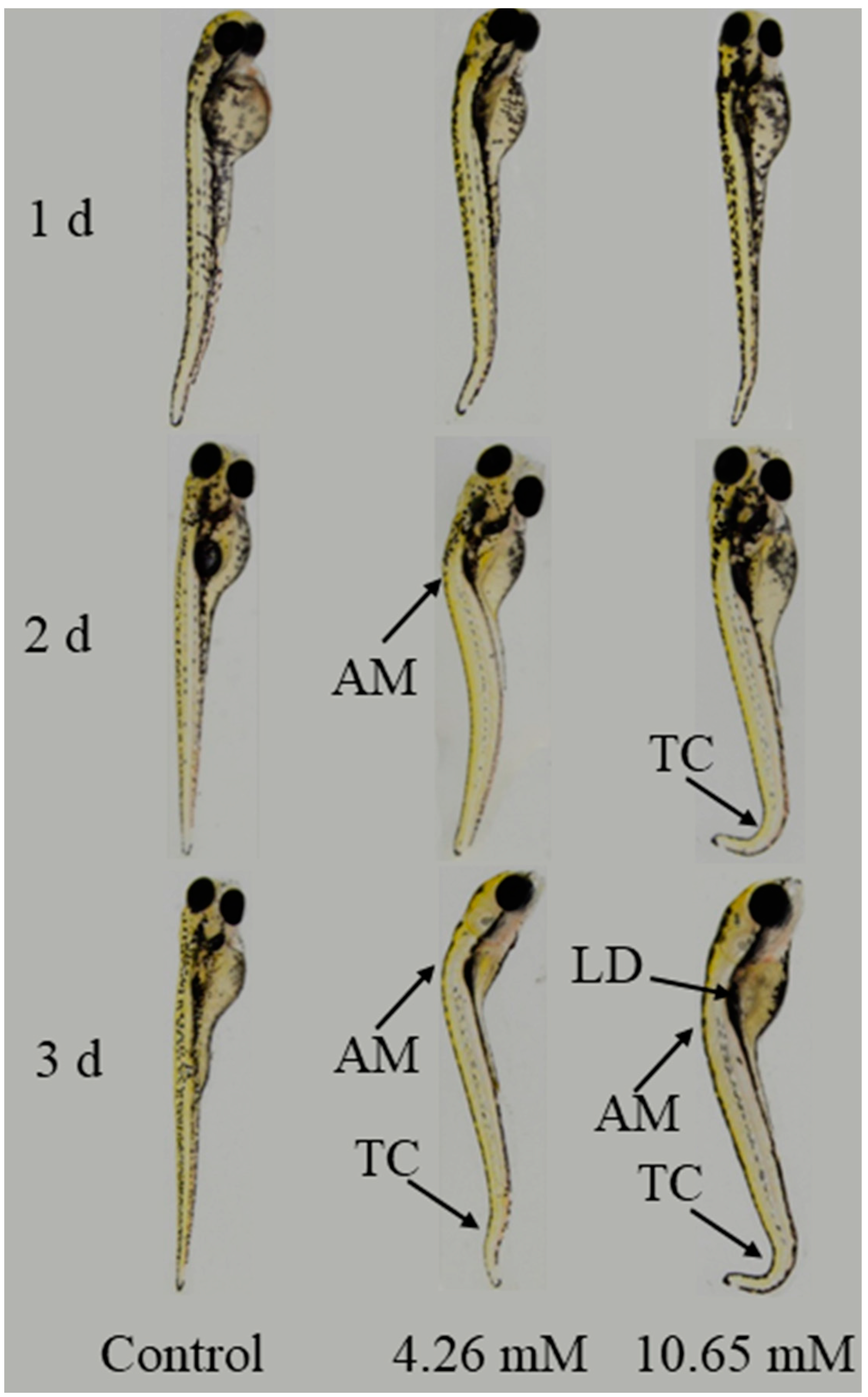
3.8. Teratogenic Effect of PFLX on Zebrafish Embryos and Larvae
3.9. Correlation Analysis of Membrane Effect and Toxic Effect of PFLX
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Liu, F.-F.; Wang, S.-C.; Huang, T.-Y.; Li, M.-R.; Zhu, Z.-L.; Liu, G.-Z. Sorption of fluoroquinolones to nanoplastics as affected by surface functionalization and solution chemistry. Environ. Pollut. 2020, 262, 114347. [Google Scholar] [CrossRef] [PubMed]
- Von Ah, S.; Stephan, R.; Zurfluh, K.; Sidler, X.; Kümmerlen, D. Occurrence of quinolone-resistant Escherichia coli in environmental samples from a sow pool system in Switzerland. Schweiz. Arch. Tierh. 2019, 161, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Kindle, P.; Zurfluh, K.; Nüesch-Inderbinen, M.; Von Ah, S.; Sidler, X.; Stephan, R.; Kümmerlen, D. Phenotypic and genotypic characteristics of Escherichia coli with non-susceptibility to quinolones isolated from environmental samples on pig farms. Porc. Health Manag. 2019, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Song, W.; Lin, H.; Wang, W.; Du, L.; Xing, W. Antibiotics and antibiotic resistance genes in global lakes: A review and meta-analysis. Environ. Int. 2018, 116, 60–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Shi, W.; Liu, W.; Li, H.; Zhang, W.; Hu, J.; Ke, Y.; Sun, W.; Ni, J. A duodecennial national synthesis of antibiotics in China’s major rivers and seas (2005–2016). Sci. Total Environ. 2018, 615, 906–917. [Google Scholar] [CrossRef]
- Du, J.; Zhao, H.; Wang, Y.; Xie, H.; Zhu, M.; Chen, J. Presence and environmental risk assessment of selected antibiotics in coastal water adjacent to mariculture areas in the Bohai Sea. Ecotoxicol. Environ. Saf. 2019, 177, 117–123. [Google Scholar] [CrossRef]
- Dorival-García, N.; Zafra-Gómez, A.; Cantarero, S.; Navalón, A.; Vílchez, J. Simultaneous determination of 13 quinolone antibiotic derivatives in wastewater samples using solid-phase extraction and ultra performance liquid chromatography–tandem mass spectrometry. Microchem. J. 2013, 106, 323–333. [Google Scholar] [CrossRef]
- Teglia, C.M.; Perez, F.A.; Michlig, N.; Repetti, M.R.; Goicoechea, H.C.; Culzoni, M.J. Occurrence, Distribution, and Ecological Risk of Fluoroquinolones in Rivers and Wastewaters. Environ. Toxicol. Chem. 2019, 38, 2305–2313. [Google Scholar] [CrossRef]
- Wang, Z.; Du, Y.; Yang, C.; Liu, X.; Zhang, J.; Li, E.; Zhang, Q.; Wang, X. Occurrence and ecological hazard assessment of selected antibiotics in the surface waters in and around Lake Honghu, China. Sci. Total Environ. 2017, 609, 1423–1432. [Google Scholar] [CrossRef]
- Rosendahl, I.; Siemens, J.; Kindler, R.; Groeneweg, J.; Zimmermann, J.; Czerwinski, S.; Lamshöft, M.; Laabs, V.; Wilke, B.-M.; Vereecken, H.; et al. Persistence of the Fluoroquinolone Antibiotic Difloxacin in Soil and Lacking Effects on Nitrogen Turnover. J. Environ. Qual. 2012, 41, 1275–1283. [Google Scholar] [CrossRef]
- Gatti, M.; Bianchin, M.; Raschi, E.; De Ponti, F. Assessing the association between fluoroquinolones and emerging adverse drug reactions raised by regulatory agencies: An umbrella review. Eur. J. Intern. Med. 2020, 75, 60–70. [Google Scholar] [CrossRef]
- Wang, D.; Ning, Q.; Dong, J.; Brooks, B.W.; You, J. Predicting mixture toxicity and antibiotic resistance of fluoroquinolones and their photodegradation products in Escherichia coli. Environ. Pollut. 2020, 262, 114275. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.M. Ocular toxicity of fluoroquinolones. Clin. Exp. Ophthalmol. 2007, 35, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Chen, J.; Li, X.; Zhang, S.; Yao, H.; Peijnenburg, W.J. Effects of lomefloxacin on survival, growth and reproduction of Daphnia magna under simulated sunlight radiation. Ecotoxicol. Environ. Saf. 2018, 166, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Luo, Y.; Wu, L.; Huang, Y.; Christie, P. Residues and potential ecological risks of veterinary antibiotics in manures and composts associated with protected vegetable farming. Environ. Sci. Pollut. Res. 2015, 22, 5908–5918. [Google Scholar] [CrossRef]
- Lu, T.; Zhu, Y.; Ke, M.; Peijnenburg, W.; Zhang, M.; Wang, T.; Chen, J.; Qian, H. Evaluation of the taxonomic and functional variation of freshwater plankton communities induced by trace amounts of the antibiotic ciprofloxacin. Environ. Int. 2019, 126, 268–278. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Yin, X.; Shi, M.; Dahlgren, R.A.; Wang, H. Toxicity Assessment of Combined Fluoroquinolone and Tetracycline Exposure in Zebrafish (Danio rerio). Environ. Toxicol. 2016, 31, 736–750. [Google Scholar] [CrossRef] [Green Version]
- Shen, R.; Yu, Y.; Lan, R.; Yu, R.; Yuan, Z.; Xia, Z. The cardiovascular toxicity induced by high doses of gatifloxacin and ciprofloxacin in zebrafish. Environ. Pollut. 2019, 254, 112861. [Google Scholar] [CrossRef]
- Peltzer, P.M.; Lajmanovich, R.C.; Attademo, A.M.; Junges, C.M.; Teglia, C.M.; Martinuzzi, C.; Curi, L.; Culzoni, M.J.; Goicoechea, H.C. Ecotoxicity of veterinary enrofloxacin and ciprofloxacin antibiotics on anuran amphibian larvae. Environ. Toxicol. Pharmacol. 2017, 51, 114–123. [Google Scholar] [CrossRef]
- Zhao, J.; Guo, X.; He, Q.; Song, Q.; Wu, F.; Zhang, C. Solvothermal synthesis of InNbO4 cubes for efficient degradation of pefloxacin. Spectrochim. Acta A 2020, 234, 118247. [Google Scholar] [CrossRef]
- Lee, H.-J.; Kim, K.Y.; Hamm, S.-Y.; Kim, M.; Kim, H.K.; Oh, J.-E. Occurrence and distribution of pharmaceutical and personal care products, artificial sweeteners, and pesticides in groundwater from an agricultural area in Korea. Sci. Total Environ. 2019, 659, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, L.; Liu, X.; Ding, J. Ranking and prioritizing pharmaceuticals in the aquatic environment of China. Sci. Total Environ. 2019, 658, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Sha, L.; Tang, X.; Liu, D.; Xu, Y.; Ding, Y.; Ding, F. Detection and Quantitation of Lomefloxacin and Pefloxacin Residues in the Organ Tissues and Eggs of Laying Hens. J. Food Prot. 2018, 81, 810–814. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; He, K.; Zhang, D.; Huang, Z. Production and characterization of a monoclonal antibody for Pefloxacin and mechanism study of antibody recognition. Biosci. Biotechnol. Biochem. 2019, 83, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Minhaz, A.; Shah, M.R.; Ain, N.U.; Khan, A.A.; Shah, K.; Ullah, S.; Ishaq, M. Highly selective and sensitive chemosensor for detection of Pefloxacin in tap water based on click generated triazole. Microchem. J. 2019, 146, 332–338. [Google Scholar] [CrossRef]
- Qu, S.; Wei, J.; Wang, Q.; Li, Y.; Jin, X.; Chaib, L. Oral toxicity of pefloxacin, norfloxacin, ofloxacin and ciprofloxacin: Comparison of biomechanical and histopathological effects on Achilles tendon in rats. J. Toxicol. Sci. 2011, 36, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; van Mil, H.G.; Richardson, M.K. Large-scale assessment of the zebrafish embryo as a possible predictive model in toxicity testing. PLoS ONE 2011, 6, e21076. [Google Scholar] [CrossRef] [Green Version]
- Di Paolo, C.; Seiler, T.-B.; Keiter, S.; Hu, M.; Muz, M.; Brack, W.; Hollert, H. The value of zebrafish as an integrative model in effect-directed analysis—A review. Environ. Sci. Eur. 2015, 27, 8. [Google Scholar] [CrossRef] [Green Version]
- Marques, I.J.; Lupi, E.; Mercader, N. Model systems for regeneration: Zebrafish. Development 2019, 146, dev167692. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Xiao, H.; Di Paolo, C.; Deutschmann, B.; Brack, W.; Hollert, H.; Seiler, T.B. Integrated zebrafish-based tests as an investigation strategy for water quality assessment. Water Res. 2019, 150, 252–260. [Google Scholar] [CrossRef]
- Babić, S.; Barišić, J.; Stipaničev, D.; Repec, S.; Lovrić, M.; Malev, O.; Martinovic-Weigelt, D.; Čož-Rakovac, R.; Klobučar, G. Assessment of river sediment toxicity: Combining empirical zebrafish embryotoxicity testing with in silico toxicity characterization. Sci. Total Environ. 2018, 643, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Stainier, D.Y. Zebrafish genetics and vertebrate heart formation. Nat. Rev. Genet. 2001, 2, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Gao, N.Y.; Gao, H.W. Transmembrane distribution of kanamycin and chloramphenicol: Insights into the cytotoxicity of antibacterial drugs. Mol. Biosyst. 2010, 6, 1901–1910. [Google Scholar] [CrossRef]
- Yu, Y.; Hu, Z.; Wang, Y.; Gao, H. Magnetic SN-functionalized diatomite for effective removals of phenols. Int. J. Miner. Process. 2017, 162, 1–5. [Google Scholar] [CrossRef]
- Xiao, C.; Han, Y.; Liu, Y.; Zhang, J.; Hu, C.-Q. Relationship Between Fluoroquinolone Structure and Neurotoxicity Revealed by Zebrafish Neurobehavior. Chem. Res. Toxicol. 2018, 31, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Asadipour, A.; Moshafi, M.H.; Khosravani, L.; Moghimi, S.; Amou, E.; Firoozpour, L.; Ilbeigi, G.; Beiki, K.; Soleimani, E.; Foroumadi, A. N-substituted piperazinyl sarafloxacin derivatives: Synthesis and in vitro antibacterial evaluation. DARU 2018, 26, 199–207. [Google Scholar] [CrossRef]
- Zhang, Z.; Xie, X.; Yu, Z.; Cheng, H. Influence of chemical speciation on photochemical transformation of three fluoroquinolones (FQs) in water: Kinetics, mechanism, and toxicity of photolysis products. Water Res. 2019, 148, 19–29. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, R.; Yu, Y.; Liu, Q.; Wang, R.; Lan, R.; Feng, H.; Yu, R.; Jin, L.; Li, Y. Toxic Effects and Mechanism of Pefloxacin on the Early Life Stage of Zebrafish (Danio rerio). Water 2022, 14, 3857. https://doi.org/10.3390/w14233857
Shen R, Yu Y, Liu Q, Wang R, Lan R, Feng H, Yu R, Jin L, Li Y. Toxic Effects and Mechanism of Pefloxacin on the Early Life Stage of Zebrafish (Danio rerio). Water. 2022; 14(23):3857. https://doi.org/10.3390/w14233857
Chicago/Turabian StyleShen, Rong, Yichang Yu, Qinfang Liu, Ruili Wang, Rong Lan, Hui Feng, Ran Yu, Lihua Jin, and Ye Li. 2022. "Toxic Effects and Mechanism of Pefloxacin on the Early Life Stage of Zebrafish (Danio rerio)" Water 14, no. 23: 3857. https://doi.org/10.3390/w14233857
APA StyleShen, R., Yu, Y., Liu, Q., Wang, R., Lan, R., Feng, H., Yu, R., Jin, L., & Li, Y. (2022). Toxic Effects and Mechanism of Pefloxacin on the Early Life Stage of Zebrafish (Danio rerio). Water, 14(23), 3857. https://doi.org/10.3390/w14233857