1. Introduction
One of the main issues preventing the efficient monitoring of the state of aquatic ecosystems and combating the consequences of their pollution is the prompt detection of pollutants making it possible to estimate their sublethal exposure [
1].
The early detection of environmental ill-being is particularly important in hazardous areas such as nuclear power plants, oil platforms, gas and oil pipelines and waste disposal zones of industrial plants and treatment facilities. Currently, there is a growing interest (due to increasing relevance) in the problem of detecting the impact of substances with weak toxicity, which constantly enter water bodies. Until recently, “biologically neutral” plastics in food packaging and agricultural raw materials have been considered the typical representatives of this class of substances. However, recent studies showed that contamination with micro- (>100 nm and <5 mm) or nanoplastics (<100 nm), which may be formed by the fragmentation of larger pieces of plastic, presents a global environmental problem of the marine ecosystem [
2,
3,
4].
Due to its small size, close to that of microalgae, microplastics can easily enter a wide range of euryhaline crustaceans [
4,
5,
6,
7] and be transmitted via the food chain [
8,
9].
The correlation between the abundance of microplastics and total number of zooplankton revealed by the authors in zooplankton samples taken from the water column makes it possible to better understand the role of these pollutants in marine ecosystems and may indicate their potential impact on various taxa and inclusion in the marine food chain [
9,
10].
There are laboratory data on
Mysidacea,
Copepods, Cladocerae,
Rotiferae, polychaete larvae and infusoria of the Baltic Sea capable of swallowing the polystyrene microspheres of 10 μm in size and transferring plastic microparticles through plankton organisms from one trophic level (mesozooplankton) to a higher level (macrozooplankton) [
8].
Laboratory studies on freshwater (
Daphnia pulex) and euryhaline (
Artemia franciscana and
Artemia salina) organisms showed that exposure to model micro- and nanospheres (mainly polystyrene) affected the growth, development and reproduction of
D. pulex crustaceans [
11], changed the sex ratio of newborns, inhibited the body length, reduced the growth rate and decreased the egg laying rate [
12]. Furthermore, the accumulation of microplastics in larvae leading to multiple molts, adsorption of particles on the surface of sensory antennas and appendages prevented the larval mobility of
A. franciscana [
13,
14] and influenced the eating behavior and life cycle of
A. salina [
15].
Recent studies showed that the indicators of the behavioral response (speed, swimming path, etc.) are more sensitive to most pollutants than survival, growth, development and reproduction [
16,
17,
18,
19]. Additionally, for most pollutants, the values of EC
50 in terms of behavioral responses [
20,
21] are much lower than those of LC
50 based on the death rate of hydrobionts, their reproduction rate and the quality of offspring [
22,
23,
24,
25], which are obtained using standard procedures [
26,
27]. Moreover, data on the parameters of behavioral responses (speed, swimming path, etc.) can be obtained in a shorter time.
The phototropic response of hydrobionts, i.e., phototaxis, is especially evident in diel vertical migration and is of particular interest among other behavioral responses of zooplankton.
However, the available data on the phototropic response of individual representatives of aquatic organisms is quite contradictory. For example, with the increase in light intensity,
D. magna try to escape from light that in natural conditions allows them to avoid attack from predatory fish in the daytime [
1]. Other authors [
28,
29,
30,
31] observed that the crustaceans gathered in the light zone and that positive phototaxis was suppressed in the presence of toxicants.
The study revealed a certain dependence of the behavior on wavelength and light intensity (up to the sign inversion of phototaxis) [
1,
32,
33]. It also showed the behavioral [
34] and gender [
35] differences of hydrobionts at both interspecific and intraspecific [
34] levels.
However, the use of behavioral responses of hydrobionts to assess environmental risks is currently quite difficult due to the lack of optimized and standard methods [
36]. Several automated high-performance behavioral analyzers have been recently created to handle different types of crustaceans. Microfluidic cameras were developed to study the behavioral responses of
Allorchestes Compressa and
A. franciscana [
17,
18], which make it possible to record changes in the swimming path in the presence of behavior-modifying compounds. Major efforts in this direction were also made by bbe Moldaenke GmbH (DaphTox II, bbe Moldaenke GmbH, Germany, Schwentinental). However, the current methods are based on the use of stationary devices with a limited set of test organisms, which significantly reduces the ability to monitor the state of ecosystems in dynamic and in situ contexts.
Therefore, there is an urgent need to register the behavioral responses of autochthonous organisms directly within the habitat using submersible tools. The significant breakthrough and transition to studies in natural aquatic systems is connected to the development of in situ holography. Submersible holographic cameras register information on all particles in the controlled volume on one hologram. The use of the time series of holograms makes it possible to build a trajectory and study the motion pattern of each particle [
37,
38,
39].
The in situ registration of behavioral responses led to the understanding that the environmental safety of pollutants, including micro- and nanoplastics needs to be assessed according to the response of biosystems to these pollutants in the habitat. It is worth noting that without the proper study of the methods of detection, identification, separation and definition of concentration (in aqueous media), the mere fact of detection of micro- and nanoplastics will not make it possible to predict the degree of their threat to aquatic ecosystems. In this case, it is advisable to use the recent safety assessment principles with regard to nanomaterials throughout their entire life cycle (from macro- to nanoforms) [
23,
40].
Furthermore, the prompt diagnostics of the altering exposure should be based on the registration of collective phenomena within the autochthonous biotic community. This allows not only controlling the most mobile (in terms of response speed) behavioral processes, but also obtaining a statistically sufficient amount of information on the plankton ensemble to forecast the most probable scenario.
At the same time, the problem of reliable identification of responses to weak exposures is largely associated with variability in the state of certain individuals within the studied population. Accordingly, the threshold and intensity of response to the test stimulus will vary between individuals resulting in large dispersion in the response array and reduction in the significance level of the received response.
Earlier we demonstrated the advantages of using the paired photostimulation of the behavioral response on three species of freshwater zooplankton:
Cladocera (
D. magna S. and
D. pulex), as well as
Copepoda (
Cyclops vicinus). The advanced use of this method shall be confirmed for saltwater reservoirs according to the response of an ensemble of aboriginal, less sensitive euryhaline organisms [
41].
The purpose of this study was to identify the general patterns of change in the phototropic response of euryhaline zooplankton during the contamination of the habitat and reveal the possibility of using paired photostimulation to detect the contamination of water areas with micro- and nanoplastics.
2. Materials and Methods
2.1. Test-Organisms
The studies were performed in laboratory conditions on A. salina and M. salina—a species of crustaceans from the Branchiopoda class. The use of saltwater crustaceans can be explained by the need to expand the scope of the paired photostimulation to saltwater reservoirs.
A. salina cysts were obtained from Neva Tropik LLC (St. Petersburg, Russia). For all experiments, 0.1 g of dry cysts washed with distilled water were placed in an aerated water tank with 0.3 dm
3 of artificial seawater. Artificial seawater was prepared according to [
42] with 35 g/dm
3 salinity and pH = 8.0–8.3. At constant aeration, daylight and temperature (22 ± 2 °C), the nauplii began to hatch after 24 ± 1 h. Time-synchronized nauplii were moved to clean artificial seawater.
M. salina females were taken from a mixture of 3 cultures (M. salina, Apocyclops panamensis, Brachionus plicatus)—live food supplied by Aquacopa GmbH. (Jabel, Germany) and kept in the laboratory in filtered (0.45 µm) artificial seawater with 35 g/dm3 salinity. Newborn crustaceans were placed in a separate tank with clean seawater. The culture of adults and synchronized culture of M. salina were kept under standard, controlled conditions: light regime—10 h light/14 h dark; temperature 23 ± 1 °C. Food was introduced daily in the form of a suspension of Chlorella vulgaris or Tetraselmis suecica algae (1.5 × 106 cells/dm3). Once a week, a suspension of baker’s yeast was added at a rate of 0.01 g/dm3. During cultivation, the density of M. salina was maintained at the level of 50 ind/dm3.
The experiments were conducted under controlled conditions according to the procedure [
43].
2.2. Used Pollutants
Substances with pronounced toxicity—a standard model toxicant K2Cr2O7 (Merck KGaA, Germany, Darmstadt)—and substances made of relatively bioinert materials—nanoplastics (nPl) and microplastics (mPl)—were used in the study as pollutants.
Monodispersed fluorescent microspheres of polystyrene SEQ-G-001 (Tianjin Saierqun Technology Co., Ltd, China, Tianjin) with a particle size of 100 nm, maximum fluorescence excitation wavelength of 488 nm, maximum fluorescence radiation wavelength of 518 nm, and concentration of 10 mg/cm3 were used as nanoplastics. We found no acute toxicity up to a concentration of 10 mg/L (LC50 = 19 mg/L) in the standard input control test for crustacean mortality.
The nPl suspension of the desired concentration was prepared by the sonification of an aliquot of the initial suspension in 0.5 dm3 of artificial seawater for 10 min at an ultrasonic frequency of 25 kHz and a power of 30 W/dm3.
Microplastics were prepared directly in the laboratory from woven polypropylene bag fibers (MIRPACK, Russia, Zhukov) aged in natural conditions for 12 years. Preliminarily washed with distilled water and dried, bag fibers were cut into about 1 mm particles. One hundred milligrams of plastic were transferred to a porcelain mortar and ground with 20 g of quartz sand for 10 min, adding 25 cm3 of distilled water dropwise. The suspension was transferred quantitatively to a 1.0 dm3 flask with the further addition of distilled water reaching 0.5 dm3. After shaking for 10 min, the resulting contents were centrifuged in a laboratory centrifuge (Eppendorf 5804, Germany, Hamburg) at 3000 rpm for 10 min for the sand particles to settle. The supernatant was filtered through a white tape filter.
The concentration and size of microparticles were determined according to images of the suspension droplets obtained by a confocal microscope when excited with a 405 nm laser. The counting of particles from another material was excluded due to the autoluminescence of microplastic particles. The particles were counted in 850 μm × 850 μm × 2.7 μm layers with a volume of 1.9510–6 mL (
Figure 1). The size of 1 pixel at this resolution was 0.83∙μm × 0.83 μm. The observed microplastic particles were visualized as compact groups with a 2–5 pixel size. The number of particles with a size of ≤10 pixels (circle diameter with an equivalent area of less than 4 microns) and >10 pixels was then calculated. The fluctuation in the correlation of these groups of particles in different samples is shown in
Figure 1b. The content of microparticles with a size of less than 10 pixels averaged over 10 samples was 15.4 ± 1.4 pcs., and the microparticles of a larger size—6.7 ± 1.3 pcs. (70 and 30%, respectively).
Thus, the microplastic concentration in the stock suspension was ~107 particles/dm3 (200 mg/dm3).
Solutions or dispersed suspensions of pollutants in artificial seawater were added in the amount of 0.5 dm3.
The cumulative exposure dose (CD causing a significant change in the phototropic response) was calculated as the product of the pollutant concentration Cpoll by the duration of its action T.
CD (mg/dm3∙h) = Cpoll (mg/dm3)∙T(h).
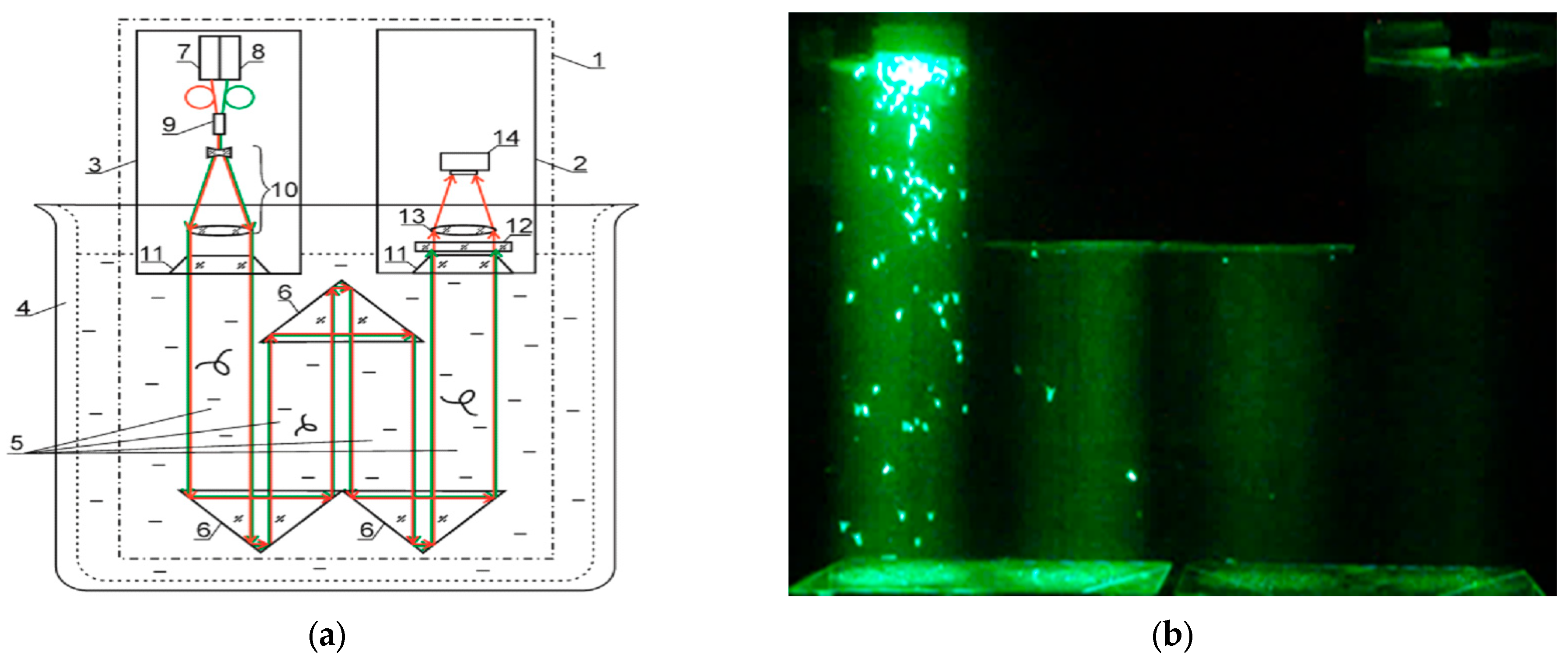
2.3. Digital Holographic Camera
The design of a submersible digital holographic camera (DHC) is described in detail in [
30]. During laboratory experiments, the DHC was placed in a 90 dm
3 tank filled with 50 dm
3 of artificial seawater [
42], which ensured that the DHC optical part was in the aqueous medium. The volume of the illuminated and controlled aqueous medium was determined by a beam of laser radiation formed by the DHC mirror-prism system to produce 0.5 dm
3 (
Figure 2).
The images of 400 layers of the studied volume with a thickness of 1.7 mm were sequentially and automatically reconstructed from each registered digital hologram of the measured volume, thusly calculating the number of crustaceans layer-by-layer using the DHC.
2.4. Paired Photostimulation of Behavioral Response to Assess the Responsiveness of Planktonic Communities
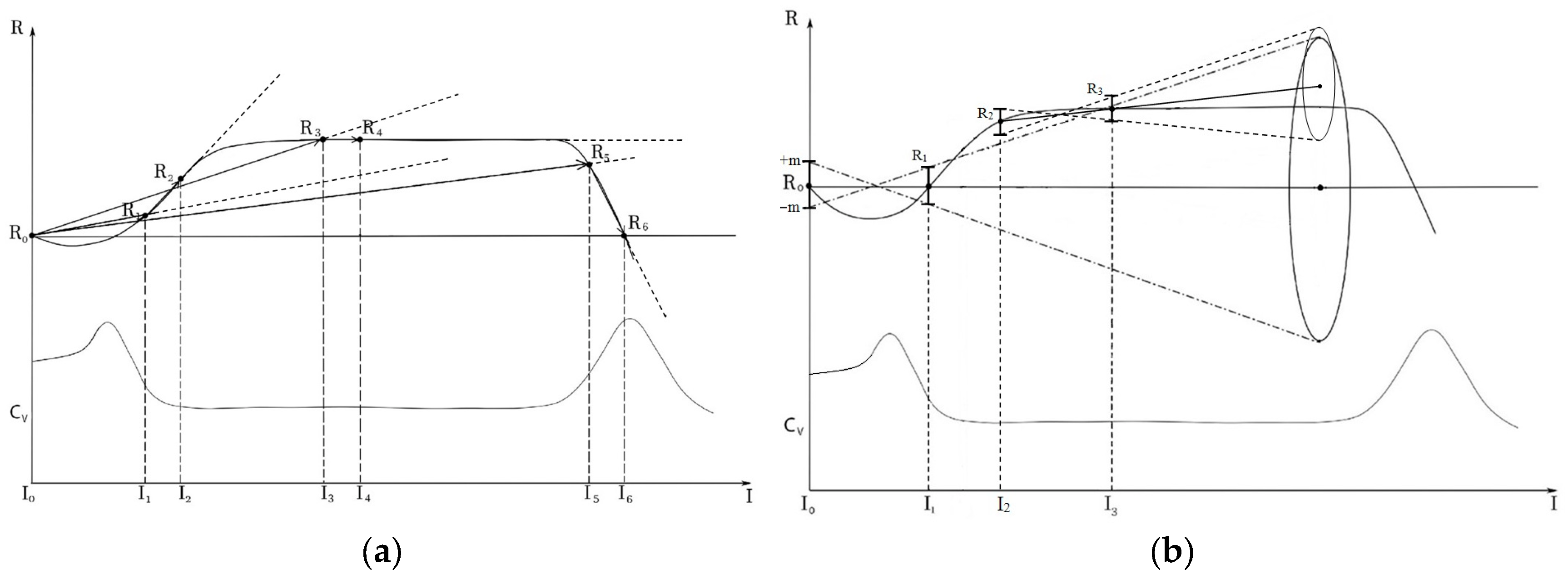
The use of the phototropic behavioral response of mesoplankton as the indicator of altering exposure implies photostimulation, registration of changes in response in comparison with the background response and determination of quantitative characteristics of the response model. The intensity of response to a stimulus may serve as an indicator of the test system state (in this study—planktonic ensemble) for the linear model: response function R = f(I) of the intensity of illuminating light I. The change of this indicator demonstrates the change in the state of the studied system.
This indicator is commonly measured by calculating the change in response in relation to the background value per unit of light intensity increase (slope of vector
to abscissa axis, where R
0 and R
N—values of the response function at zero and N stimulation). It should be considered that the test illumination is capable of changing the state of a system; therefore, R = f(I) can differ from the linear form passing through standard phases of activation, resistance and oppression [
44]. In other words, the forecast of change in a biological system according to its response to a random intensity of attracting light may markedly differ from reality.
Figure 3a shows that forecasts on the basis of vectors
correspond to a standard method. It is possible to obtain a wide range of vectors depending on the intensity of the stimulating light (I
1, I
2, I
3, …) and the transition of a system to a new state. The predictive value of such vectors is quite doubtful. For example, all three specified vectors show that a further increase in attracting light intensity in R
1, R
3 and R
5 states will lead to response growth. However, it is true only when the system is in the activation phase (alarm response) R
1. In the phase of stability (resistance) R
3 and, furthermore, in the phase of oppression R
5, the response inversion is even possible.
Notably, the accuracy of predicting the dynamics of the state of the system decreases for several reasons (
Figure 3b): (1) non-zero variability in the behavioral response of plankton in the background (inter-individual variability); (2) increased variability of the behavioral response in case of the insufficient intensity of photostimulation, which does not activate the functional system ensuring the behavioral response. Due to high variability (Cv) in the background (spread of R values around R
0: R
0 ± m) and in the photostimulation (spread of R values around R
1) the single photostimulation I
1 leads to a wide extrapolation ‘funnel’ of the response function.
An alternative solution to this situation is paired photostimulation. The first, lower intensity illumination (I
2,
Figure 3b) activates the functional system [
45] responsible for the behavioral response, which reduces the inter-individual variability and entropy in the system [
46]. This decreases the coefficient of variation. The second, more intense illumination (I
3,
Figure 3b), also being within the activation range of the functional system, causes crustaceans to move at a rate appropriate to their physiological state.
Figure 3b indicates that such paired photostimulation (I
2–I
3) significantly narrows the extrapolation funnel of vector
and allows improving the accuracy of measuring the change in the state of the planktonic community during ecotoxicological exposure.
Figure 3a shows that paired photostimulations I
1–I
2, I
3–I
4, I
5–I
6 correspond to this approach. Slope angles of the corresponding vectors
characterize the responsiveness of the system.
Hereafter, I1 refers to the intensity of the first light stimulus, and I2 to the second.
2.5. Experimental Design
During laboratory experiments, the DHC was placed in a 90 dm
3 tank filled with 50 dm
3 of artificial seawater [
42], which ensured that the DHC optical part was in the aqueous medium. One hour before the start of the experiment 200 ± 10 crustaceans of the synchronized daily culture were carefully placed into the tank with the DHC, which created a concentration of 4 ind/dm
3 (4000 ind/m
3) corresponding to the concentration in natural reservoirs. The increase in the concentration of crustaceans up to 30 ind/dm
3 does not cause erroneous results.
Eight cycles with further hologram recording were performed after the adaptation of crustaceans: one (first) cycle before exposure (background value—Bv) and seven consecutive cycles every 10, 30, 60, 90, 120, 150 and 180 min after water or pollutant were added. Each cycle included the hologram recording in darkness (without attracting light, 5 min, 10 holograms), after the activation of the attracting light with the intensity of 1150 lx (I1, 5 min, 10 holograms) and subsequent second photostimulation with the lighting intensity of 3450 lx (I2, 5 min, 10 holograms). This was followed by a 15 min pause to restore the number of crustaceans. Pollutants and clean water were added in the amount of 0.5 dm3.
The results of processing 10 sequentially registered holograms were used to calculate the average concentration of crustaceans during the first and second photostimulation stages (C1 and C2, respectively), as well as the ratio of the growth in the concentration of crustaceans during the transition from the first to the second intensity in the concentration of crustaceans during the second intensity: ΔC/C2= (C2 − C1)/C2 × 100%.
Statistical processing was performed using Statistica 10 (StatSoft, Inc., Tulsa, OK, USA). Parametric or non-parametric methods were chosen after checking the registered indicators for the normality of distribution according to the Shapiro-Wilk’s W-test. The average data is given in Mean ± Std.err. of mean. The difference from the zero slope of the fitting lines was determined according to the Student’s t-test (two-sided p level) due to the normal distribution of the obtained values.
3. Results
The validity of the paired photostimulation for prompt monitoring of the state of a marine environment according to the phototropic behavioral response of zooplankton was studied in laboratory conditions on the saltwater test organisms A. salina and M. salina using such pollutants as K2Cr2O7, nanoplastics and microplastics.
Earlier we showed that a multivariate regression model of the response to the action of a toxicant (completeness of the model adjusted R
2 = 0.74) may only require predictors reflecting the concentration of crustaceans in the measured volume at different lighting levels. The parameters of speed and direction of their movement were not so informative [
37]. Therefore, the subsequent analysis uses the following parameters: concentration of crustaceans without photostimulation (C
0), at light intensity I
1 (C
1) and I
2 (C
2), and derived values ΔC, ΔC/C
2.
3.1. Response of A. salina to Clean Seawater
The stability over time of the phototropic response parameters for paired photostimulation and effect of the introduced clean seawater in a volume of 0.5 dm3 equivalent to the volume of added pollutants on the phototropic response were studied in the control series.
Sixty minutes after the crustaceans were placed in the experimental tank, their distribution throughout the tank was registered, and the speed and nature of swimming were restored, which indicated the adaptation of A. salina to the experimental conditions.
The concentration of crustaceans in the illuminated water column is a typical response to the attracting light. The average concentration over 5-min registration periods increases compared with the background without attracting light (Bv) C = 2.1 ± 0.57 ind/dm3 to 5.8 ± 1.04 ind/dm3 with the illumination of 1150 lx. In response to the second photostimulation with the lighting intensity of 3450 lx, an additional increase of almost 1.5 times was registered in the number of A. salina individuals (up to 8.17 ± 0.58 ind/dm3).
The introduction of 0.5 dm3 of clean seawater did not reveal significant changes in the response of nauplii to the paired photostimulation over the entire observed period (180 min). The ΔC/C2 ratio varied in the range of 56–64% over 180 min of the experiment with an average of 58.6 ± 3.5%.
Thus, neither the introduction of the additional volume of liquid, nor the three-hour exposure in intact medium changed the parameters of the phototropic response of crustaceans to the paired photostimulation.
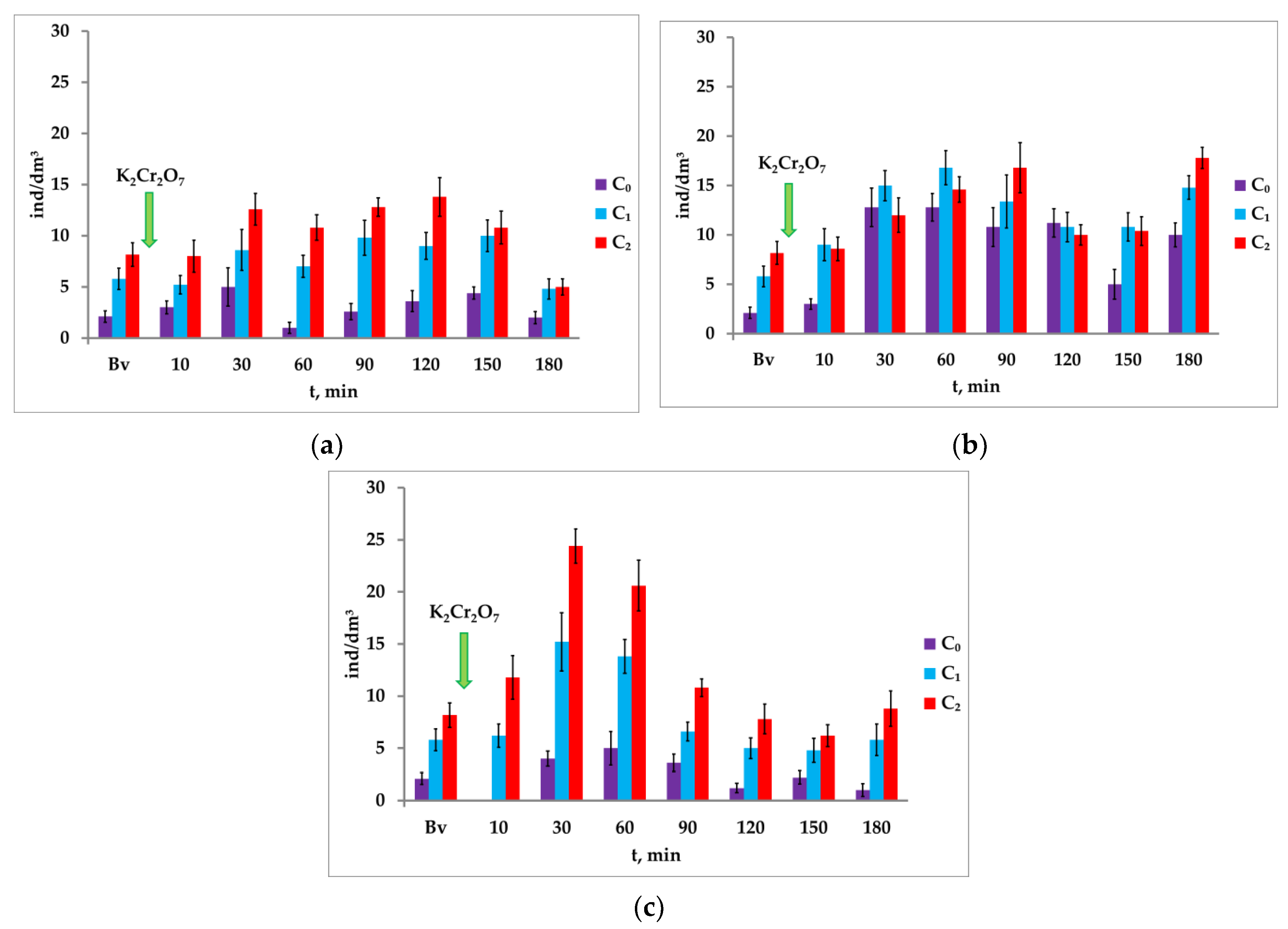
3.2. Response of A. salina and M. sakina to Paired Photostimulation in the Presence of K2Cr2O7
Significant differences were noted in the medium contaminated with potassium bichromate (C = 4.0 mg/dm
3) when the crustaceans gathered in the illuminated water column during stimulation with light of different intensity (
Figure 4a). Within 90 min after the toxicant was introduced, the concentration of
A. salina increased to a greater extent in response to attracting light with intensity I
2 = 3450 lx (second stage of paired photostimulation). During the photostimulation with intensity I
1 = 1150 lx, the concentration of crustaceans ranged from C = 2.1 ± 0.57 ind/dm
3 to 10.0 ± 1.55 ind/dm
3 with a maximum of 150 min. During subsequent attracting light with intensity I
2, the concentration of crustaceans was significantly higher and varied from 8.17 ± 1.16 ind/dm
3 to 13.8 ± 1.90 ind/dm
3 with a maximum of 120 min.
A slightly different phototropic response of crustaceans to the paired photostimulation was noted during the contamination of the medium with potassium bichromate at the concentration of 8.0 mg/dm
3 (
Figure 4b): during 60 min the positive phototaxis prevailed for light with lower intensity I
1; after 90 min and further for light with higher intensity I
2.
The number of nauplii exceeded the background values in four cycles following the background in the medium contaminated with potassium bichromate at the concentration of 16.0 mg/dm
3 (
Figure 4c). In each cycle, in response to the second photostimulus with intensity I
2, the positive phototaxis showed an increase compared with the light with intensity I
1. The maximum concentration of nauplii (C = 24.4 ± 1.65 ind/dm
3) to the second photostimulus was registered 30 min after the toxicant was introduced. After an hour of exposure to potassium bichromate, we registered the inhibition of the phototropic response, and
A. salina nauplii responded less to paired photostimulation; at the same time, their number in the controlled volume was reduced with each subsequent cycle.
ΔC ratio—an increase in the concentration of nauplii in response to the second light stimulus: ΔC = C2 − C1—may indicate the change in the phototaxis of nauplii to paired photostimulation. Within 120 min of exposure to potassium bichromate (4.0 mg/dm3), the ΔC ratio increased to 4.8 ± 2.3 ind/dm3 and exceeded its value in the medium without a toxicant (ΔC = 2.37 ± 1.56 ind/dm3). However, by the end of the observed period (minutes 150 and 180), the ΔC dropped sharply to 0.8 ind/dm3 and 0.2 ind/dm3, respectively. In a medium contaminated with potassium bichromate at the concentration of 8.0 mg/dm3, the changes in the ΔC are unreliable almost throughout the entire observed period, except for the first (background) and last (180 min after the introduction of the toxicant) cycles. In the medium contaminated with potassium bichromate (16.0 mg/dm3), the ΔC decreased starting from minute 60 to minute 150 in each subsequent cycle, and its value depended on the exposure time of the toxicant.
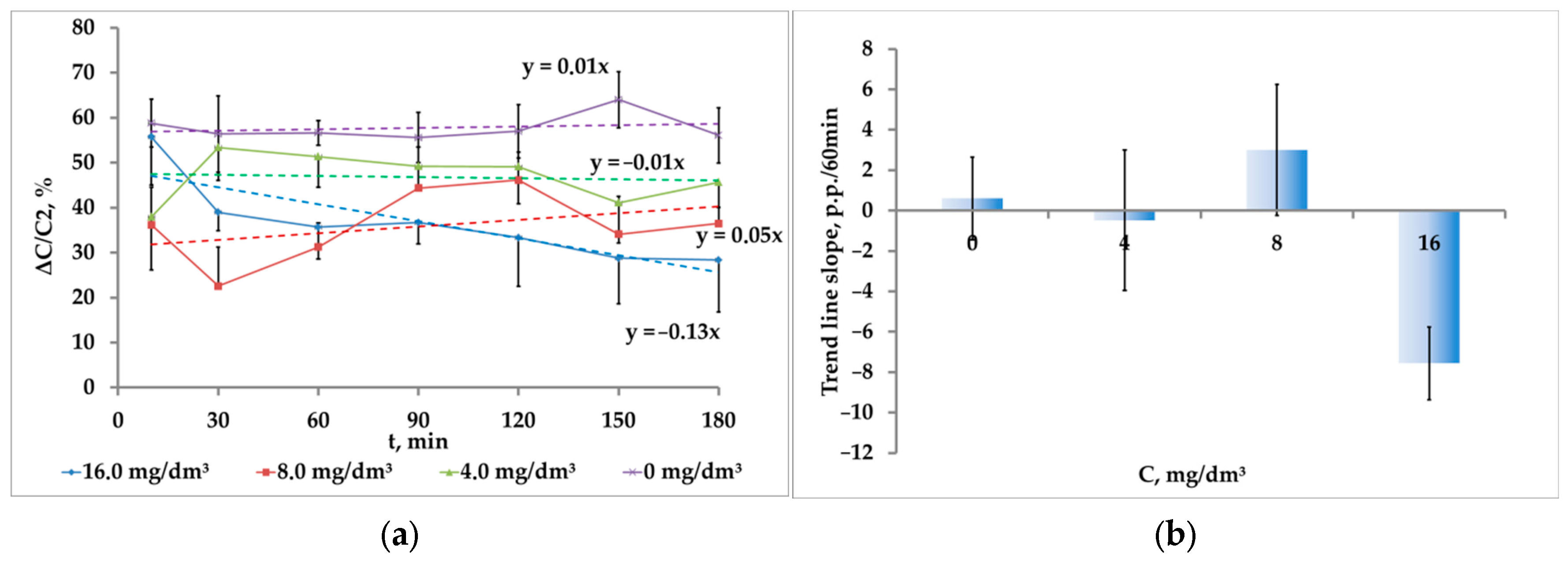
It is possible to ensure a more visual analysis of the results by presenting them as the percentage of the increase in the concentration of crustaceans during the transition from the first to the second intensity of photostimulation to the value of phototaxis at the maximum illumination: ΔC/C
2 × 100% (
Figure 5a).
Regardless of the fluctuations in this indicator, it is possible to conduct an integrated assessment over the observation period using an approximating linear dependence. In this case, the coefficient at the independent variable, being the tangent of the inclination angle of the approximating line, reflects the rate of change of the indicator, and its sign—the direction of changes, increase or decrease (
Figure 5b).
The exposure to the medium contaminated with potassium bichromate at the concentration of 16 mg/dm3 caused the time-increasing inhibition of the phototaxis. The ΔC/C2 decreased at a rate of 7.6 ± 1.8 p.p./h and was significantly different from zero (p = 0.002, t = 4.2, df = 9). This pattern was not observed at lower concentrations of K2Cr2O7 and in its absence. In the medium without a toxicant, as well as in the contaminated medium at K2Cr2O7, concentrations of 4 mg/dm3 and 8 mg/dm3, the trend line slope almost did not differ from zero: 0.6 ± 2.0 p.p./h (p = 0.78, t = 0.29, df = 9), −0.5 ± 3.5 p.p./h (p = 0.89, t = 0.14, df = 9) and 3.0 ± 3.2 p.p./h (p = 0.38, t = 0.92, df = 9), respectively.
For K
2Cr
2O
7, the cumulative dose of exposure that changes the behavioral response (16 mg∙dm
−3∙h = 48 mg∙dm
−3∙h) is significantly less than LD
10 (16.6 × 48 h = 800 mg∙dm
−3∙h) determined by the death rate of crustaceans. Notably, this cumulative dose of exposure for
A. salina is 200 times higher than the cumulative dose of exposure that caused a significant change in the phototropic response of freshwater
Daphnia magna S. amounting to 0.24 mg∙dm
−3∙h [
30].
The response of M. salina to potassium bichromate is similar to that of A. salina, but is shifted towards lower concentrations. The primary increase in the rate of change of the ΔC/C2 ratio from −6.9 ± 2.0 p.p./h in clean water to 24.5 ± 3.8 p.p./h (p < 0.0001, t = 7.4, df = 18) was observed at the concentration of 4 mg/dm3, but not at 8mg/dm3, as with A. salina. Further dynamics demonstrated the reduction of this indicator to −18.7 ± 1.6 p.p./h (p = 0.0001, t = 4.8, df = 18) at 16.0 mg/dm3 and to −21.3 ± 4.0 p.p./h (p < 0.004, t = 3.3, df = 18) at 16 mg/dm3.
In other words, in terms of the phototropic response M. salina show a slightly higher sensitivity to potassium bichromate compared with A. salina.
3.3. Phototropic Response of Saltwater A. salina and M. salina during the Contamination of the Medium with Nanoplastics
3.3.1. Response of A. salina to Paired Photostimulation in the Presence of Nanoplastics
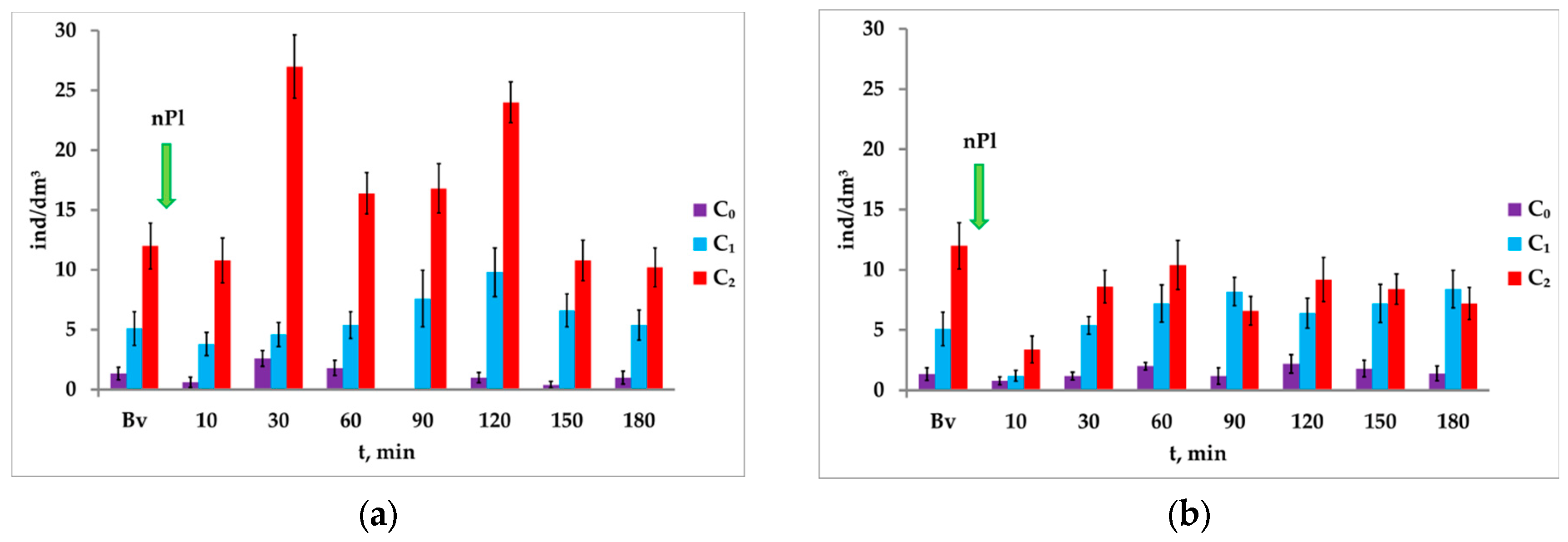
In response to attracting light I1, the concentration of nauplii in the controlled volume before the introduction of nanoplastics increased more than three times (from 1.35 ± 0.52 to 5.10 ± 1.40 ind/dm3). The second light stimulus I2 led to an increase in the number of A. salina by almost 9 times compared with the background value (up to 12.00 ± 1.93 ind/dm3).
The medium contaminated with nanoplastics (0.1 mg/dm
3) showed an additional increase in the concentration of
A. salina throughout the experiment in response to the attracting light with higher intensity (
Figure 6a).
The ΔC in the medium contaminated with nanoplastics from minute 30 to minute 120 was several times higher than the values in the clean medium. The maximum movement of a significant number of nauplii to the illuminated water column in response to the second light stimulus of the paired photostimulation is registered at minute 30 (ΔC = 22.4 ± 2.83 ind/dm3). With further exposure of crustaceans (from minute 60 to minute 120) the difference in the concentration growth to two successive light stimuli decreased, but remained above the values registered in the medium without nanoplastics. The last two cycles registered the minimum ΔC values (4.2 ± 2.18 ind/dm3 in 150 min and 4.8 ± 2.05 ind/dm3 in 180 min after the application of nanoplastics).
In a contaminated aqueous medium with a nanoplastic concentration of 1.0 mg/dm
3, the number of nauplii in response to attracting light I
1 after a significant decrease (from 5.1 ± 1.40 to 1.20 ± 0.44 ind/dm
3) at minute 10 of the exposure was restored after 30 min of exposure (
Figure 6b).
Attracting light I2 caused additional movement of crustaceans into the illuminated water column (except for minute 90). However, the increase in the concentration of nauplii in response to the second light stimulus was significantly less than in the medium with the concentration of nanoplastics at 0.1 mg/dm3.
Therefore, during the contamination of the medium with nPl at the concentration of 0.1 mg/dm3 with similar dynamics, a more pronounced phototropic response of A. salina to the second light stimulus was registered than at the concentration of 1.0 mg/dm3.
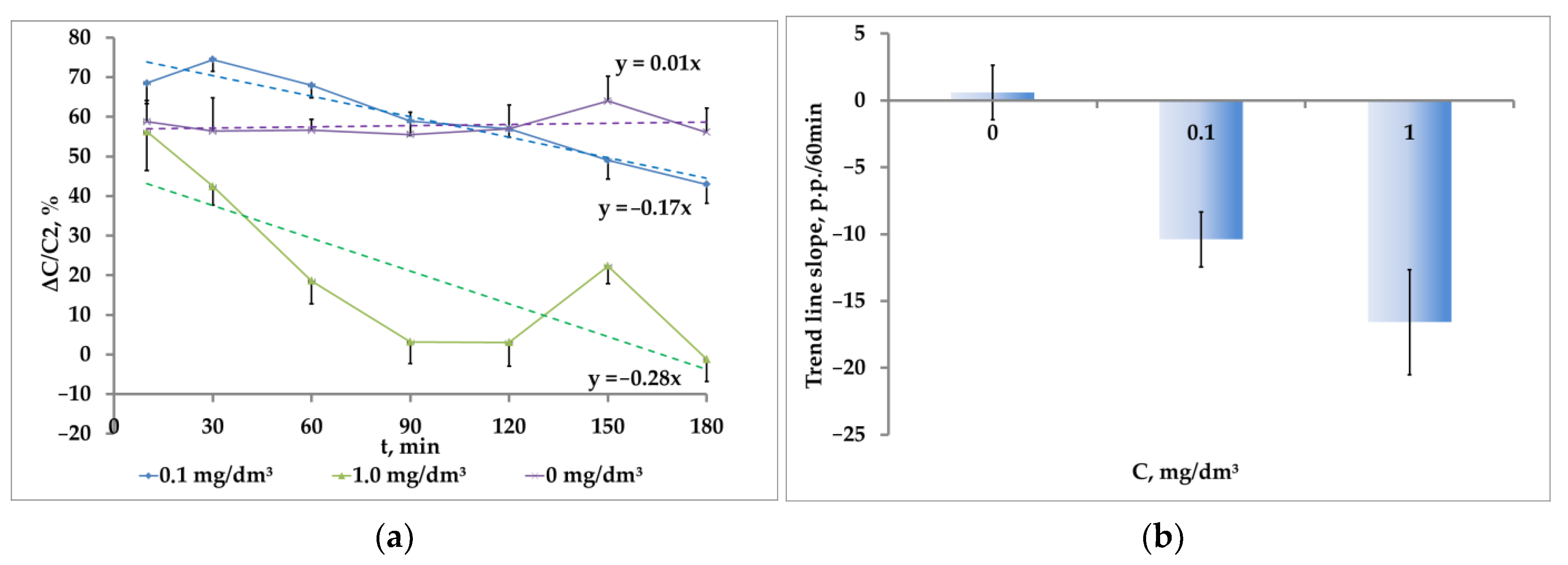
The analysis of the dynamics of ΔC/C
2 parameters showed the increasing inhibition of phototaxis to paired photostimulation with regard to the exposure of crustaceans to the tested concentration of nanoplastics (
Figure 7a).
The ΔC/C
2 ratio decreased at the concentration of nanoplastics at 0.1 mg/dm
3 as the exposure time of crustaceans in the contaminated medium increased. At the same time, the rate of its decrease was 10.4 ± 2.1 p.p./h being significantly different from the values in clean water (
p = 0.001, t = 3.8, df = 18). At the nanoplastics concentration of 1.0 mg/dm
3, the rate of phototaxis inhibition increased to 16.6 ± 3.9 p.p./h and was significantly different from the values in clean water (
p = 0.001, t = 3.9, df = 18). The differences (by 6.2 p.p./h) at the studied concentrations were statistically unreliable (
p = 0.09, t = 1.8, df = 18) (
Figure 7b).
The cumulative dose of exposure causing a significant change in the phototaxis was 0.3 mg∙dm−3∙h, which is significantly less than the LD10 = 1.9 × 48 h = 91 mg∙dm−3∙h determined by the death rate of artemia crustaceans.
3.3.2. Response of M. salina to Paired Photostimulation in the Presence of Nanoplastics
The dynamics of the phototropic response of
M. salina in the nPl-contaminated aqueous medium was slightly different from the dynamics of the
A. salina response (
Figure 8).
An additional increase in the concentration of M. salina crustaceans was registered in response to attracting light only in 4 cycles after the introduction of nPl (0.1 mg/dm3). When the medium was contaminated with nanoplastics at a higher concentration (1.0 mg/dm3), the paired photostimulation caused irregular changes in the number of M. salina crustaceans. Thus, a phase change in the inhibition rate of the phototropic response was observed.
At the nPl concentration of 0.1 mg/dm
3 with an increase in exposure time, the ΔC/C
2 decreased at a rate of 28.2 ± 2.4 p.p./h (
Figure 9a) and was significantly different from the values in clean water (
p < 0.0001, t = 9.1, df = 18). At the nPl concentration of 1.0 mg/dm
3, the rate of phototaxis inhibition decreased to 8.2 ± 3.0 p.p./h (
p < 0.01, t = 9.1, df = 18), and was different from values in clean water and even more significantly different (by 20 p.p./h,
p < 0.002, t = 3.7, df = 18) from the values at the concentration of 0.1 mg/dm
3 (
Figure 9b).
It should be noted that the ΔC/C2 dynamics has a phase nature, and a statistically significant change in the inhibition rate of the phototropic response is observed at the 3 h exposure interval. This duration of the observation interval should be taken into account to generate the contamination alarm signal.
The cumulative dose of exposure causing a significant change in the phototropic response was 0.3 mg∙dm−3∙h, which coincides with the cumulative dose for A. salina. However, the rate at which the phototropic response was inhibited at an equal dose for M. salina was 17.8 p.p./h (p < 0.0001, t = 3.7, df = 18) higher than for A. salina.
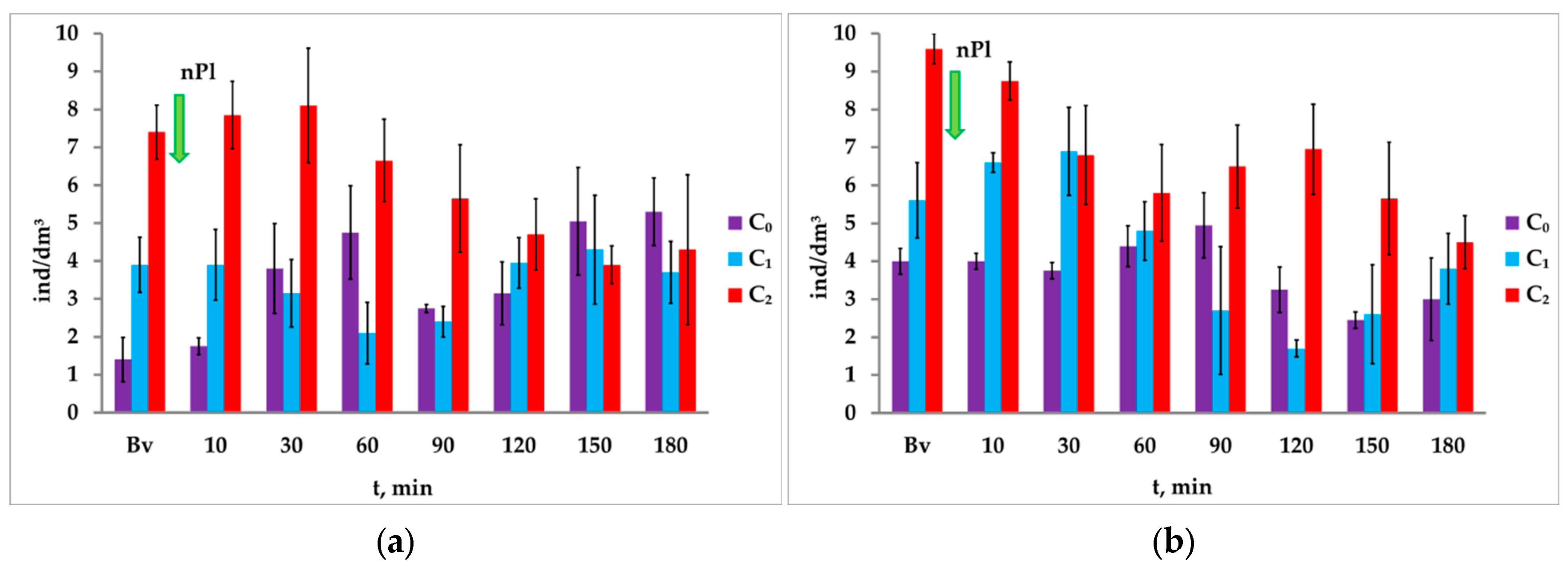
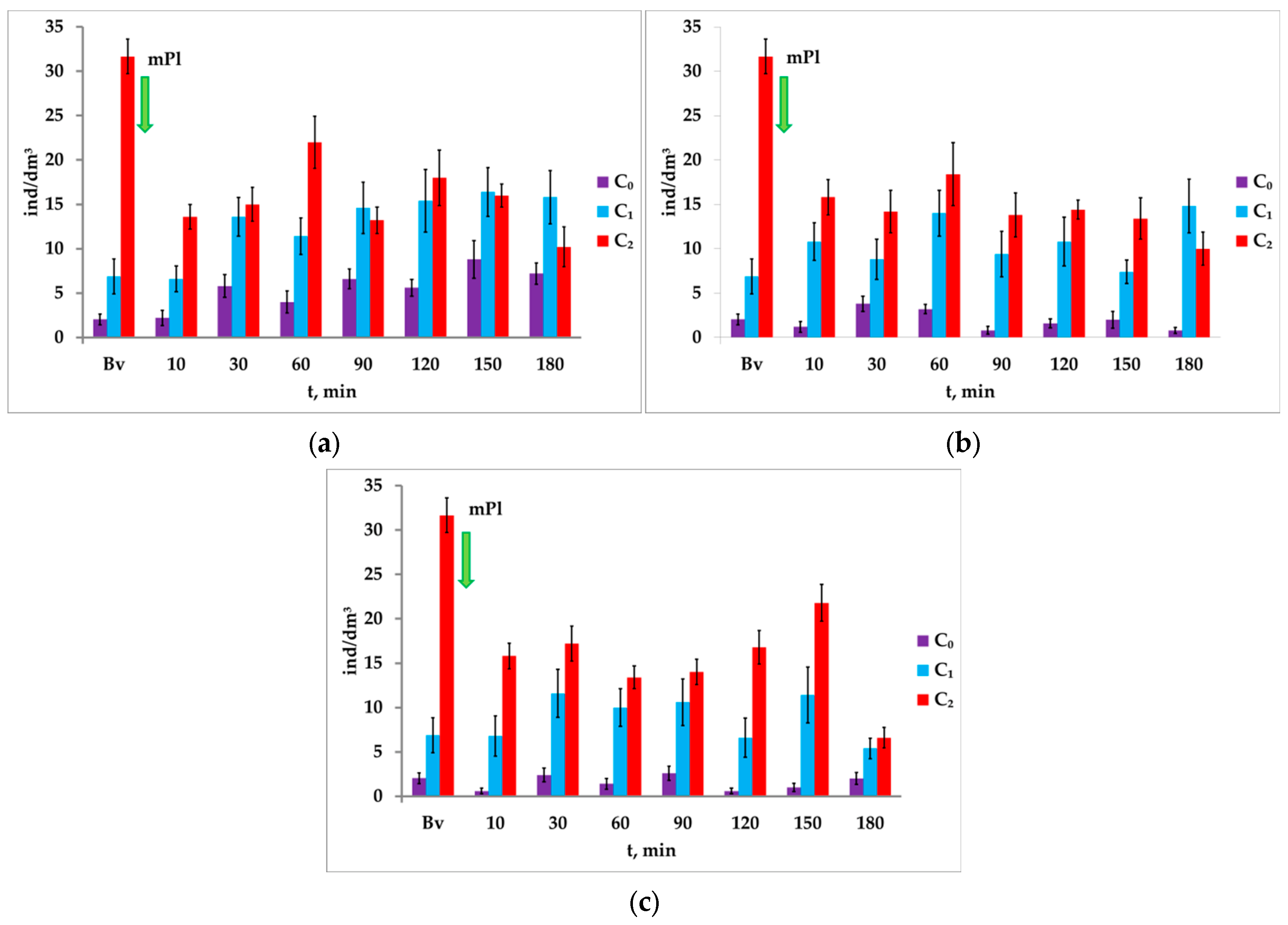
3.4. Response of A. salina to Paired Photostimulation in the Presence of Microplastics
It was noted that the microplastics at the concentration of 0.05 mg/dm
3 in the first 10 min did not affect the response of
A. salina nauplii to the first light stimulus (I
1), but significantly reduced the phototropic response (from 31.67 ± 1.97 ind/dm
3 to 13.60 ± 1.44 ind/dm
3) to the second light stimulus. With the increase in exposure time to microplastics, the concentration of nauplii to the first light stimulus I
1 of the paired photostimulation varied from 6.6 ± 1.46 ind/dm
3 to 16.4 ± 2.75 ind/dm
3 (
Figure 10a).
Despite the pronounced movement of nauplii into the illuminated column of the microplastic-contaminated medium under attracting light I2, their concentration during the entire experiment was significantly lower than in the background. Furthermore, in response to the light stimulus I2, a negative phototaxis was registered in the last cycle (after 180 min)—a decrease in the number from 15.8 ± 1.3 to 10.2 ± 2.2 ind/dm3 (p = 0.042, t = 2.19, df = 18).
It was noted that after the introduction of microplastics, the ΔC ratio during the experiment changed unevenly: after 10 min it changed sharply, and then it gradually (throughout the experiment) decreased and was fully negative by the end of the experiment.
A similar response of
A. salina crustaceans to paired photostimulation was registered at the microplastic concentrations of 0.1 mg/dm
3 and 1.0 mg/dm
3 (
Figure 10b,c).
A significant increase in the concentration of crustaceans in response to the second attracting light I2 during paired photostimulation was noted in each registered cycle, including the background, with the exception of the last cycle (in 180 min of exposure to microplastics with the concentration of 1.0 mg/dm3). The change ranged from 24.8 ind/dm3 (background) to 1.20 ind/dm3 (minute 180).
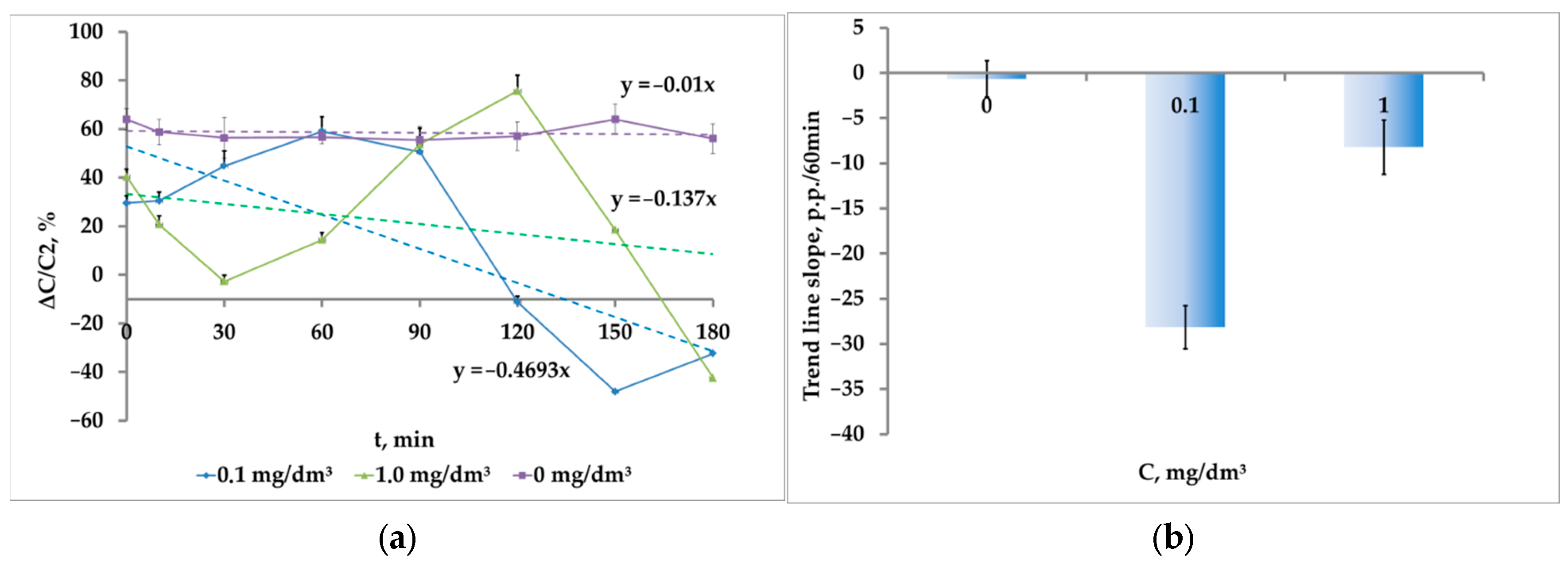
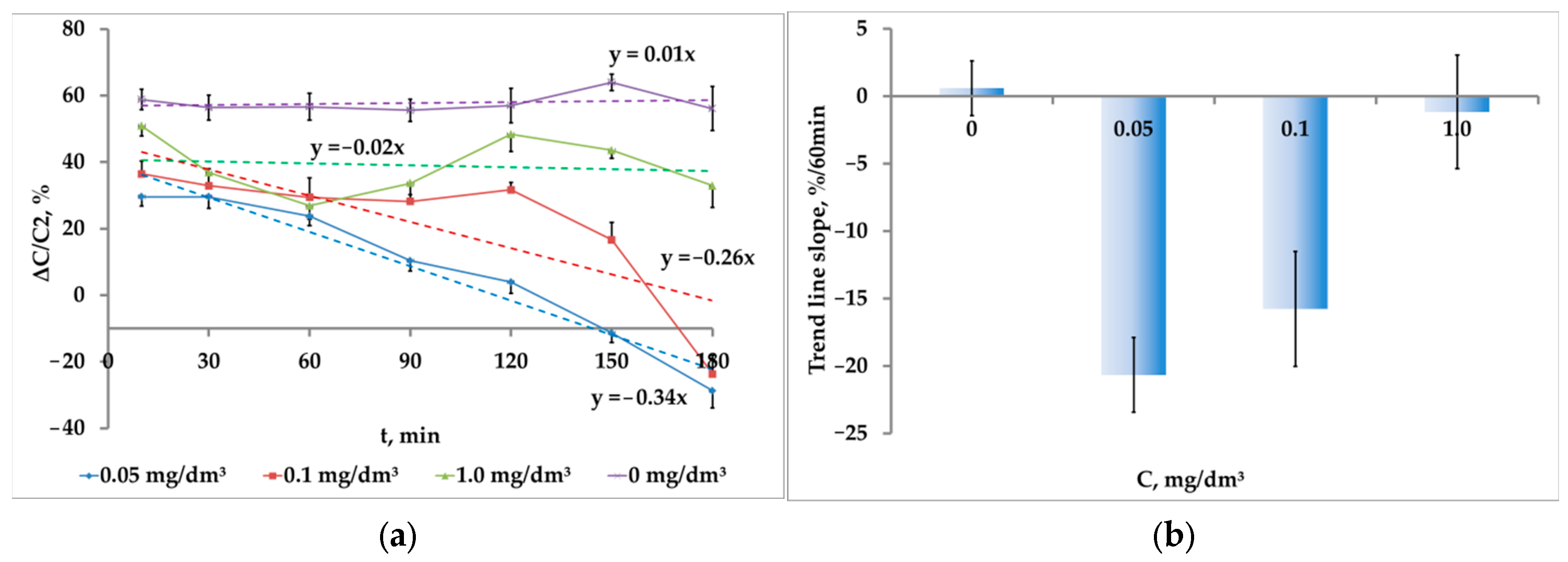
While the crustaceans are exposed to the medium contaminated with microplastics, there is also a gradual inhibition of phototaxis up to the response sign inversion (
Figure 11a). However, unlike nanoplastics, the dependence on concentration is reverse. The highest rate of phototaxis inhibition (20.6 ± 2.8 p.p./h) was registered at the concentration of 0.05 mg/dm
3. As the concentration increased, the inhibition rate decreased to 15.8 ± 4.3 p.p./h at the microplastic concentration of 0.1 mg/dm
3; and at the microplastic concentration of 1.0 mg/dm
3 , to almost the background value (1.2 ± 4.2 p.p./h) (
Figure 11b). It should be noted that at the concentration of 1.0 mg/dm
3 , the difference from zero was unreliable (
p = 0.78, t = 0.28, df = 9).
4. Discussion
This paper describes the results obtained using the DHC camera, which allows conducting similar in situ studies in freshwater and saltwater reservoirs at depths up to 500 m.
The available literature does not contain data on the use of paired photostimulation in the bioindication of pollution of aquatic ecosystems. However, there are numerous data on the light-dependent migration of zooplankton [
1,
34,
47,
48,
49,
50]. Moreover, this migration changes up to the inversion sign of the phototaxis during the contamination of the aquatic medium [
1,
32,
33,
48,
49,
51,
52], including with the photoreactive particles, such as zinc nanoxide and microplastics [
53,
54]. There is evidence of the correlation of the phototropic response with wavelength, intensity and duration of applicable light stimuli [
32,
34,
55,
56]. Furthermore, the results revealed differences in behavior both at the interspecific and intraspecific levels [
34], as well as among males and females [
35].
This study established that despite the phase directionality of changes, the general pattern during the contamination of the medium with nano- and microplastic particles is the inhibition of the phototropic response of the saltwater crustaceans A. salina and M. salina to paired photostimulation.
Potassium bichromate K
2Cr
2O
7 is a model toxicant in most standard biotests [
20,
21,
26,
27] that is used to determine the sensitivity of hydrobionts; therefore, it is used by numerous authors as a test exposure in laboratory studies. The response to the toxic exposure of K
2Cr
2O
7 in increasing concentrations is explicit and gradual, which makes it possible to compare the sensitivity of test organisms and test responses during biotesting.
It was established that the cumulative dose of exposure to potassium bichromate (product of pollutant concentration at the time of exposure in the contaminated medium), which causes a significant change in the phototropic response of
A. salina to paired photostimulation, was 48 mg/dm
3∙h (16 mg/dm
3 × 3 h), which is significantly less than NOEL (no observed effect level) = 16.6 × 48 h = 800 mg∙dm
−3∙h defined by the death rate of crustaceans. Earlier works showed that the cumulative dose of exposure for
A. salina is 200 times higher than the cumulative dose of exposure, which caused a significant change in the phototropic response of freshwater
D. magna S., which is 0.24 mg∙dm
−3∙h [
30].
A significant change in the phototropic response of
M. salina registered at the cumulative dose of 24 mg/dm
3∙h indicates a higher sensitivity of these crustaceans compared with
A. salina. It is quite difficult to compare the data obtained by us with those available in the literature due to different experimental designs, and mainly due to limited data on the phototropic response of saltwater crustaceans. At the same time, there is data [
57] showing a higher sensitivity of freshwater
Moina micrura Kurz to the action of K
2Cr
2O
7 compared with
D. magna S. and
D. pulex. A few studies determined the cumulative dose of exposure on the behavioral responses of marine mezoplankton; however, there are studies [
58] indicating the dependence of the immobilization rate of
A. franscana on the concentration and time of exposure to nanosized pollutants.
The differences in the phototropic response of the
A. salina and
M. salina communities were particularly evident when the medium was contaminated with nanoplastics (
Figure 7b and
Figure 9b). Both species of crustaceans show significant inhibition of the phototropic response to paired photostimulation at the nanoplastic concentration of 0.1 mg/dm
3. However, the inhibition rate of the phototropic response at this dose was 2.8 times higher for
M. salina compared with
A. salina.
A more pronounced inhibition of the phototropic response of M. salina to paired photostimulation during the contamination of seawater with nanoplastics compared with that of A. salina gives them higher priority during the bioindication of the ecological well-being of saltwater ecosystems.
The comparison of the toxicity of nano- and microplastics for euryhaline crustaceans assumed that the toxic properties of plastics will increase as their size decreases and, accordingly, based on the literature [
3,
59,
60], increase their bioavailability for crustaceans. However, with an equal inhibition rate we revealed a significant decrease in the phototropic response of
A. salina (
Figure 7b and
Figure 11b) at a lower microplastic concentration (0.05 mg/dm
3) compared with nanoplastic (0.1 mg/dm
3).
The conclusion that micro- and nanoplastics are equally toxic is contradictory since for M. salina, we have not revealed any significant inhibition of the phototropic response in the medium contaminated with microplastics in the range of concentrations we used.
This fact may be explained by the difference in the size of crustaceans: M. salina crustaceans are 1.5–2 times smaller than those of A. salina. The smaller size, including the digestive system, reduces the bioavailability of plastic particles, especially noticeable for relatively large micron-sized particles. It is natural that with an even larger particle size, the microplastics are no longer dangerous for this mesoplankton community, at least as an anti-nutrient or ballast agent that adsorbs and concentrates harmful substances during transmission along the food chain.
Thus, the toxicity of plastics to mesoplankton is determined not only by the size of its particles, but also by the crustacean species themselves. The correlation between the sensitivity and species, size and even food preferences of crustaceans is described in a study [
61]. This fact causes the need to control the collective response of the mesoplankton community in a controlled reservoir, which is only possible by measuring the responses of autochthonous plankton in situ.
Along with the above patterns, some cases reveal the inversion of the dependence of the phototaxis inhibition on the increase in the pollutant concentration.
The noted increase in the inhibition rate of the phototropic response depends on the concentration of a toxicant. When the medium is contaminated with a molecular toxicant K
2Cr
2O
7 and a nanosized pollutant,
A. salina shows an increase (in modulus) of this rate. However, the revealed pattern is violated in the case of microplastic contamination of the
M. salina community (
Figure 11b), as well as in the contamination with nanoplastics (
Figure 9b).
There may be several reasons for this inverted response to the reduced toxicity of microplastics as their concentration increases. The reduction of the inhibitory effect of the higher nanoplastic concentration reflects the previously established phenomenon of the phase reduction of toxicity of dispersed nanomaterial systems [
23,
62] as their concentration increases. The mechanism of this phenomenon is associated with aggregation processes. It is generally known that a 10-fold increase in the concentration decreases the distance between nanoparticles by more than 2 times, which enhances the likelihood of collisions and the formation of large aggregates comparable to the size of microplastics.
Such aggregates, on the one hand, reduce the bioavailability of plastic particles, and on the other hand, enhance flotation thus redistributing the particles into the upper water layers and, accordingly, eliminating them from the zone of exposure on crustaceans. We previously showed that the aggregation rate of nanoparticles increases in the distilled water—drinking water—0.9% NaCl—3.0% NaCl series [
63]. This means that the elimination of free nanoparticles is observed the most in a saltwater medium. There is data confirming that the diversity of the size and buoyancy of microplastics may affect their vertical distribution in the water column in the oceans [
64].
It can be assumed that the leading process in changing the biological activity for nanoplastics is the bioavailability reduction due to aggregation, which is most pronounced for smaller M. salina crustaceans. The flotation process may come to the fore for microplastics with 2–15 μm particles, there may be the modulating influence to partial shielding and light scattering with relatively large (compared with bichromate molecular solution and nanosized plastic suspension) microplastic particles. However, these mechanisms can only operate in fairly narrow ranges of low microplastic concentrations.
Thus, the paired photostimulation of the behavioral response showed the validity of this method for the detection of the contamination of saltwater reservoirs with micro- and nanoplastics. Furthermore, the sensitivity of this method to the biological activity of low-toxic pollutants significantly exceeds the applied methods used to monitor the state of plankton communities in terms of pathogenicity.
5. Conclusions
The phototropic response of saltwater crustaceans to paired photostimulation (two successive stimuli for attracting light of increasing intensity) was expressed in a gradual increase in the concentration of crustaceans in the illuminated water column. The concentration growth at the first illumination intensity of 1150 lx is variable and does not allow making reliable conclusions on the changes in response to the introduction of pollutants. The increase in the concentration of crustaceans during the transition from the first to the second level of illumination (3450 lx) is more pronounced and stable. Moreover, the dispersion of this value is lower than the dispersion during the transition to this illumination from the state of darkness (single photostimulation).
When the medium is contaminated with a model toxicant K2Cr2O5, the inhibition of the phototropic response is registered, showing the decrease in the concentration growth of crustaceans in response to the illumination growth. The decrease increases as the toxicant concentration and exposure time in the contaminated medium increase.
Despite the phase nature of changes, the general pattern during the contamination of the medium with nano- and microplastic particles is the inhibition of the phototropic response of the saltwater A. salina and M. salina crustaceans to paired photostimulation.
The higher sensitivity of paired photostimulation of the behavioral response is confirmed compared with the standard methods of pollutant toxicity according to LD50. The toxicity of nano- and microplastics for euryhaline crustaceans is modulated by both the aggregation of nano- and microparticles and their different bioavailability for different species of crustaceans.
The paired photostimulation of the behavioral response showed the validity of this method for the detection of the contamination of saltwater reservoirs with micro- and nanoplastics after a short time of their exposure. This indicates a promising potential of this method for the alarm response in the monitoring of the ecological well-being of hydrobionts.




 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










