
Multivariate Analysis of Rotifer Community and Environmental Factors Using the Decomposed Components Extracted from a Time Series

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
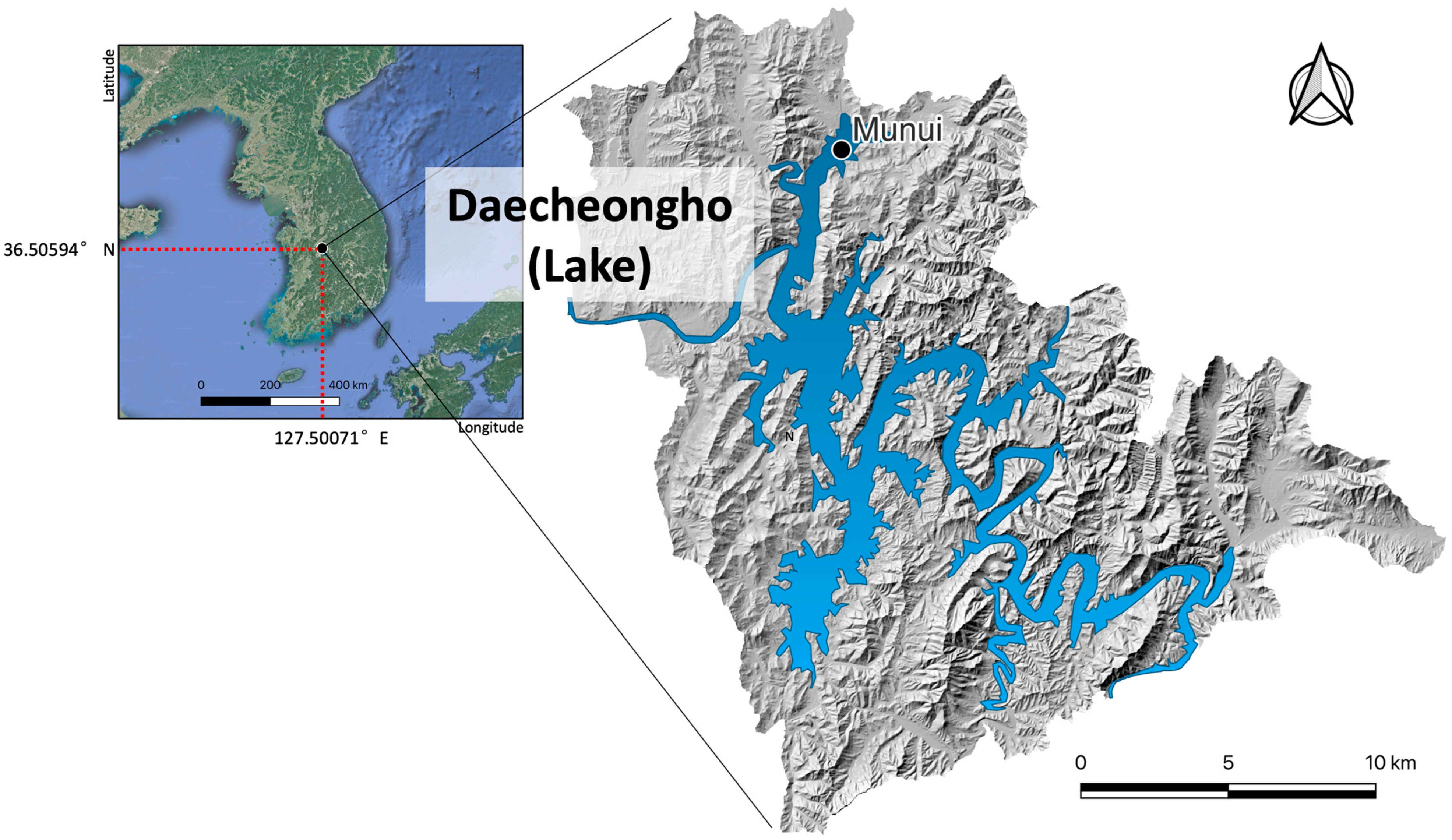
2.1. Study Site
2.2. Water Sampling and Measurement of Abiotic Factors
2.3. Plankton Sampling
2.4. Analyzed Factors
2.4.1. Rotifer Groups
2.4.2. Environmental Factors
2.5. Statistical Analysis
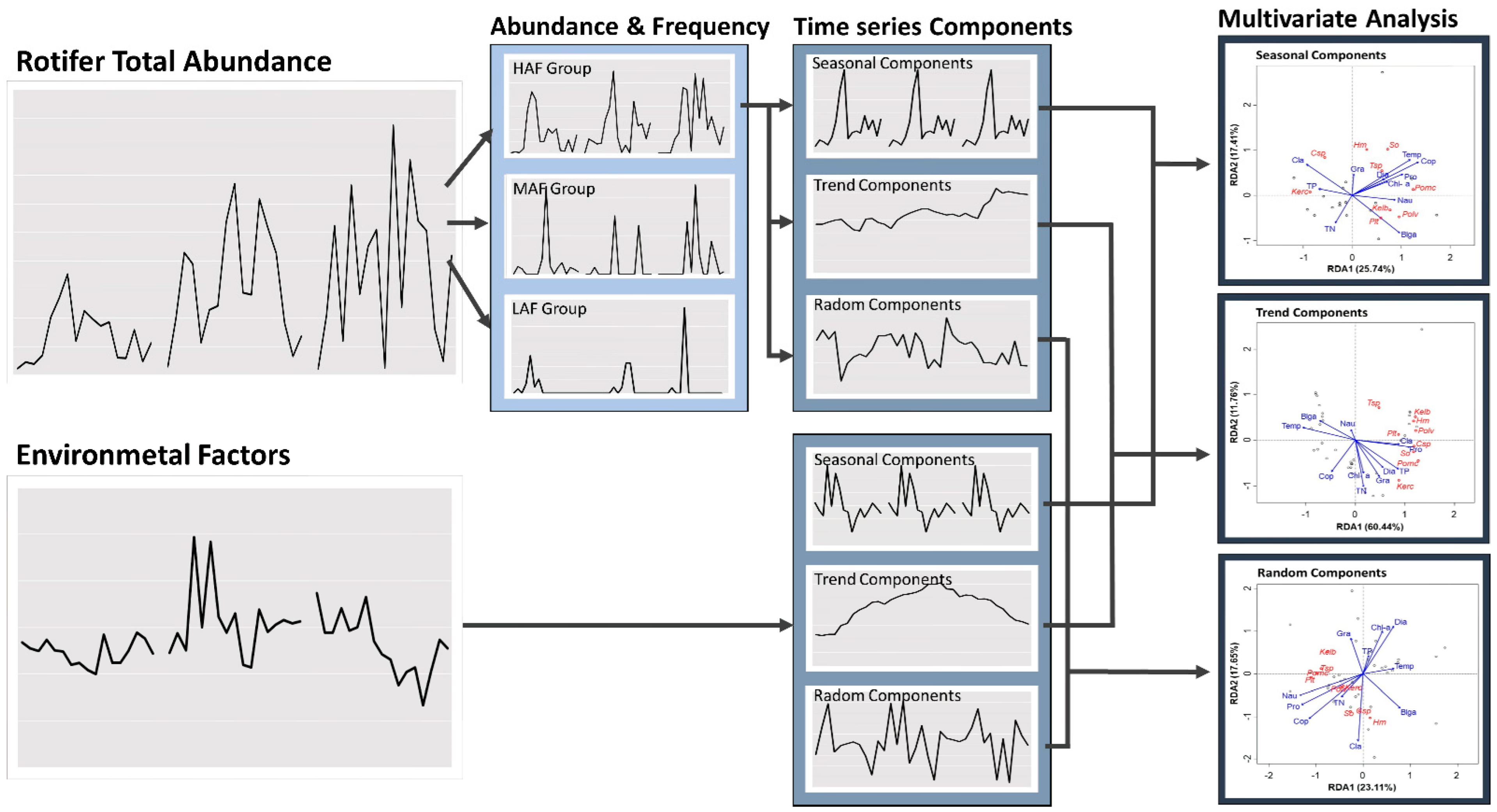
2.5.1. Decomposition Procedure
2.5.2. Multivariate Analysis Based on Decomposed Components
3. Results
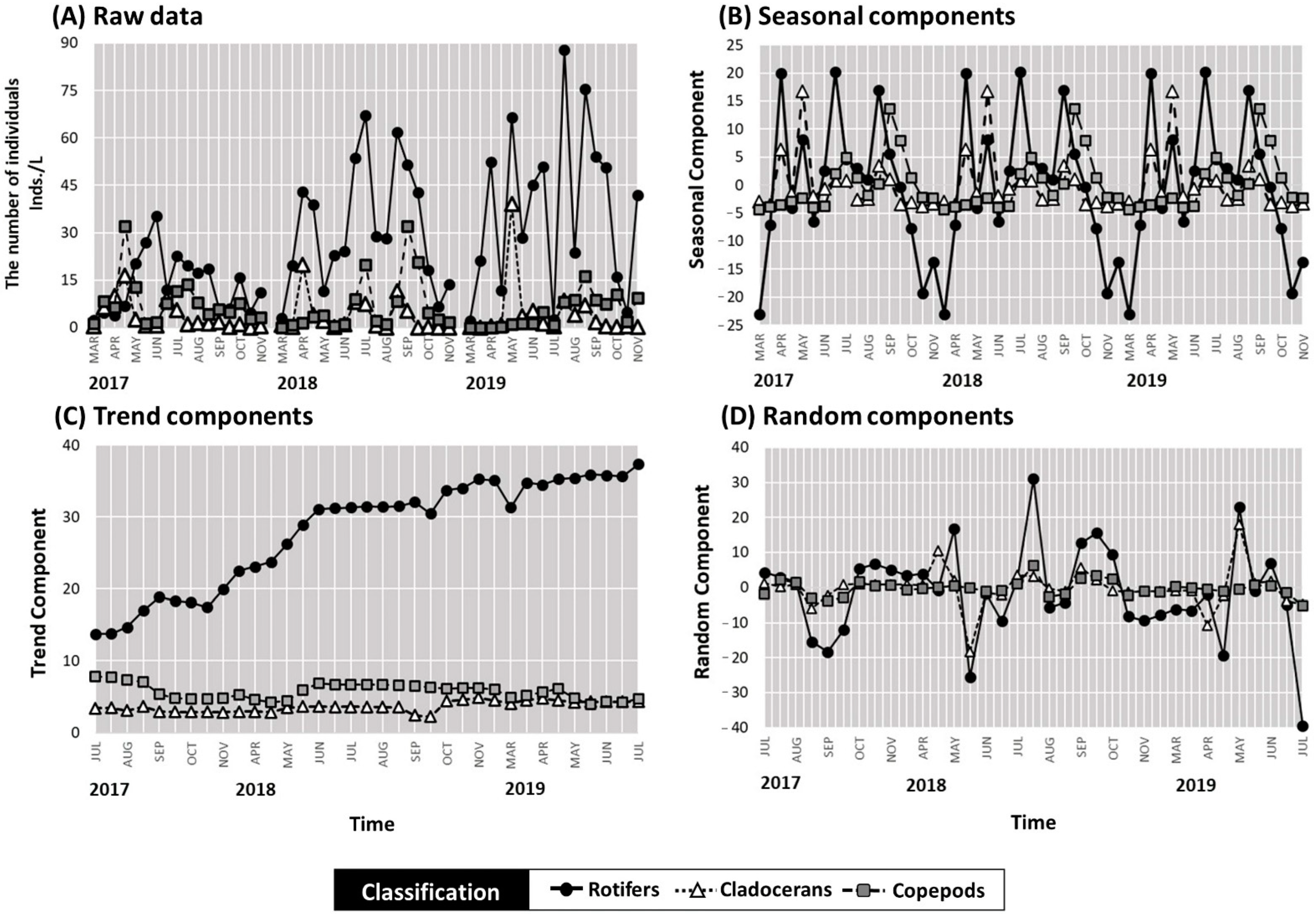
3.1. Variability in the Abundance of Zooplankton Groups and Results of the Time Series Analysis
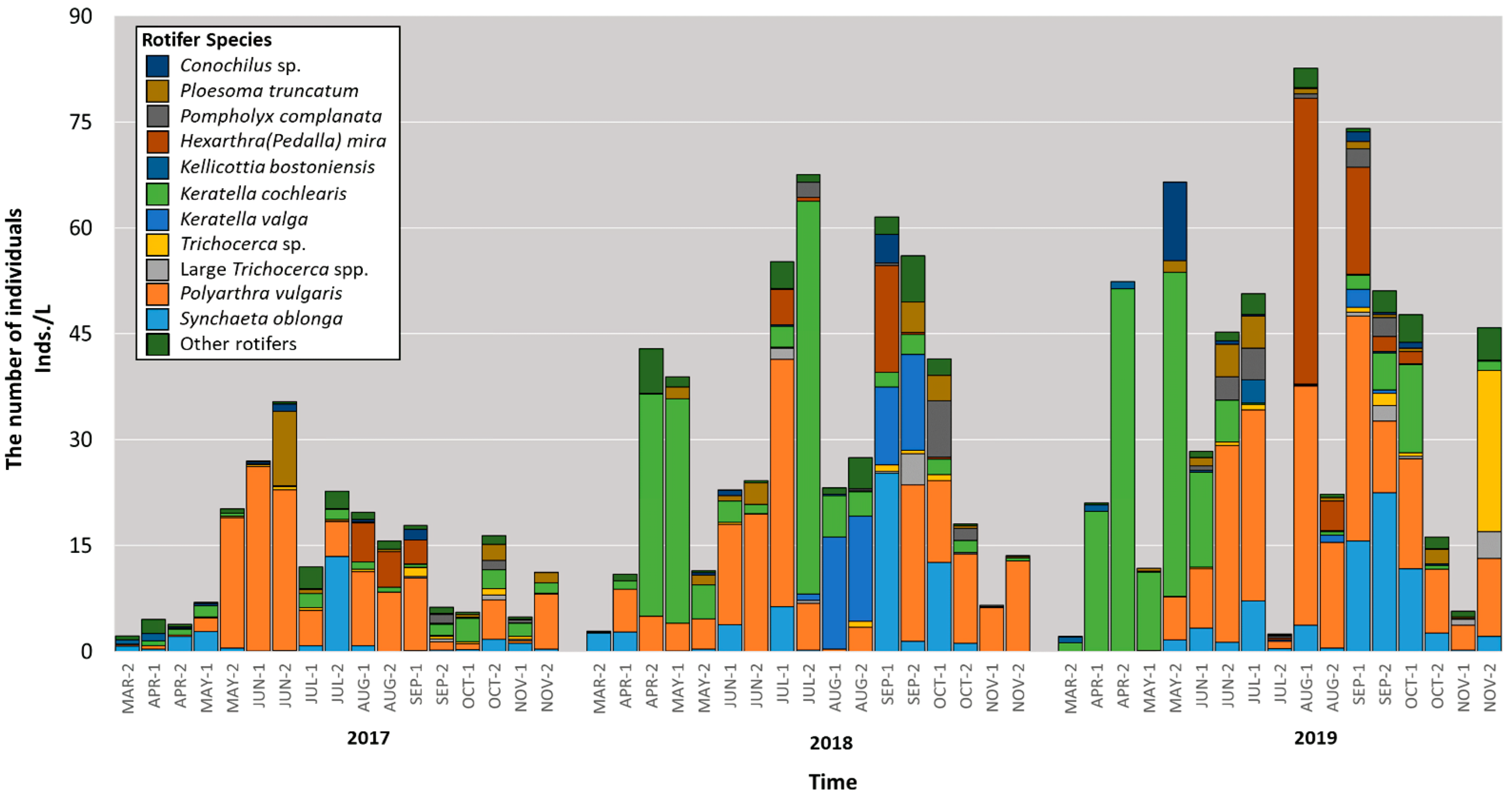
3.2. Abundance-Based Classification of Rotifers
3.3. RDA of Rotifer Groups Based on Undecomposed Population Data
3.4. RDA of Rotifer Groups Based on the Seasonal Component
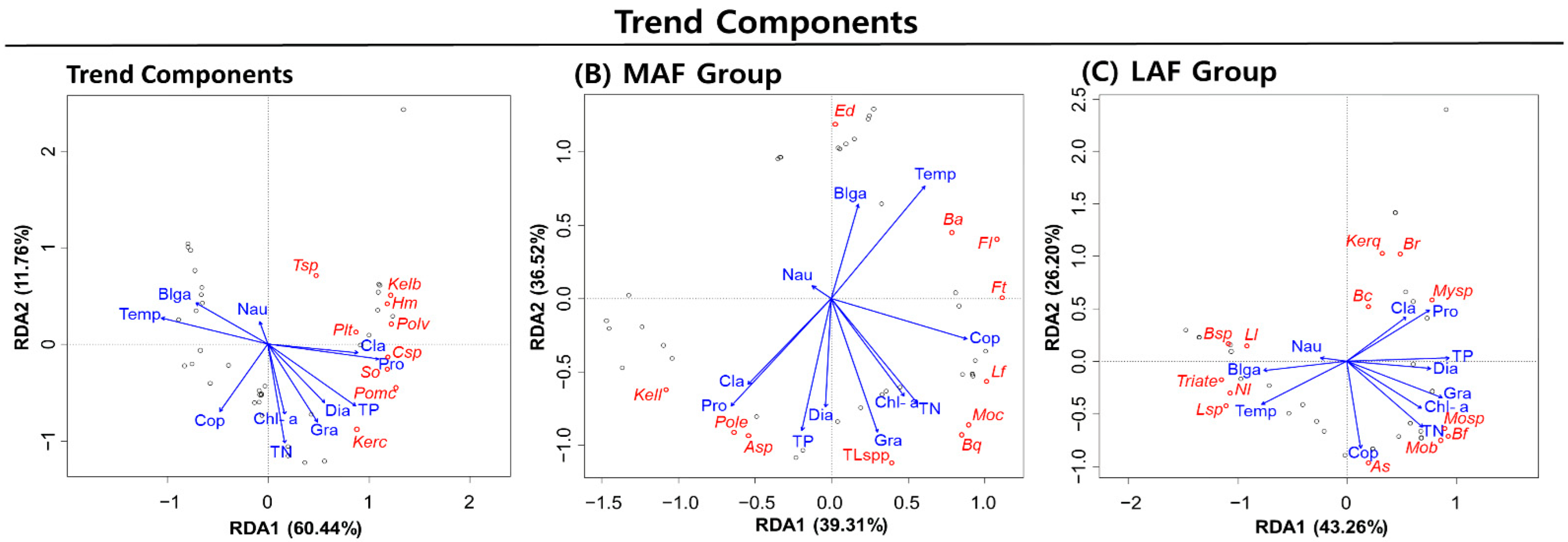
3.5. RDA of Rotifer Groups Based on the Trend Component
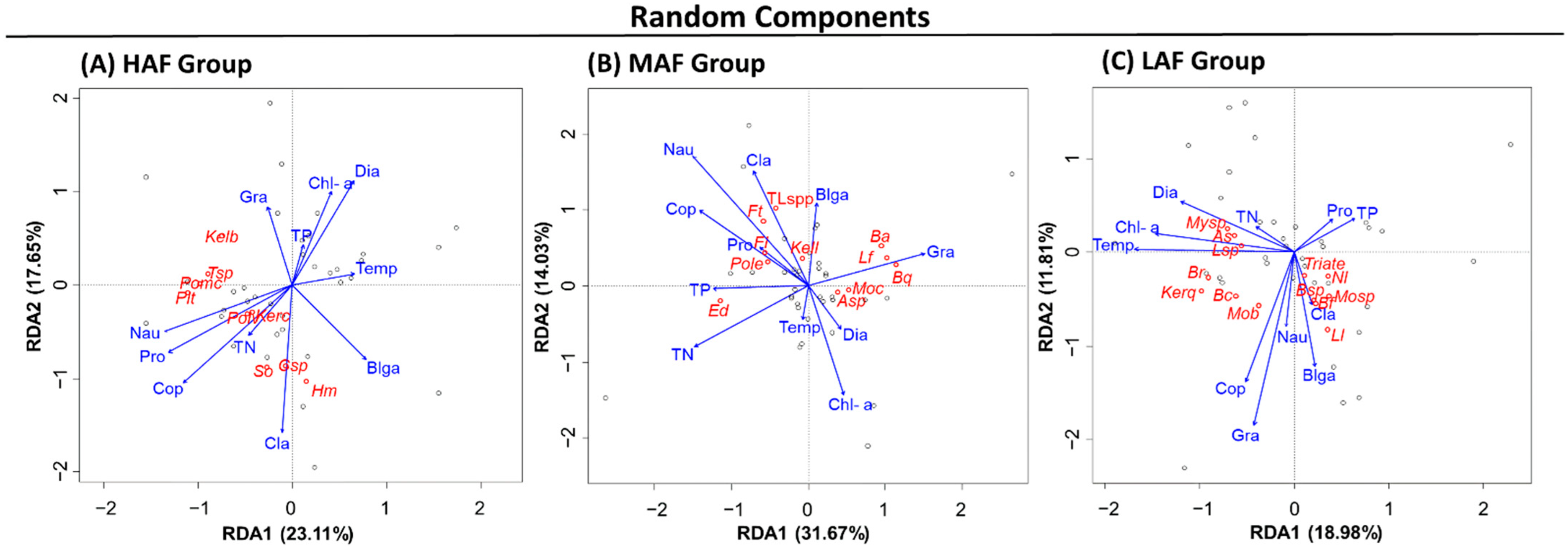
3.6. RDA of Rotifer Groups Based on the Random Component
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gilbert, J.J. Suppression of rotifer populations by Daphnia: A review of the evidence, the mechanisms, and the effects on zooplankton community structure. Limnol. Oceanogr. 1988, 33, 1286–1303. [Google Scholar] [CrossRef]
- Pace, M.L.; Findlay, S.E.G.; Lints, D. Zooplankton in advective environments: The Hudson River community and a comparative analysis. Can. J. Fish. Aquat. Sci. 1992, 49, 1060–1069. [Google Scholar] [CrossRef]
- Gannon, J.E.; Stemberger, R.S. Zooplankton (especially crustaceans and rotifers) as indicators of water quality. Trans. Am. Microsc. Soc. 1978, 97, 16–35. [Google Scholar] [CrossRef]
- Lampert, W.; Fleckner, W.; Rai, H.; Taylor, B.E. Phytoplankton control by grazing zooplankton: A study on the spring clear-water phase. Limnol. Oceanogr. 1986, 31, 478–490. [Google Scholar] [CrossRef]
- Shurin, J.B. Interactive effects of predation and dispersal on zooplankton communities. Ecology 2001, 82, 3404–3416. [Google Scholar]
- Fischer, J.; Visbeck, M. Seasonal variation of the daily zooplankton migration in the Greenland Sea. Deep Sea Res. I 1993, 40, 1547–1557. [Google Scholar] [CrossRef]
- Jose, R.; Sanalkumar, M.G. Seasonal variations in the zooplankton diversity of River Achencovil. Int. J. Sci. Res. Publ. 2012, 2, 1–5. [Google Scholar]
- Buckley, L.J.; Durbin, E.G. Seasonal and inter-annual trends in the zooplankton prey and growth rate of Atlantic cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) larvae on Georges Bank. Deep Sea Res. II 2006, 53, 2758–2770. [Google Scholar] [CrossRef]
- Allan, J.D. Life history patterns in zooplankton. Am. Nat. 1976, 110, 165–180. [Google Scholar] [CrossRef]
- Vargas, A.L.; Santangelo, J.M.; Bozelli, R.L. Recovery from drought: Viability and hatching patterns of hydrated and desiccated zooplankton resting eggs. Int. Rev. Hydrobiol. 2019, 104, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Fernando, C.H. The freshwater zooplankton of Sri Lanka, with a discussion of tropical freshwater zooplankton composition. Int. Rev. Gesamten. Hydrobiol. Hydrogr. 1980, 65, 85–125. [Google Scholar] [CrossRef] [Green Version]
- Shurin, J.B. Dispersal limitation, invasion resistance, and the structure of pond zooplankton communities. Ecology 2000, 81, 3074–3086. [Google Scholar]
- Mackas, D.L.; Greve, W.; Edwards, M.; Chiba, S.; Tadokoro, K.; Eloire, D.; Mazzocchi, M.G.; Batten, S.; Richardson, A.J.; Johnson, C.; et al. Changing zooplankton seasonality in a changing ocean: Comparing time series of zooplankton phenology. Prog. Oceanogr. 2012, 97–100, 31–62. [Google Scholar] [CrossRef]
- Nogueira, M.G. Zooplankton composition, dominance and abundance as indicators of environmental compartmentalization in Jurumirim Reservoir (Paranapanema River), São Paulo, Brazil. Hydrobiologia 2001, 455, 1–18. [Google Scholar] [CrossRef]
- Kosobokova, K.N.; Hopcroft, R.R.; Hirche, H.-J. Patterns of zooplankton diversity through the depths of the Arctic’s central basins. Mar. Biodivers. 2011, 41, 29–50. [Google Scholar] [CrossRef]
- Lindeque, P.K.; Parry, H.E.; Harmer, R.A.; Somerfield, P.J.; Atkinson, A. Next generation sequencing reveals the hidden diversity of zooplankton assemblages. PLoS ONE 2013, 8, e81327. [Google Scholar] [CrossRef]
- Vincent, K.; Mwebaza-Ndawula, L.; Makanga, B.; Nachuha, S. Variations in zooplankton community structure and water quality conditions in three habitat types in northern Lake Victoria. Lakes Reserv. Res. Manag. 2012, 17, 83–95. [Google Scholar] [CrossRef]
- Roman, M.R.; Dam, H.G.; Gauzens, A.L.; Urban-Rich, J.; Foley, D.G.; Dickey, T.D. Zooplankton variability on the equator at 140 W during the JGOFS EqPac study. Deep Sea Res. II 1995, 42, 673–693. [Google Scholar] [CrossRef]
- Clark, R.A.; Frid, C.L.J.; Batten, S. A critical comparison of two long-term zooplankton time series from the central-west North Sea. J. Plankton. Res. 2001, 23, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Colebrook, J.M. Continuous plankton records: Relationships between species of phytoplankton and zooplankton in the seasonal cycle. Mar. Biol. 1984, 83, 313–323. [Google Scholar] [CrossRef]
- Tavernini, S. Seasonal and inter-annual zooplankton dynamics in temporary pools with different hydroperiods. Limnologica 2008, 38, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Araujo, H.M.P.; Nascimento-Vieira, D.A.; Neumann-Leitão, S.; Schwamborn, R.; Lucas, A.P.; Alves, J.P. Zooplankton community dynamics in relation to the seasonal cycle and nutrient inputs in an urban tropical estuary in Brazil. Braz. J. Biol. 2008, 68, 751–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinle, D.R. Temperature and zooplankton. Chesap. Sci. 1969, 10, 186–209. [Google Scholar] [CrossRef]
- Pepin, P.; Colbourne, E.; Maillet, G. Seasonal patterns in zooplankton community structure on the Newfoundland and Labrador Shelf. Prog. Oceanogr. 2011, 91, 273–285. [Google Scholar] [CrossRef]
- David, V.; Sautour, B.; Chardy, P.; Leconte, M. Long-term changes of the zooplankton variability in a turbid environment: The Gironde estuary (France). Estuar. Coast. Shelf Sci. 2005, 64, 171–184. [Google Scholar] [CrossRef]
- Richardson, A.J. In hot water: Zooplankton and climate change. ICES J. Mar. Sci. 2008, 65, 279–295. [Google Scholar] [CrossRef] [Green Version]
- Angeler, D.G.; Moreno, J.M. Zooplankton community resilience after press-type anthropogenic stress in temporary ponds. Ecol. Appl. 2007, 17, 1105–1115. [Google Scholar] [CrossRef]
- Greve, W.; Reiners, F.; Nast, J.; Hoffmann, S. Helgoland Roads meso-and macrozooplankton time-series 1974 to 2004: Lessons from 30 years of single spot, high frequency sampling at the only off-shore island of the North Sea. Helgol. Mar. Res. 2004, 58, 274–288. [Google Scholar] [CrossRef] [Green Version]
- Möllmann, C.; Müller-Karulis, B.; Kornilovs, G.; St John, M.A. Effects of climate and overfishing on zooplankton dynamics and ecosystem structure: Regime shifts, trophic cascade, and feedback loops in a simple ecosystem. ICES J. Mar. Sci. 2008, 65, 302–310. [Google Scholar] [CrossRef] [Green Version]
- Kostelich, E.J.; Schreiber, T. Noise reduction in chaotic time-series data: A survey of common methods. Phys. Rev. E 1993, 48, 1752–1763. [Google Scholar] [CrossRef]
- Mackas, D.L.; Beaugrand, G. Comparisons of zooplankton time series. J. Mar. Syst. 2010, 79, 286–304. [Google Scholar] [CrossRef]
- Yoshida, T.; Urabe, J.; Elser, J.J. Assessment of “top down” and ‘‘bottom-up’’ forces as determinants of rotifer distribution among lakes in Ontario, Canada. Ecol. Res. 2003, 18, 639–650. [Google Scholar] [CrossRef]
- Arndt, H. Rotifers as predators on components of microbial web (bacteria, heterotrophic flagellates, ciliates). Hydrobiologia 1993, 255, 231–246. [Google Scholar] [CrossRef]
- Gilbert, J.J. Further observations on developmental polymorphism and its evolution in the rotifer Brchionus calyciflorus. Freshw. Biolog. 1980, 10, 281–294. [Google Scholar] [CrossRef]
- Devetter, M. Influence of environmental factors on the rotifer assemblage in an artificial lake. Hydrobiologia 1998, 387–388, 171–178. [Google Scholar] [CrossRef]
- Liang, D.; Wang, Q.; Wei, N.; Tang, C.; Sun, X.; Yang, Y. Biological indicators of ecological quality in typical urban river-lake ecosystems: The planktonic rotifer community and its response to environmental factors. Ecol. Indic. 2020, 112, 106–127. [Google Scholar] [CrossRef]
- Chang, K.H.; Doi, H.; Imai, H.; Gunji, F.; Nakano, S.A. Longitudinal changes in zooplankton distribution below a reservoir outfall with reference to river planktivory. Limnology 2008, 9, 125–133. [Google Scholar] [CrossRef]
- Thouvenot, A.; Debroas, D.; Richardot, M.; Bertin Jugnia, L.; Dévaux, J. A study of changes between years in the structure of plankton community in a newly-flooded reservoir. Arch. Für Hydrobiol. 2000, 149, 131–152. [Google Scholar] [CrossRef]
- Lee, J.M.; Yoon, S.M.; Lee, J.J.; Park, J.G.; Lee, J.H.; Chang, C.Y. Zooplankton Fauna and the Interrelationship Among Cladoceran Populations and Microcystis aeruginosa (Cyanophyceae) during the Cyanobacterial Blooming Season at Daecheong Lake, South Korea. Korean J. Ecol. Environ. 2005, 38, 146–159. [Google Scholar]
- Patterson, D.J.; Hedley, S. Free-Living Freshwater Protozoa; CRC Press: London, UK, 1996. [Google Scholar]
- Flössner, D. Die Haplopoda und Cladocera (ohne Bosminidae) Mitteleuropas; Backhuys Publishers: Leiden, The Netherlands, 2000; p. 428. [Google Scholar]
- Jeong, H.G.; Kotov, A.A.; Lee, W. Checklist of the freshwater Cladocera (Crustacea: Branchiopoda) of South Korea. Proc. Biol. Soc. Wash. 2014, 127, 216–228. [Google Scholar] [CrossRef]
- Chang, C.Y. Illustrated Encyclopedia of Fauna & Flora of Korea: Inland Water Copepoda; Jeonghaeng-sa: Seoul, Republic of Korea, 2009; Volume 42. [Google Scholar]
- Williamson, C.E. Invertebrate predation on planktonic rotifers. Hydrobiologia 1983, 104, 385–396. [Google Scholar] [CrossRef]
- Gilbert, J.J. Competition between rotifers and Daphnia. Ecology 1985, 66, 1943–1950. [Google Scholar] [CrossRef]
- Bonecker, C.C.; Aoyagui, A.S.M. Relationships between rotifers, phytoplankton and bacterioplankton in the Corumbá reservoir, Goiás State, Brazil. In Rotifera X; Springer: Dordrecht, The Netherlands, 2005; pp. 415–421. [Google Scholar]
- Zhang, M.; Shi, X.; Yang, Z.; Yu, Y.; Shi, L.; Qin, B. Long-term dynamics and drivers of phytoplankton biomass in eutrophic Lake Taihu. Sci. Total Environ. 2018, 645, 876–886. [Google Scholar] [CrossRef]
- Pirasteh, S.; Mollaee, S.; Fatholahi, S.N.; Li, J. Estimation of phytoplankton chlorophyll-a concentrations in the Western Basin of Lake Erie using Sentinel-2 and Sentinel-3 data. Can. J. Remote Sens. 2020, 46, 585–602. [Google Scholar] [CrossRef]
- Zhu, C.; Zhang, J.; Nawaz, M.Z.; Mahboob, S.; Al-Ghanim, K.A.; Khan, I.A.; Lu, Z.; Chen, T. Seasonal succession and spatial distribution of bacterial community structure in a eutrophic freshwater Lake, Lake Taihu. Sci. Total Environ. 2019, 669, 29–40. [Google Scholar] [CrossRef]
- Kendall, M.; Stuart, A. The Advanced Theory of Statistics; Griffin: London, UK, 1983; Volume 3, pp. 410–414. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 15 August 2022).
- Hill, M.O.; Gauch, H.G. Detrended Correspondence Analysis: An Improved Ordination Technique. Vegetatio 1980, 42, 47–58. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package Version 2013, 2, 321–326. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 August 2022).
- Zhao, C.; Liu, C.; Zhao, J.; Xia, J.; Yu, Q.; Eamus, D. Zooplankton in highly regulated rivers: Changing with water environment. Ecol. Eng. 2013, 58, 323–334. [Google Scholar] [CrossRef]
- Inaotombi, S.; Gupta, P.K.; Mahanta, P.C. Influence of abiotic factors on the spatio-temporal abundance of rotifers in a subtropical lake of western Himalaya. Water Air Soil Poll. 2016, 227, 1–15. [Google Scholar] [CrossRef]
- Nandini, S.; Sánchez-Zamora, C.; Sarma, S.S.S. Toxicity of cyanobacterial blooms from the reservoir Valle de Bravo (Mexico): A case study on the rotifer Brachionus calyciflorus. Sci. Total Environ. 2019, 688, 1348–1358. [Google Scholar] [CrossRef]
- Schlüter, M.; Groeneweg, J.; Soeder, C.J. Impact of rotifer grazing on population dynamics of green microalgae in high-rate ponds. Water Res. 1987, 21, 1293–1297. [Google Scholar] [CrossRef]
- Dodds, W.K.; Bouska, W.W.; Eitzmann, J.L.; Pilger, T.J.; Pitts, K.L.; Riley, A.J.; Schloesser, J.T.; Thornbrugh, D.J. Eutrophication of US freshwaters: Analysis of potential economic damages. Environ. Sci. Technol. 2009, 43, 12–19. [Google Scholar] [CrossRef] [Green Version]
- May, L. Rotifer occurrence in relation to water temperature in Loch Leven, Scotland. Hydrobiologia 1983, 104, 311–315. [Google Scholar] [CrossRef]
- Park, G.S.; Marshall, H.G. The trophic contributions of rotifers in tidal freshwater and estuarine habitats. Estuar. Coast. Shelf Sci. 2000, 51, 729–742. [Google Scholar] [CrossRef]
- Lehtovaara, A.; Arvola, L.; Keskitalo, J.; Olin, M.; Rask, M.; Salonen, K.; Sarvala, J.; Tulonen, T.; Vuorenmaa, J. Responses of zooplankton to long-term environmental changes in a small boreal lake. Boreal Environ. Res. 2014, 19, 97–111. [Google Scholar]
- Karabin, A. The pressure of pelagic predators of the genus Mesocyclops (Copepoda, Crustacea) on small zooplankton. Ekol. Pol. 1978, 26, 241–257. [Google Scholar]
- Stemberger, R.S.; Evans, M.S. Rotifer seasonal succession and copepod predation in Lake Michigan. J. Great Lakes Res. 1984, 10, 417–428. [Google Scholar] [CrossRef]
- Storch, H.V. Misuses of statistical analysis in climate research. In Analysis of Climate Variability; Springer: Berlin, Germany, 1999; pp. 11–26. [Google Scholar]
- Yule, G.U. Why do we sometimes get nonsense-correlations between time-series?—A study in sampling and the nature of time-series. J. R. Stat. Soc. 1926, 89, 1–63. [Google Scholar] [CrossRef]
- Findley, D.F.; Monsell, B.C.; Bell, W.R.; Otto, M.C.; Chen, B.C. New capabilities and methods of the X-12-ARIMA seasonal-adjustment program. J. Bus. Econ. Stat. 1998, 16, 127–152. [Google Scholar]
- Cleveland, R.B.; Cleveland, W.S.; McRae, J.E.; Terpenning, I. STL: A seasonal-trend decomposition. J. Off. Stat. 1990, 6, 3–73. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environment Factor | Value | Unit | Min | Max | Average | SD | CV (%) |
|---|---|---|---|---|---|---|---|
| Water Quality Factor | Temperature | °C | 6.53 | 27.7 | 19.36 | 6.23 | 32.15 |
| TN | mg/L | 0.66 | 2.473 | 1.37 | 0.33 | 23.82 | |
| TP | mg/L | 0.00 | 0.052 | 0.02 | 0.01 | 59.80 | |
| Chl-a | mg/m3 | 0.00 | 39.11 | 3.80 | 6.66 | 175.23 | |
| Biological Factor | Cladocerans | ind/L | 0.00 | 39.11 | 3.80 | 6.66 | 175.23 |
| Copepods | ind/L | 0.00 | 12.82 | 2.89 | 3.01 | 104.03 | |
| Nauplii | ind/L | 0.00 | 26.47 | 3.51 | 5.08 | 144.58 | |
| Protozoa | ind/L | 0.00 | 53.61 | 5.64 | 10.76 | 190.78 | |
| Blue-green algae | cells/mL | 0.00 | 345,538 | 12,409.88 | 49,921.40 | 402.27 | |
| Diatom | cells/mL | 8.00 | 3472 | 596.63 | 743.86 | 124.68 | |
| Green algae | cells/mL | 4.00 | 1190 | 223.53 | 270.94 | 121.21 |
| Abundance Rank | Rotifer Species | Total Individuals (ind/L) | Species Ratio (%) | Frequency | Rotifer Group |
|---|---|---|---|---|---|
| 1 | Polyarthra vulgaris | 489.33 | 35.127 | 47 | HAF Group |
| 2 | Keratella cochlearis | 342.37 | 24.577 | 48 | HAF Group |
| 3 | Synchaeta oblonga | 155.06 | 11.131 | 40 | HAF Group |
| 4 | Hexarthra (Pedalla) mira | 99.62 | 7.151 | 17 | HAF Group |
| 5 | Keratella valga | 60.62 | 4.352 | 12 | * |
| 6 | Ploesoma truncatum | 49.34 | 3.542 | 32 | HAF Group |
| 7 | Trichocerca sp. | 38.13 | 2.737 | 29 | HAF Group |
| 8 | Pompholyx complanata | 30.53 | 2.192 | 19 | HAF Group |
| 9 | Conochilus sp. | 24.17 | 1.735 | 19 | HAF Group |
| 10 | Kellicottia bostoniensis | 24 | 1.723 | 19 | HAF Group |
| 11 | Large Trichocerca spp. (elongata, cylindrica) | 16.25 | 1.167 | 17 | MAF Group |
| 12 | Ascomorpha ecaudis | 12.69 | 0.911 | 20 | * |
| 13 | Filinia longiseta | 9.55 | 0.686 | 12 | MAF Group |
| 14 | Monostyla closterocerca | 9.45 | 0.678 | 8 | MAF Group |
| 15 | Filinia terminalis | 5.05 | 0.363 | 5 | MAF Group |
| 16 | Euchlanis dilatata | 4.59 | 0.329 | 12 | MAF Group |
| 17 | Brachionus angularis | 4.41 | 0.317 | 5 | MAF Group |
| 18 | Polyarthra euryptera | 3.73 | 0.268 | 5 | MAF Group |
| 19 | Asplanchna sp. | 3.5 | 0.251 | 11 | MAF Group |
| 20 | Lecane flexilis | 2.45 | 0.176 | 5 | MAF Group |
| 21 | Kellicottia longispina | 1.98 | 0.142 | 3 | MAF Group |
| 22 | Brachionus quadridentatus | 1.76 | 0.126 | 3 | MAF Group |
| 23 | Keratella quadrata | 0.94 | 0.067 | 2 | LAF Group |
| 24 | Mytilina sp. | 0.63 | 0.045 | 2 | LAF Group |
| 25 | Brachionus forficula | 0.5 | 0.036 | 3 | LAF Group |
| 26 | Asplanchna sieboldi | 0.39 | 0.028 | 2 | LAF Group |
| 27 | Brachionus sp. | 0.36 | 0.026 | 2 | LAF Group |
| 28 | Monostyla bulla | 0.3 | 0.022 | 2 | LAF Group |
| 29 | Brachionus rubens | 0.28 | 0.020 | 1 | LAF Group |
| 30 | Brachionus calyciflorus | 0.26 | 0.019 | 1 | LAF Group |
| 31 | Lecane sp. | 0.26 | 0.019 | 3 | LAF Group |
| 32 | Monostyla sp. | 0.26 | 0.019 | 1 | LAF Group |
| 33 | Trichotria tetractis | 0.16 | 0.011 | 1 | LAF Group |
| 34 | Lecane luna | 0.08 | 0.006 | 1 | LAF Group |
| 35 | Notholca labis | 0.04 | 0.003 | 1 | LAF Group |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, G.-H.; Chang, K.-H.; Oh, H.-J.; Choi, Y.; Han, S.; Jeong, H.-G. Multivariate Analysis of Rotifer Community and Environmental Factors Using the Decomposed Components Extracted from a Time Series. Water 2022, 14, 4113. https://doi.org/10.3390/w14244113
Hong G-H, Chang K-H, Oh H-J, Choi Y, Han S, Jeong H-G. Multivariate Analysis of Rotifer Community and Environmental Factors Using the Decomposed Components Extracted from a Time Series. Water. 2022; 14(24):4113. https://doi.org/10.3390/w14244113
Chicago/Turabian StyleHong, Geun-Hyeok, Kwang-Hyeon Chang, Hye-Ji Oh, Yerim Choi, Sarang Han, and Hyun-Gi Jeong. 2022. "Multivariate Analysis of Rotifer Community and Environmental Factors Using the Decomposed Components Extracted from a Time Series" Water 14, no. 24: 4113. https://doi.org/10.3390/w14244113
APA StyleHong, G. -H., Chang, K. -H., Oh, H. -J., Choi, Y., Han, S., & Jeong, H. -G. (2022). Multivariate Analysis of Rotifer Community and Environmental Factors Using the Decomposed Components Extracted from a Time Series. Water, 14(24), 4113. https://doi.org/10.3390/w14244113