
Cross-Priming Approach Induced Beneficial Metabolic Adjustments and Repair Processes during Subsequent Drought in Olive

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
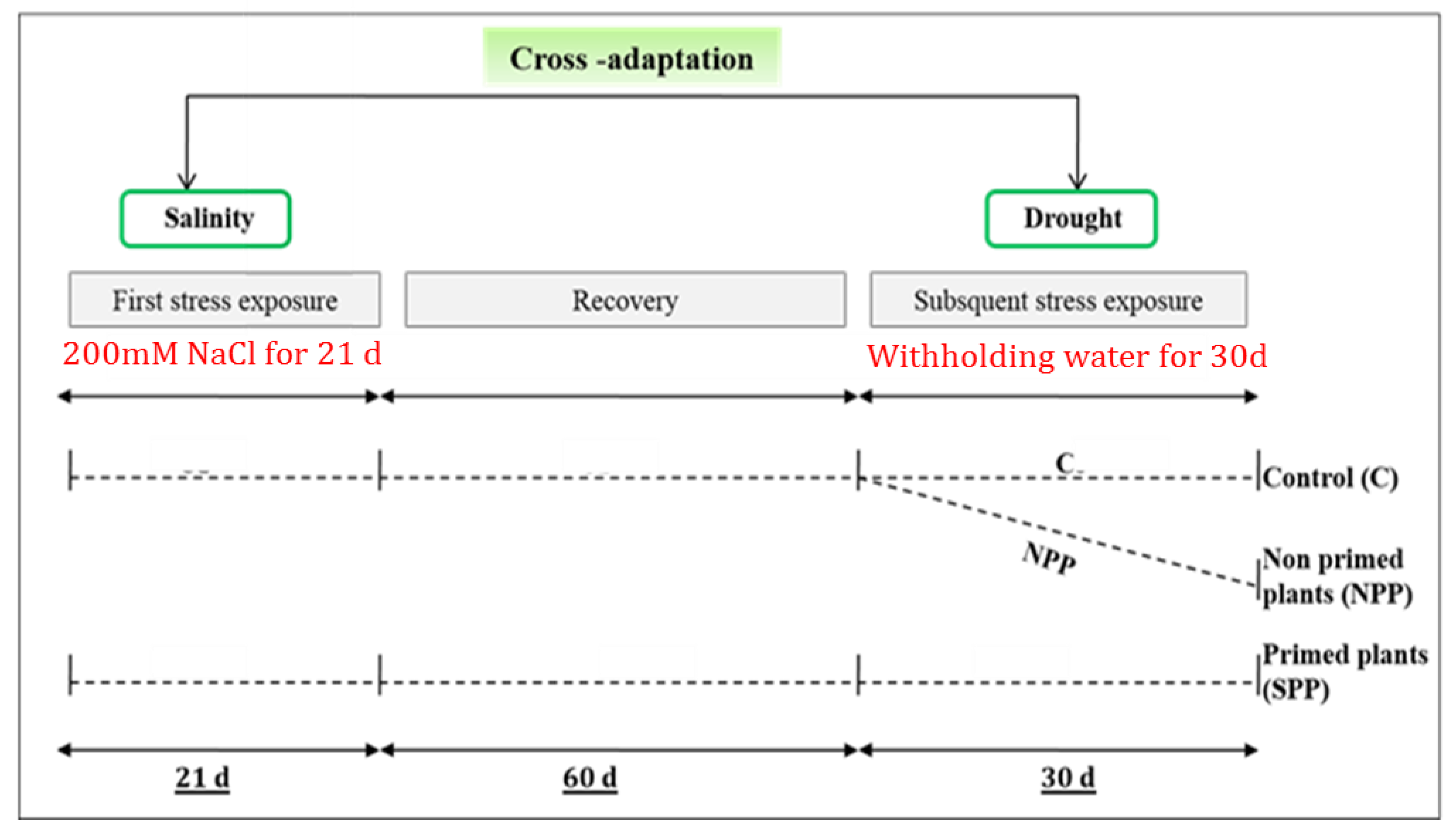
2.1. Greenhouse Conditions and Cross-Priming Application
2.2. Physiological Analysis
2.2.1. Growth and Relative Water Content Determination
2.2.2. Gas-Exchange, Pigment, and Fluorescence Measurement
2.2.3. Determination of Leaf Density
2.3. Biochemical Analysis
2.3.1. Proline and Sugar Content
2.3.2. Estimation of Lipid Peroxidation and Electrolyte Leakage
2.3.3. Protein Extraction and Antioxidant Enzyme Activities
2.3.4. Estimation of Total Phenol Amount and Its Antioxidant Activity
2.3.5. Lipid Extraction and Fatty Acid Determination
2.4. Statistical Analysis
3. Results
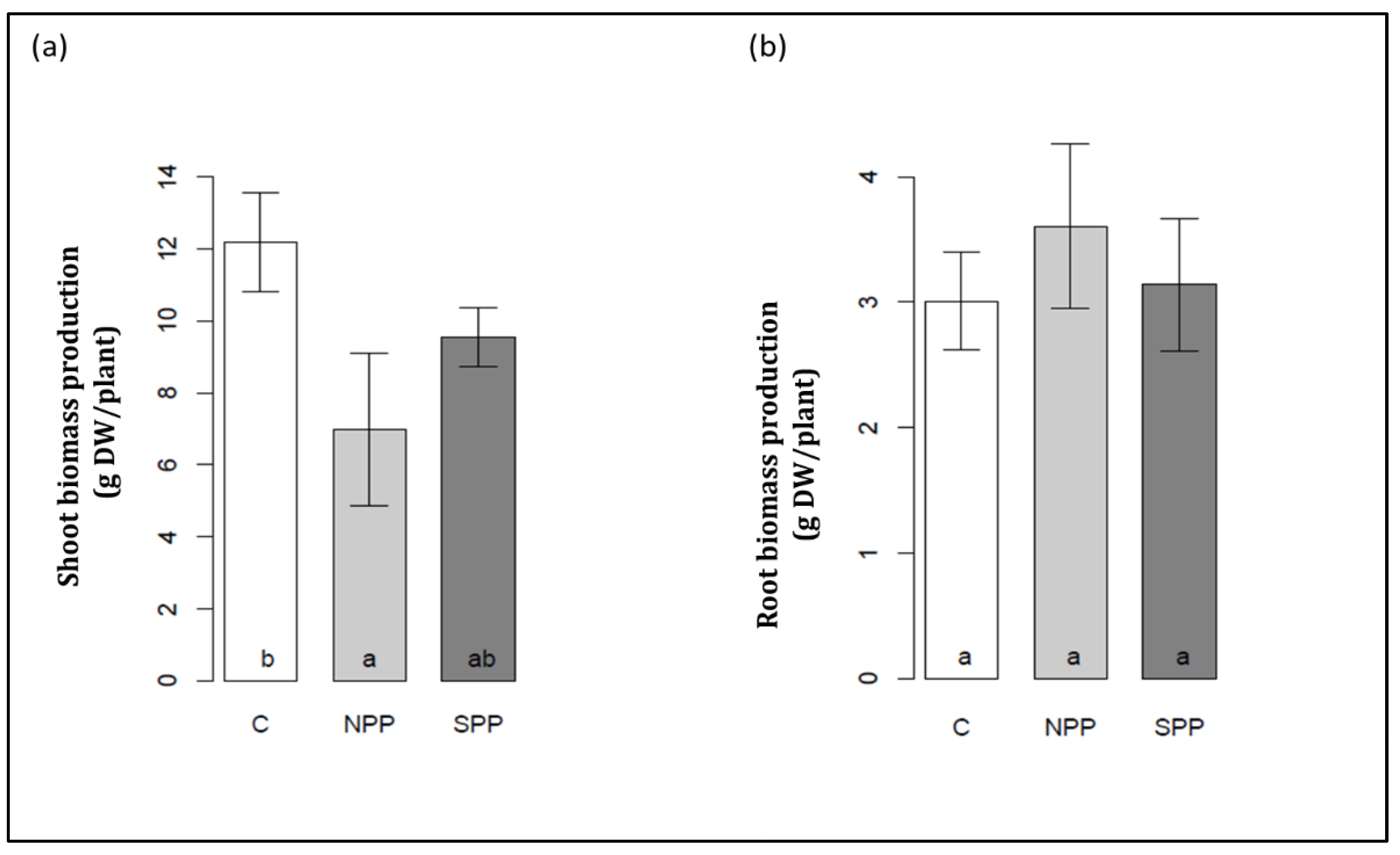
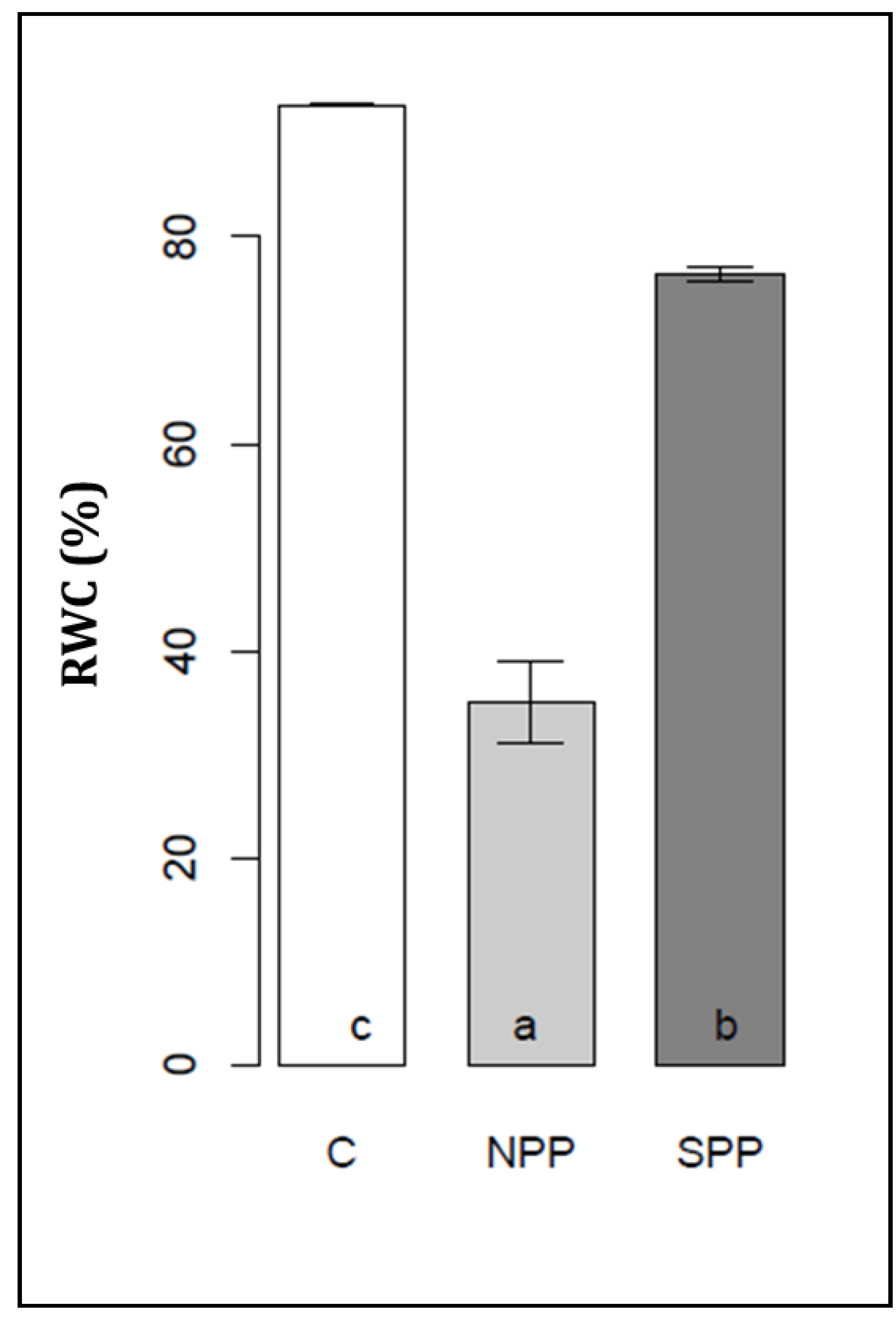
3.1. Changes in Seedlings Biomass and Relative Water Content
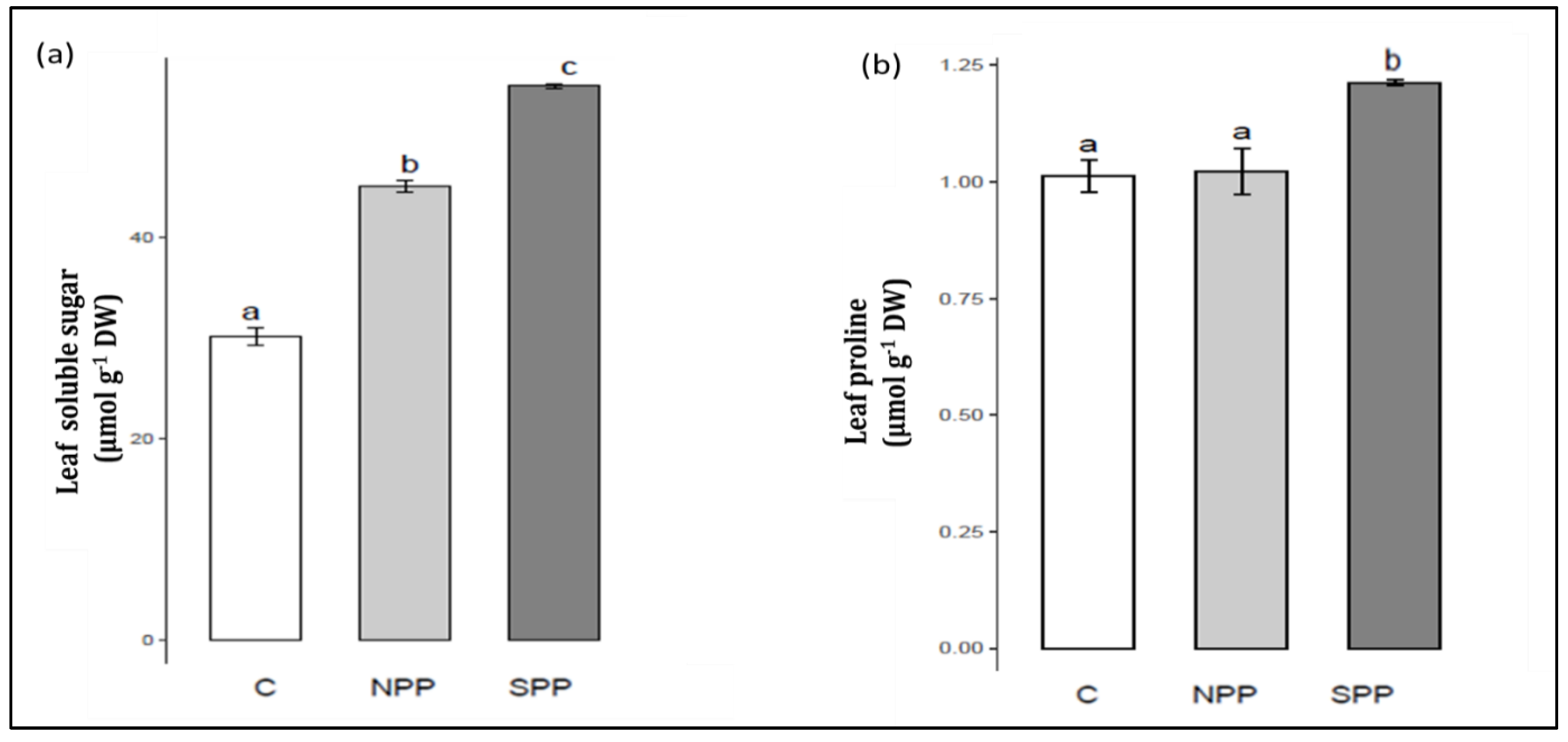
3.2. Changes in Osmoticums
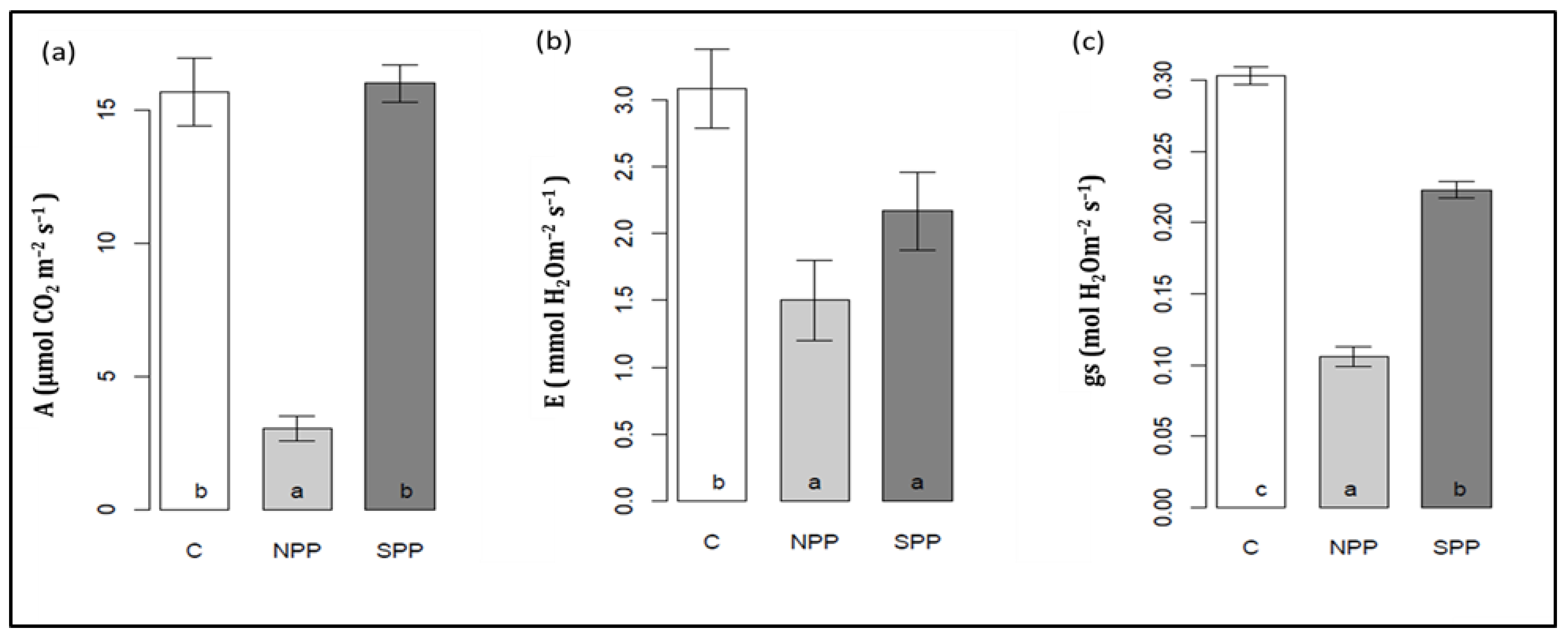
3.3. Changes in Photosynthesis Parameters
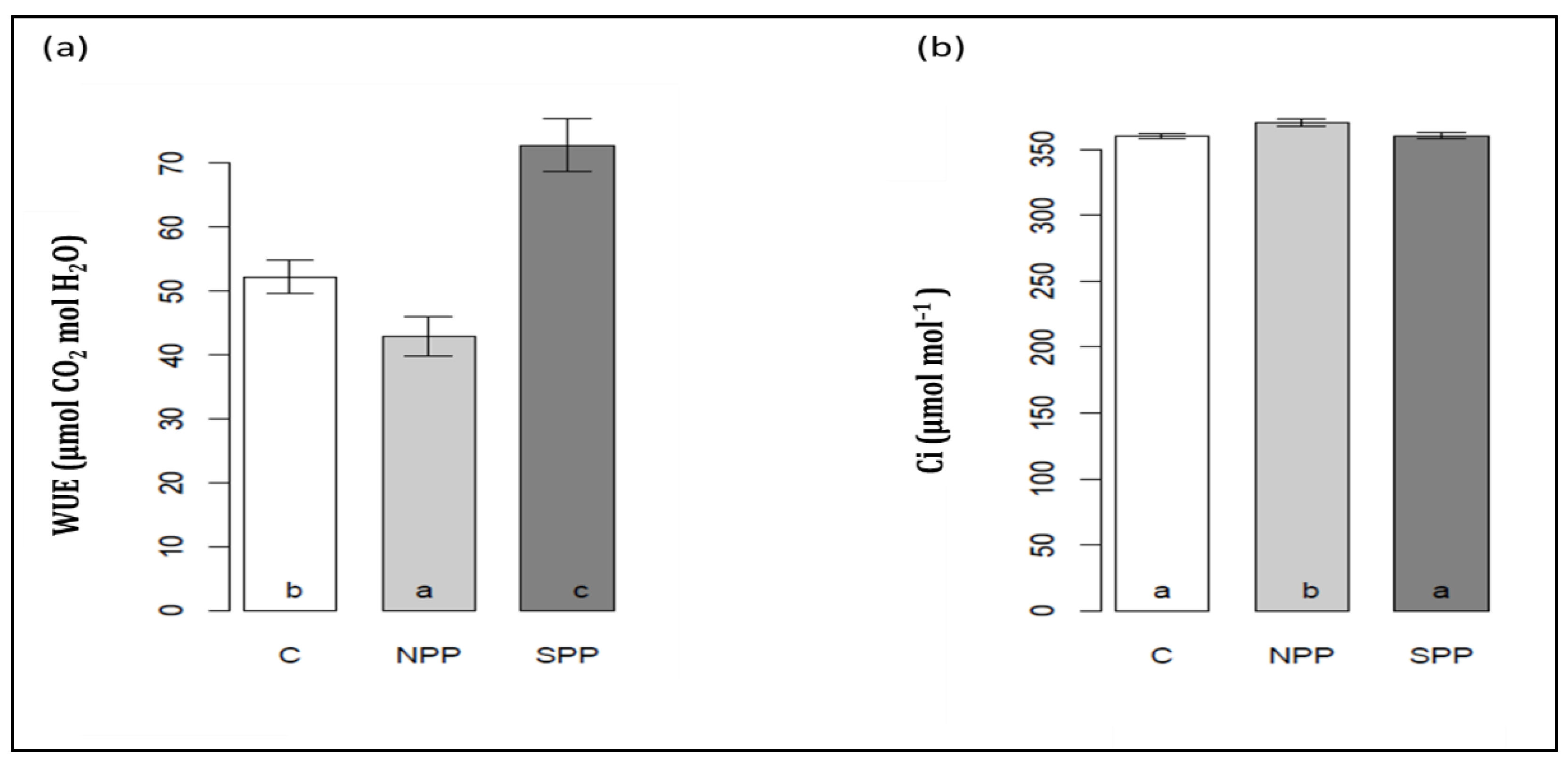
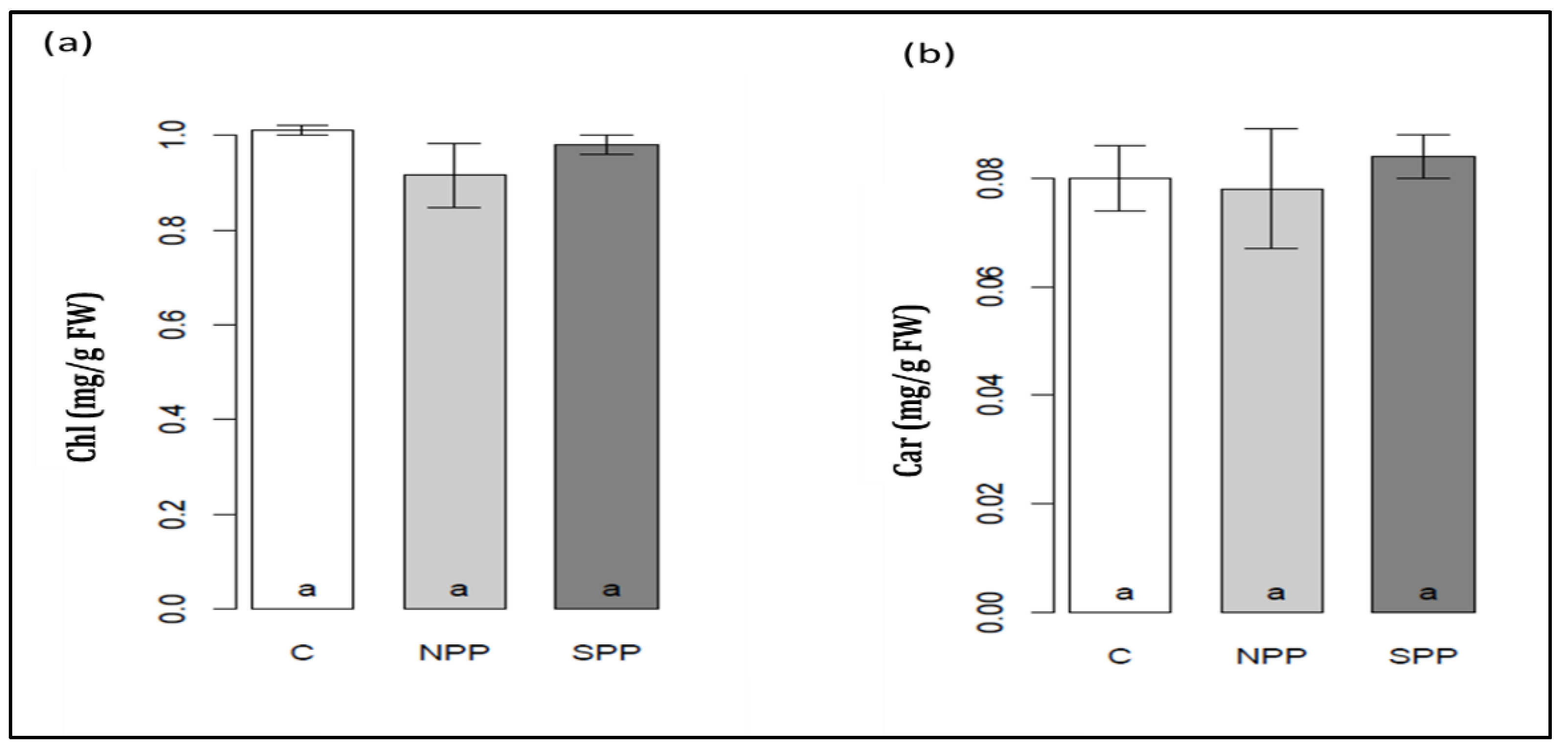
3.4. Changes in Leaf Density, Intercellular CO2 Assimilation, Water-Use Efficiency, Photosynthetic Pigments, and PSII Photochemistry
3.5. Effect on Lipid Metabolism
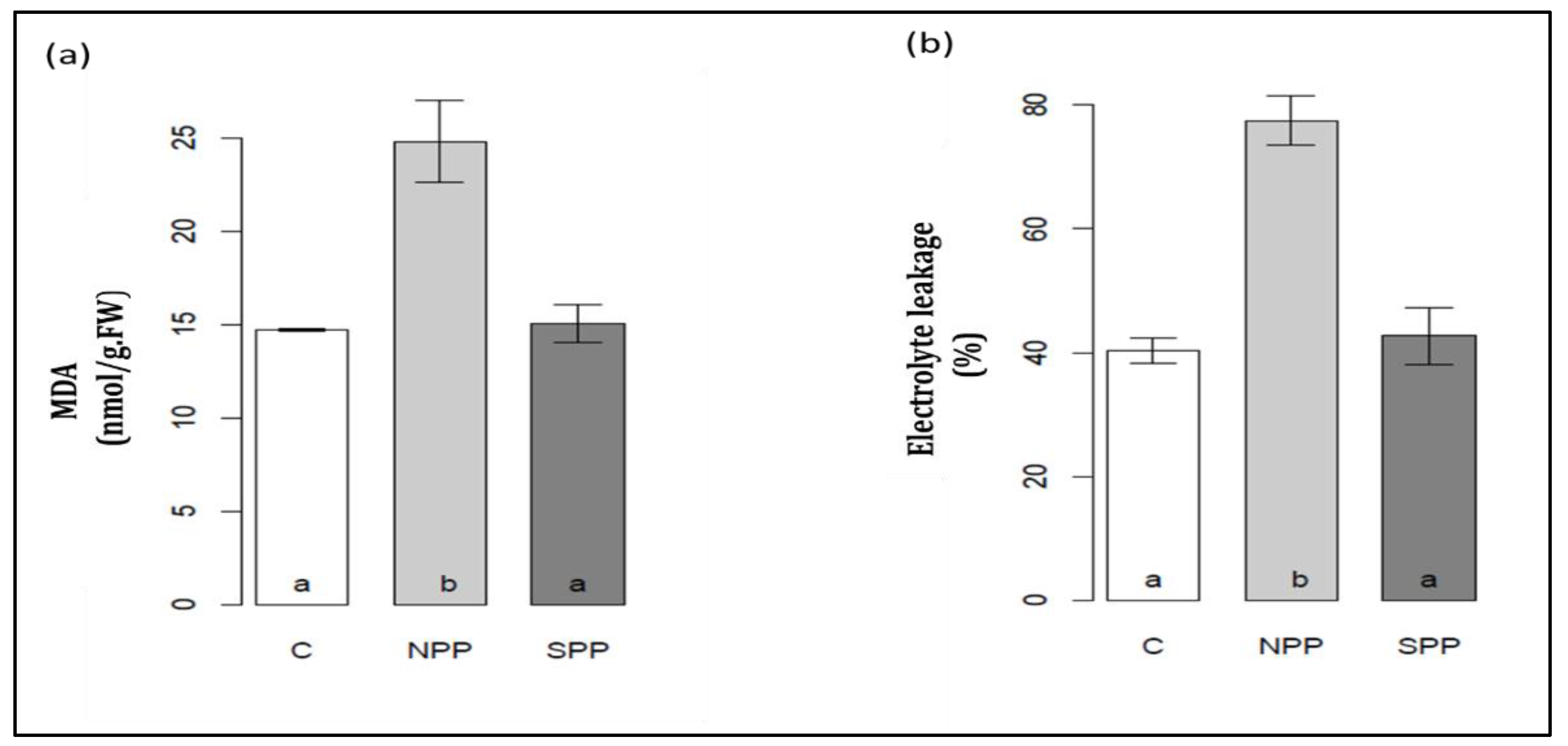
3.6. Changes in MDA Contents and Electrolyte Leakage
3.7. Changes in Antioxidant Enzymes Activity
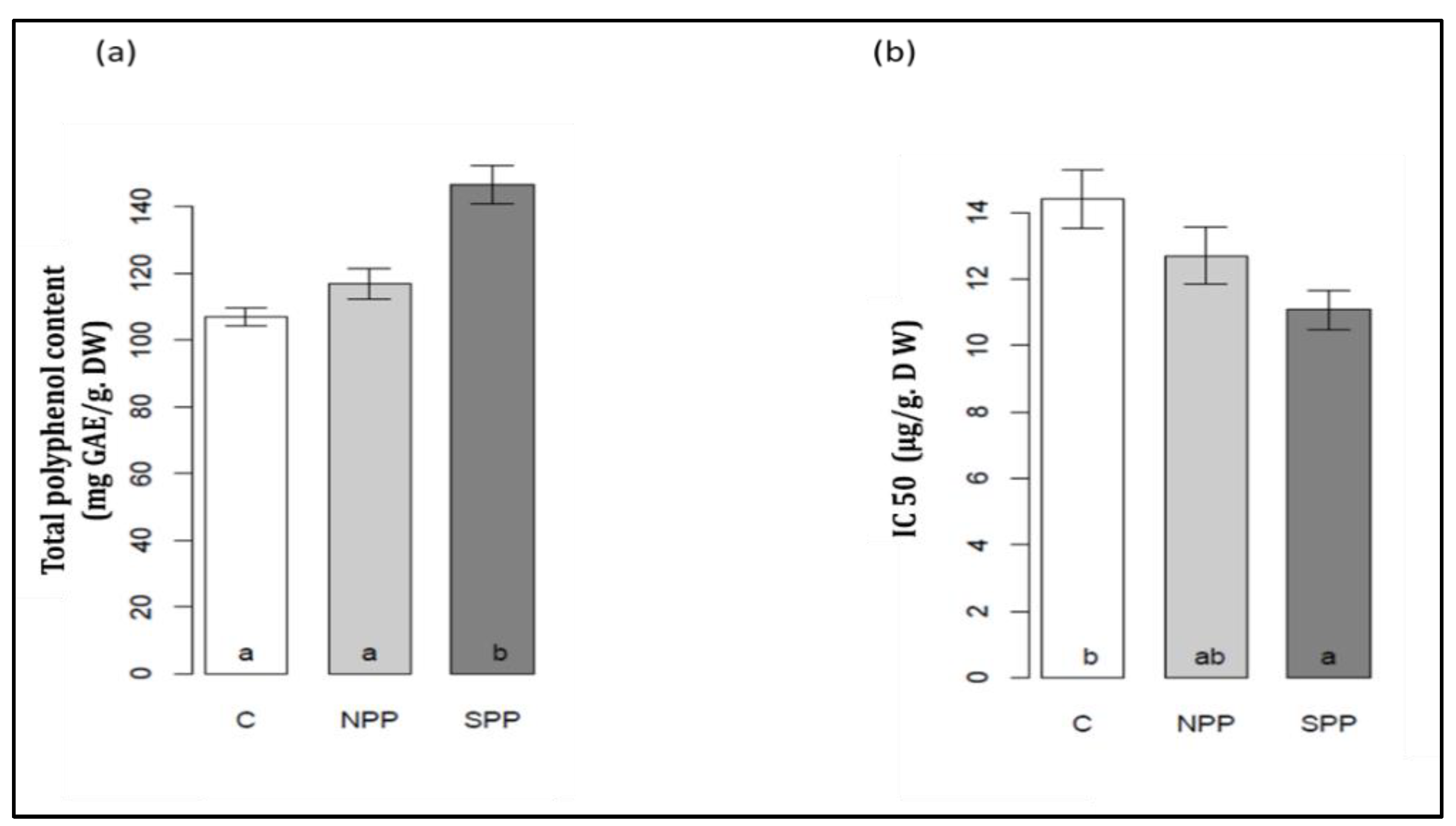
3.8. Changes in Phenolic Content and on Antioxidant Activity (IC50)
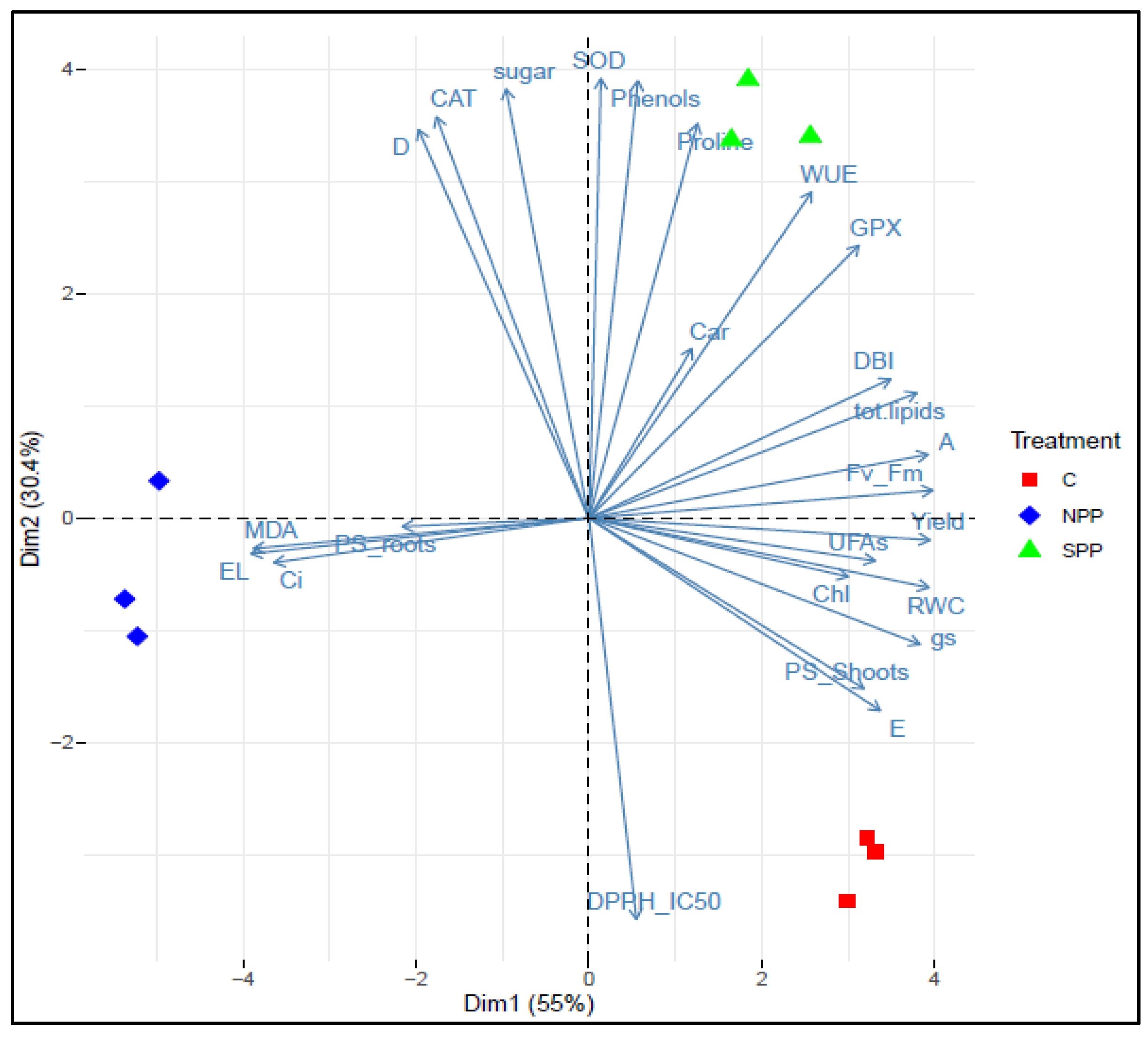
3.9. PCA Analysis
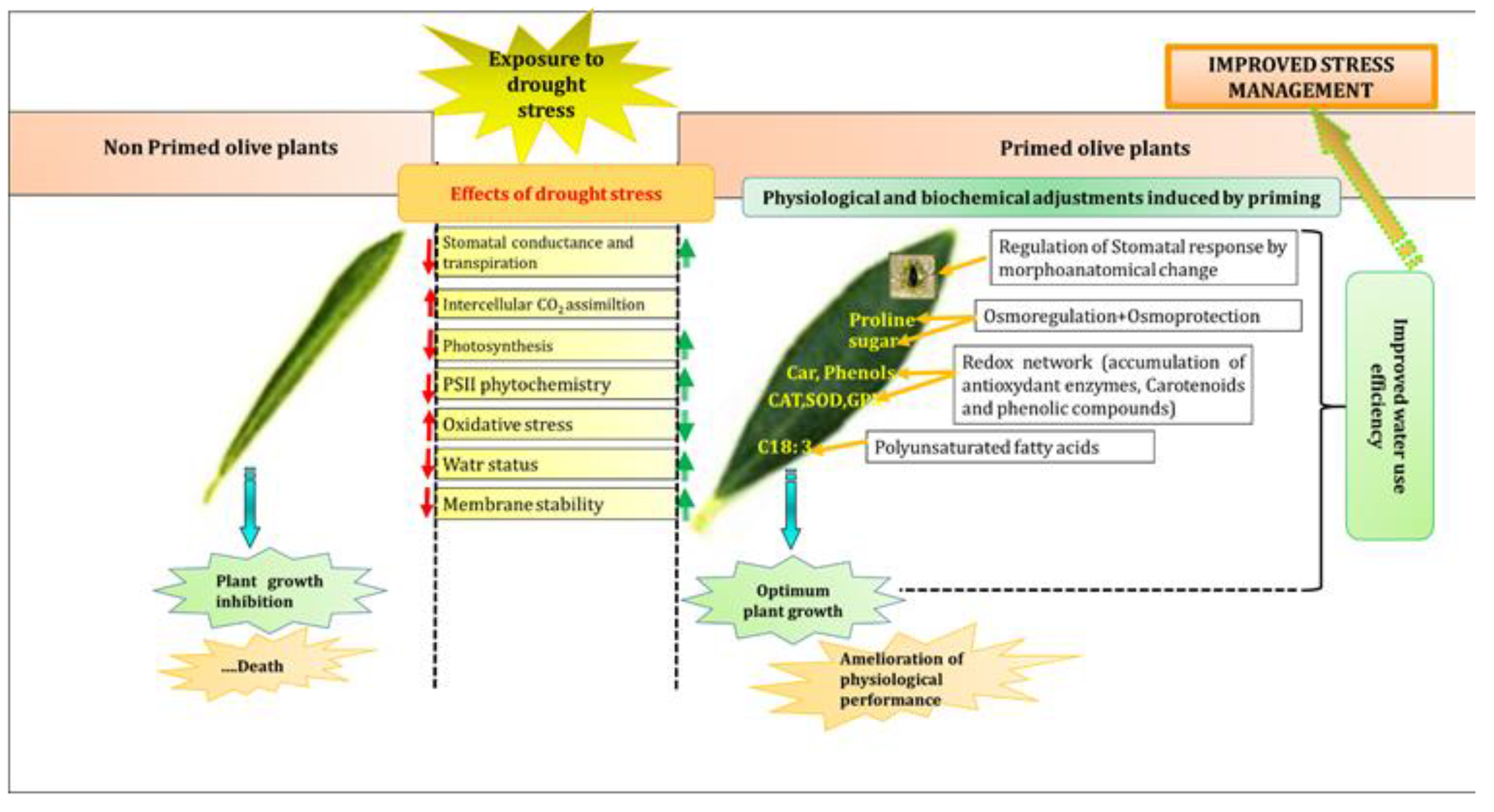
4. Discussion
4.1. Improvement of Growth, Water Homeostasis, Photosynthetic Activity, Structural Membrane Lipids Contents, and ROS Elimination Confirming the Beneficial Effects of Salt Priming to Mitigate Subsequent Drought Damages in Olive Seedlings
4.2. Physiological and Metabolic Adjustments Induced by Cross-Priming under Subsequent Drought in Young Olive Seedlings
4.2.1. Cross-Priming Induced Accumulation of Osmoticums and Morphofunctional Mechanisms in Olive Leaves
4.2.2. Cross-Priming Allowed to Olive Plants to Maintain Their Structural Lipid Contents as Well as Their Fatty Acid Composition to Protect the Photosystem Functionality
4.2.3. Cross-Priming Induces Antioxidant Defense to Survive Drought Stress
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and salinity stress responses and microbe-induced tolerance in plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Quan, W.; Bartels, D. Stress memory responses and seed priming correlate with drought tolerance in plants: An overview. Planta 2022, 255, 45. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.F.; Santos, F.L. Yield and olive oil characteristics of a low-density orchard (cv. Cordovil) subjected to different irrigation regimes. Agric. Water Manag. 2010, 97, 363–373. [Google Scholar] [CrossRef]
- Nardini, A.; Lo, M.A.; Trifilò, P.; Salleo, S. The challenge of the Mediterranean climate to plant hydraulics: Response and adaptations. Environ. Exp. Bot. 2014, 97, 30–39. [Google Scholar] [CrossRef]
- Silva, S.; Santos, C.; Serodio, J.; Silva, A.M.; Dias, M.C. Physiological performance of drought-stressed olive plants when exposed to a combined heat–UV-B shock and after stress relief. Funct. Plant Biol. 2018, 45, 1233–1240. [Google Scholar] [CrossRef]
- Ben Abdallah, M.; Trupiano, D.; Polzella, A.; De Zio, E.; Sassi, M.; Scaloni, A.; Zarrouk, M.; Ben Youssef, N.; Scippa, G.S. Unraveling physiological, biochemical and molecular mechanisms involved in olive (Olea europaea L. cv. Chétoui) tolerance to drought and salt stresses. J. Plant Physiol. 2018, 220, 83–95. [Google Scholar] [CrossRef]
- Ben Ahmed, C.; Ben Rouina, B.; Boukhris, M.M. Effects of water deficit on olive trees cv. Chemlali under field conditions in arid region in Tunisia. Sci. Hortic. 2007, 113, 267–277. [Google Scholar] [CrossRef]
- Proietti, P.; Antognozzi, E. Effect of irrigation on fruit quality of table olives, cultivar Ascolana tenera. N. Z. J. Crop Hortic. Sci. 1996, 24, 175–181. [Google Scholar] [CrossRef]
- Bartolni, S.; Leccese, A.; Andreini, L. Influence of canopy fruit location on morphological, histochemical and biochemical changes in two oil olive cultivars. Plant Biosyst. 2014, 148, 1221–1230. [Google Scholar] [CrossRef]
- Petridis, A.; Therios, I.; Samouris, G.; Koundouras, S.; Giannakoula, A. Effect of water deficit on leaf phenolic composition, gas exchange, oxidative damage and antioxidant activity of four Greek olive (Olea europaea L.) cultivars. Plant Physiol. Biochem. 2012, 60, 1–11. [Google Scholar] [CrossRef]
- Martinelli, F.; Remorini, D.; Saia, S.; Massai, R.; Tonutti, P. Metabolic profiling of ripe olive fruit in response to moderate water stress. Sci. Hortic. 2013, 159, 52–58. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, B. Drought priming-induced heat tolerance: Metabolic pathways and molecular mechanisms. In Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Academic Press: Cambridge, MA, USA, 2020; pp. 149–160. [Google Scholar] [CrossRef]
- Tippmann, H.F.; Schlüter, U.; Collinge, D.B.; Teixeira DaSilva, J.A. Common themes in biotic and abiotic stress signalling in plants. Floric.Ornam. Plant Biotechnol. 2006, 3, 52–67. [Google Scholar]
- Bowler, C.; Fluhr, R. The role of calcium and activated oxygens as signals for controlling cross-tolerance. Trends Plant Sci. 2000, 5, 1360–1385. [Google Scholar] [CrossRef] [PubMed]
- Katam, R.; Shokri, S.; Murthy, N.; Singh, S.K.; Suravajhala, P.; Khan, M.N.; Bahmani, M.; Sakata, K.; Reddy, K.R. Proteomics, physiological, and biochemical analysis of cross tolerance mechanisms in response to heat and water stresses in soybean. PLoS ONE 2020, 15, e0233905. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Able, A.J.; Able, J.A. Priming crops for the future: Rewiring stress memory. Trends Plant Sci. 2022, 27, 699–716. [Google Scholar] [CrossRef]
- Perincherry, L.; Stępień, Ł.; Vasudevan, S.E. Cross-Tolerance and Autoimmunity as Missing Links in Abiotic and Biotic Stress Responses in Plants: A Perspective toward Secondary Metabolic Engineering. Int. J. Mol. Sci. 2021, 22, 11945. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. Cross-stress tolerance and stress “memory” in plants. Environ. Exp. Bot. 2013, 94, 1–88. [Google Scholar] [CrossRef]
- Walter, J.; Jentsch, A.; Beierkuhnlein, C.; Kreyling, J. Ecological stress memory and cross stress tolerance in plants in the face of climate extremes. Environ. Exp. Bot. 2013, 94, 3–8. [Google Scholar] [CrossRef]
- Hussain, M.; Farooq, M.; Sattar, A.; Ijaz, M.; Sher, A.; Ul-Allah, S. Mitigating the adverse effects of drought stress through seed priming and seed quality on wheat (Triticum aestivum L.) productivity. Pak. J. Agric. Sci. 2018, 55, 313–319. [Google Scholar]
- Zandalinas, S.I.; Sengupta, S.; Fritschi, F.B.; Azad, R.K.; Nechushtai, R.; Mittler, R. The impact of multifactorial stress combination on plant growth and survival. New Phytol. 2021, 230, 1034–1048. [Google Scholar] [CrossRef]
- Gong, M.; Chen, B.; Li, Z.; Guo, L. Heat-shock-induced cross adaptation to heat, chilling, drought and salt stress in maize seedlings and involvement of H2O2. J. Plant Physiol. 2001, 158, 1125–1130. [Google Scholar] [CrossRef]
- Wang, L.J.; Huang, W.D.; Li, J.Y.; Liu, Y.F.; Shi, Y.L. Peroxidation of membrane lipid and Ca2+ homeostasis in grape mesophyll cells during the process of cross adaptation to temperature stresses. Plant Sci. 2004, 167, 71–77. [Google Scholar] [CrossRef]
- Kuznetsov, V.V.; Rakitin, V.Y.; Zholkevich, V.N. Effects of preliminary heat-shock treatment on accumulation of osmolytes and drought resistance in cotton plants during water deficiency. Physiol. Plant. 1999, 107, 399–406. [Google Scholar] [CrossRef]
- Saini, R.; Adhikary, A.; Nayyar, H.; Kumar, S. Cross-priming accentuates key biochemical and molecular indicators of defense and improves cold tolerance in chickpea (Cicer arietinum L.). Acta Physiol. Plant. 2019, 41, 181. [Google Scholar] [CrossRef]
- Li, X.; Topbjerg, H.B.; Jiang, D.; Liu, F. Drought priming at vegetative stage improves the antioxidant capacity and photosynthesis performance of wheat exposed to a short-term low temperature stress at jointing stage. Plant Soil 2015, 393, 307–318. [Google Scholar] [CrossRef]
- Yang, X.; Zou, F.; Zhang, Y.; Shi, J.; Qi, M.; Liu, Y.; Li, T. NaCl Pretreatment Enhances the Low Temperature Tolerance of Tomato Through Photosynthetic Acclimation. Front. Plant Sci. 2022, 1959. [Google Scholar] [CrossRef]
- Pushpavalli, R.; Berger, J.D.; Turner, N.C.; Siddique, K.H.; Colmer, T.D.; Vadez, V. Cross-tolerance for drought, heat and salinity stresses in chickpea (Cicer arietinum L.). J. Agron. Crop Sci. 2020, 206, 405–419. [Google Scholar] [CrossRef]
- Feng, X.C.; Sun, Y.L.; Bai, X.F. Salinity induced cross tolerance to drought in cotton seedlings is associated with reduced lipid peroxidation and photoprotection through activation of antioxidant system. Int. J. Agric. Biol. 2019, 21, 338–344. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Chen, H.; Huang, J.; Zhang, Y.; Qi, M.; Liu, Y.; Li, T. Photosynthetic response mechanism of soil salinity-induced cross-tolerance to subsequent drought stress in tomato plants. Plants 2020, 9, 363. [Google Scholar] [CrossRef] [Green Version]
- Isayenkov, S.V. Genetic sources for the development of salt tolerance in crops. Plant Growth Regul. 2019, 89, 1–17. [Google Scholar] [CrossRef]
- Yadav, R.; Juneja, S.; Kumar, S. Cross priming with drought improves heat-tolerance in chickpea (Cicer arietinum L.) by stimulating small heat shock proteins and antioxidative defense. Environ. Sustain. 2021, 4, 171–182. [Google Scholar] [CrossRef]
- Guerfel, M.; Baccouri, O.; Boujnah, D.; Chaibi, W.; Zarrouk, M. Impacts of water stress on gas exchange water relations, chlorophyll content and leaf structure in the two main Tunisian olive (Olea europaea L.) cultivars. Sci. Hortic. 2009, 11, 257–263. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plants extracts by anthrone. Biochem. J. 1954, 90, 508–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Abdallah, M.; Methenni, K.; Nouairi, I.; Zarrouk, M.; Ben Youssef, N. Drought priming improves subsequent more severe drought in a drought-sensitive cultivar of olive cv. Chétoui. Sci. Hortic. 2017, 221, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I: Kinetic and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilising the principle of protein-dyebinding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Skerget, M.; Kotnik, P.; Hadolin, M.; Hras, A.R.; Simoni, M.; Knez, Z. Phenols, proanthocyanidins, flavones and flavonols in some plant materials and their antioxidant activities. Food Chem. 2005, 89, 191–198. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Metcalfe, D.; Schmitz, A.; Pelka, J.R. Rapid preparation of fatty acid esters from lipids for gas chromatographic analysis. Anal. Chem. 1966, 38, 524–535. [Google Scholar] [CrossRef]
- Nouairi, I.; Methenni, K.; Mhadhbi, H.; Jbara, M. Effects of CaCl2 pretreatment on antioxidant enzyme and leaf lipid content of faba bean (Vicia faba L.) seedlings under cadmium stress. Plant Growth Regul. 2012, 68, 37–47. [Google Scholar] [CrossRef]
- Trabelsi, L.; Gargouri, K.; Hassena, A.B.; Mbadra, C.; Ghrab, M.; Ncube, B.; Van Staden, J.; Gargouri, R. Impact of drought and salinity on olive water status and physiological performance in an arid climate. Agric. Water Manag. 2019, 213, 749–759. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Vé ronique, S., Alberola, C., Eds.; Springer: Amsterdam, The Netherlands, 2009; pp. 153–188. [Google Scholar]
- Dias, M.C.; Correia, C.; Serôdio, J.; Silva, A.M.S.; Freitas, H.; Santos, C. Chlorophyll fluorescence and oxidative stress endpoints to discriminate olive cultivars tolerance to drought and heat episodes. Sci. Hortic. 2018, 231, 31–35. [Google Scholar] [CrossRef]
- Guerfel, M.; Baccouri, O.; Boujnah, D.; Zarrouk, M. Changes in lipid composition, water relations and gas exchange in leaves of two young ‘Chemlali’ and ‘Chetoui’ olive trees in response to water stress. Plant Soil 2008, 311, 121–129. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Sofo, A.; Dichio, B.; Xiloyannis, C.; Masia, A. Lipoxygenase activity and proline accumulation in leaves and roots of olive tree in response to drought stress. Physiol. Plant. 2004, 121, 58–65. [Google Scholar] [CrossRef]
- Ben Youssef, N.; Al-Moaikal, R.M.S.; Al-Hawas, G.H.S.; Alrammah, F.M. Up-modulation of membrane lipid composition and functionality by seed priming under salinity in the Hasawi rice variety. J. Appl. Bot. Food Qual. 2020, 93, 159–167. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Pinior, A.; Grunewaldt-Stöcker, G.; Von Alten, H.; Strasser, R.J. Mycorrhizal impact on drought stress tolerance of rose plants probed by chlorophyll a fuorescence, proline content and visual scoring. Mycorrhiza 2005, 15, 596–605. [Google Scholar] [CrossRef]
- Jiang, Y.; Huang, B. Osmotic adjustment and root growth associated with drought preconditioning enhanced heat tolerance in Kentucky bluegrass. Crop Sci. 2001, 41, 1168–1173. [Google Scholar] [CrossRef]
- Ennajeh, M.; Tounekti, T.; Vadel, A.M.; Khemira, H.; Cochard, H. Water relations and drought-induced embolism in two olive (Olea europaea L.) varieties ‘Meski’ and ‘Chemlali’ under severe drought conditions. Tree Physiol. 2008, 28, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Chartzoulakis, K.; Patakas, A.; Bosabalidis, A.M. Changes in water relations, photosynthesis and leaf anatomy induced by intermittent drought in two olive cultivars. Environ. Exp. Bot. 1999, 42, 113–120. [Google Scholar] [CrossRef]
- Evans, J.R.; Loreto, F. Acquisition and diffusion of CO2 in higher plant leaves. In Photosynthesis: Physiology and Metabolism; Leegood, R.C., Sharkey, T.D., von Caemmerer, S., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 321–351. [Google Scholar]
- Singha, A.; Soothar, R.K.; Wang, C.; Marín, E.E.T.; Tankari, M.; Hao, W.; Wang, Y. Drought priming alleviated salinity stress and improved water use efficiency of wheat plants. Plant Growth Regul. 2022, 96, 357–368. [Google Scholar] [CrossRef]
- Veneklaas, E.J.; Silva, M.S.; den Ouden, F. Determinants of growth rate in Ficus benjamina L. compared to related faster-growing woody and herbaceous species. Sci. Hortic. 2002, 93, 75–84. [Google Scholar] [CrossRef]
- Niinemets, U. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology 2001, 82, 453–469. [Google Scholar] [CrossRef]
- Hossain, M.A.; Li, Z.G.; Hoque, T.S.; Burritt, D.J.; Fujita, M.; Munné-Bosch, S. Heat or cold priming-induced cross-tolerance to abiotic stresses in plants: Key regulators and possible mechanisms. Protoplasma 2018, 255, 399–412. [Google Scholar] [CrossRef]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Kanervo, E.; Aro, E.M.; Murata, N. Low unsaturation level of thylakoid membrane lipids limits turnover of the D1 protein of photosystem II at high irradiance. FEBS Lett. 1995, 364, 239–242. [Google Scholar] [CrossRef] [Green Version]
- Quinn, P.J.; Willliams, W.P. The structural role of lipids in photosynthetic membranes. Biochim. Biophys. Acta 1983, 737, 223–266. [Google Scholar] [CrossRef]
- Gounaris, K.; Mannock, D.A.; Sen, A.; Brain, A.P.R.; Williams, W.P.; Quinn, P.J. Polyunsaturated fatty acyl residues of galactolipids are involved in the control of bilayer/ non bilayer lipid transitions in higher plant chloroplasts. Biochim. Biophys. Acta 1983, 732, 229–242. [Google Scholar] [CrossRef]
- Zhang, C.; Tian, S. Peach fruit acquired tolerance to low temperature stress by accumulation of linolenic acid and N-acylphosphatidylethanolamine in plasma membrane. Food Chem. 2010, 120, 864–872. [Google Scholar] [CrossRef]
- Peng, Y.; Xu, C.; Xu, L.; Huang, B. Improved heat tolerance through drought preconditioning associated with changes in lipid composition, antioxidant enzymes, and protein expression in Kentucky bluegrass. Crop Sci. 2012, 52, 807–817. [Google Scholar] [CrossRef]
- Wang, J.; Juliani, H.R.; Jespersen, D.; Huang, B. Differential profiles of membrane proteins, fatty acids, and sterols associated with genetic variations in heat tolerance for a perennial grass species, hard fescue (Festuca trachyphylla). Environ. Exp. Bot. 2017, 140, 65–75. [Google Scholar] [CrossRef]
- Hossain, M.A.; Mostofa, M.G.; Fujita, M. Heat-shock positively modulates oxidative protection of salt and drought-stressed mustard (Brassica campestris L.) seedlings. J. Plant Sci. Mol. Breed. 2013, 2, 2. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Fv/Fm | Y | D |
|---|---|---|---|
| C | 0.79 ± 0.01 b | 0.76 ± 0.03 b | 435 ± 60 a |
| SPP | 0.75 ± 0.02 b | 0.65 ± 0.02 b | 853 ± 40 c |
| NPP | 0.13 ± 0.02 a | 0.20 ± 0.03 a | 769 ± 50 b |
| C14:0 | C14:1 | C16:0 | C16:1 | C18:0 | C18:1 | C18:2 | C18:3 | C20:0 |
|---|---|---|---|---|---|---|---|---|
| 0.60 ± 0.03 a | 1.10 ± 0.12 a | 17.70 ± 1.55 a | 0.80 ± 0.05 a | 3.00 ± 0.04 a | 20.49 ± 0.46 a | 10.50 ± 0.52 a | 40.00 ± 1.41 b | 3.20 ± 0.20 a |
| 0.78 ± 0.04 b | 0.87 ± 0.09 b | 20.01 ± 0.49 b | 0.94 ± 0.09 a | 2.88 ± 0.07 b | 19.44 ± 1.23 a | 10.57 ± 0.27 b | 41.73 ± 1.64 a | 2.76 ± 0.10 a |
| 1.74 ± 0.32 a | 2.47 ± 0.38 a | 21.48 ± 1.23 b | 0.93 ± 0.03 a | 4.16 ± 0.21 a | 19.20 ± 0.91 a | 11.51 ± 0.43 b | 35.30 ± 0.48 c | 3.19 ± 0.32 a |
| Treatment | CAT | SOD | GP |
|---|---|---|---|
| C | 2.60 ± 0.31 a | 119.20 ± 11.12 a | 3.17 ± 0.50 b |
| SPP | 8.08 ± 0.33 b | 187.73 ± 9.67 b | 5.28 ± 0.26 c |
| NPP | 6.97 ± 0.44 b | 143.50 ± 12.50 a | 1.18 ± 0.08 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben Abdallah, M.; Methenni, K.; Taamalli, W.; Hessini, K.; Ben Youssef, N. Cross-Priming Approach Induced Beneficial Metabolic Adjustments and Repair Processes during Subsequent Drought in Olive. Water 2022, 14, 4050. https://doi.org/10.3390/w14244050
Ben Abdallah M, Methenni K, Taamalli W, Hessini K, Ben Youssef N. Cross-Priming Approach Induced Beneficial Metabolic Adjustments and Repair Processes during Subsequent Drought in Olive. Water. 2022; 14(24):4050. https://doi.org/10.3390/w14244050
Chicago/Turabian StyleBen Abdallah, Mariem, Kawther Methenni, Wael Taamalli, Kamel Hessini, and Nabil Ben Youssef. 2022. "Cross-Priming Approach Induced Beneficial Metabolic Adjustments and Repair Processes during Subsequent Drought in Olive" Water 14, no. 24: 4050. https://doi.org/10.3390/w14244050
APA StyleBen Abdallah, M., Methenni, K., Taamalli, W., Hessini, K., & Ben Youssef, N. (2022). Cross-Priming Approach Induced Beneficial Metabolic Adjustments and Repair Processes during Subsequent Drought in Olive. Water, 14(24), 4050. https://doi.org/10.3390/w14244050