
Simple Is the Best: An Alternative Method for the Analysis of Free-Living Nematode Assemblage Structure



Abstract
:1. Introduction
2. Materials and Methods
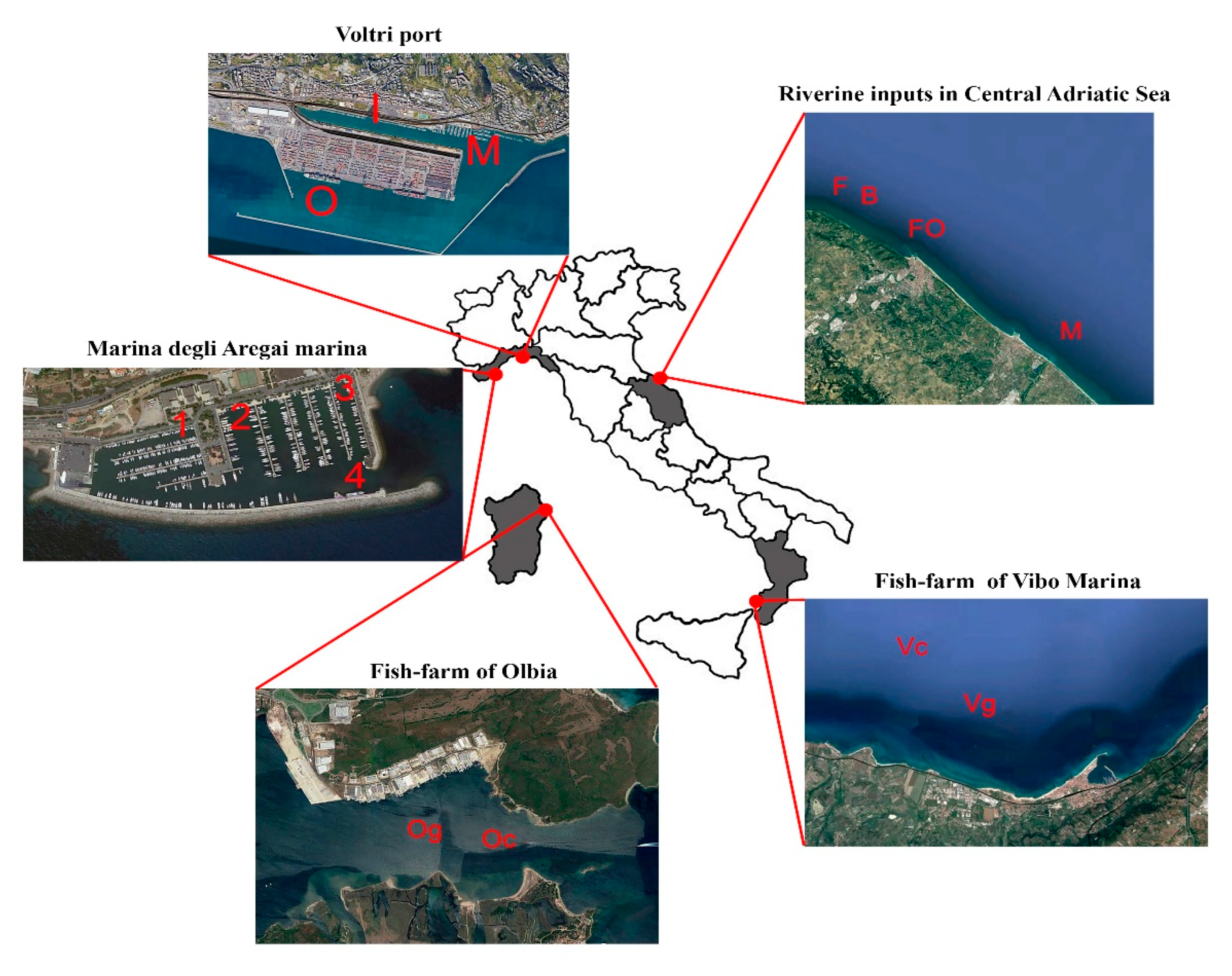
2.1. Study Areas
2.1.1. Port-Contaminated Sediments from Ligurian Sea (NW Mediterranean)
2.1.2. Coastal Sediments Influenced by Riverine Inputs in Central Adriatic Sea
2.1.3. Fish Farm Impact in the Tyrrhenian Sea
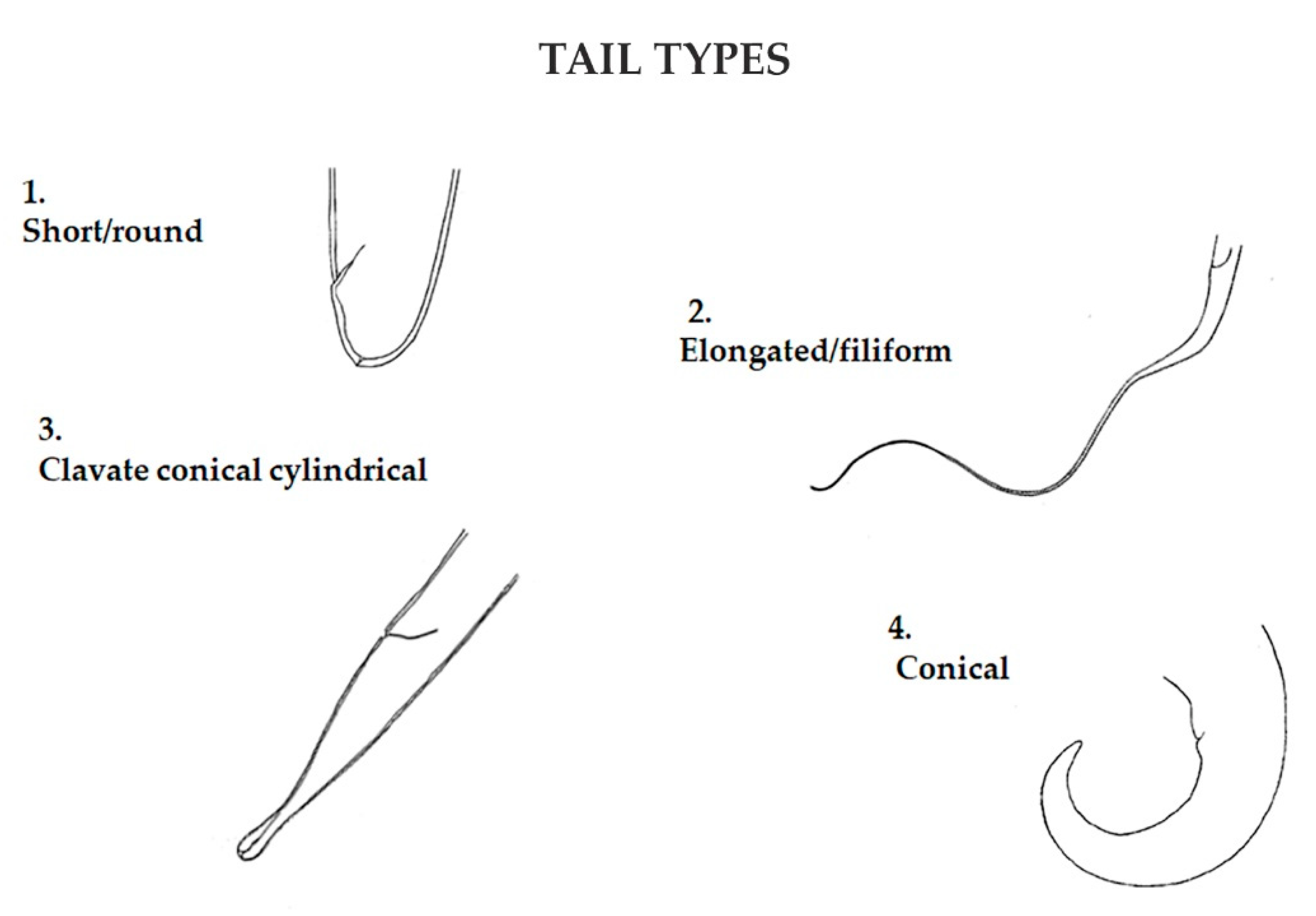
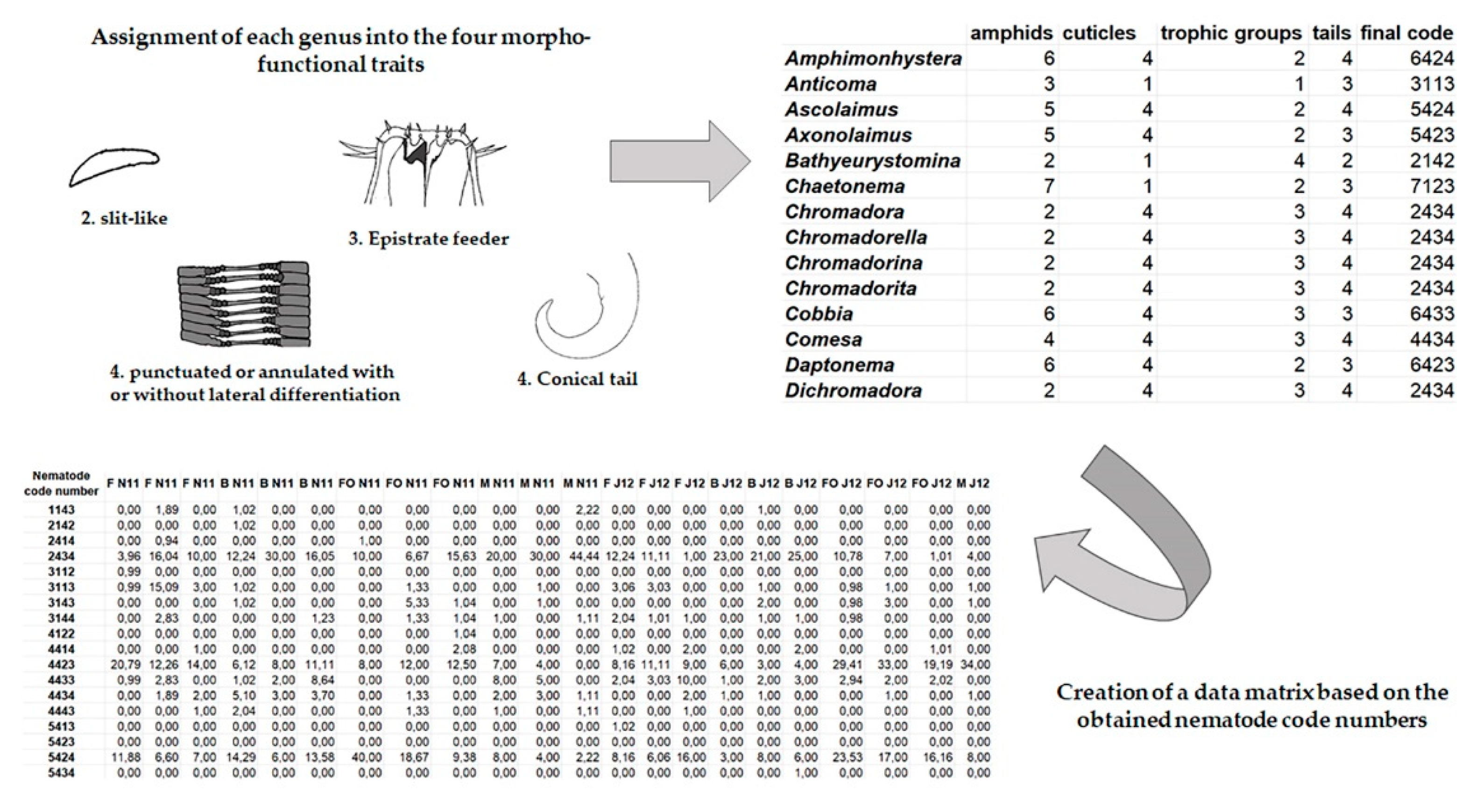
2.2. Nematodes
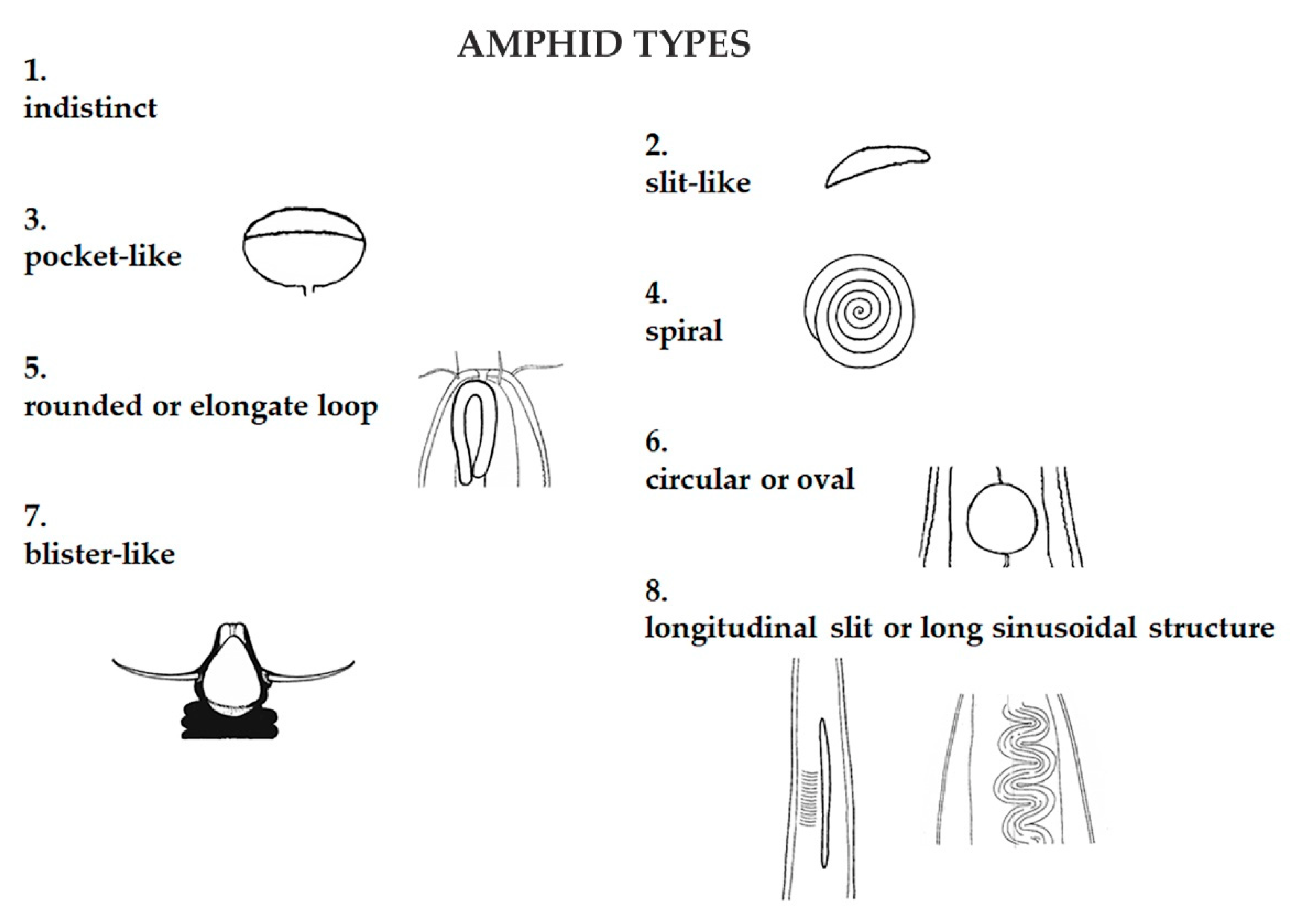
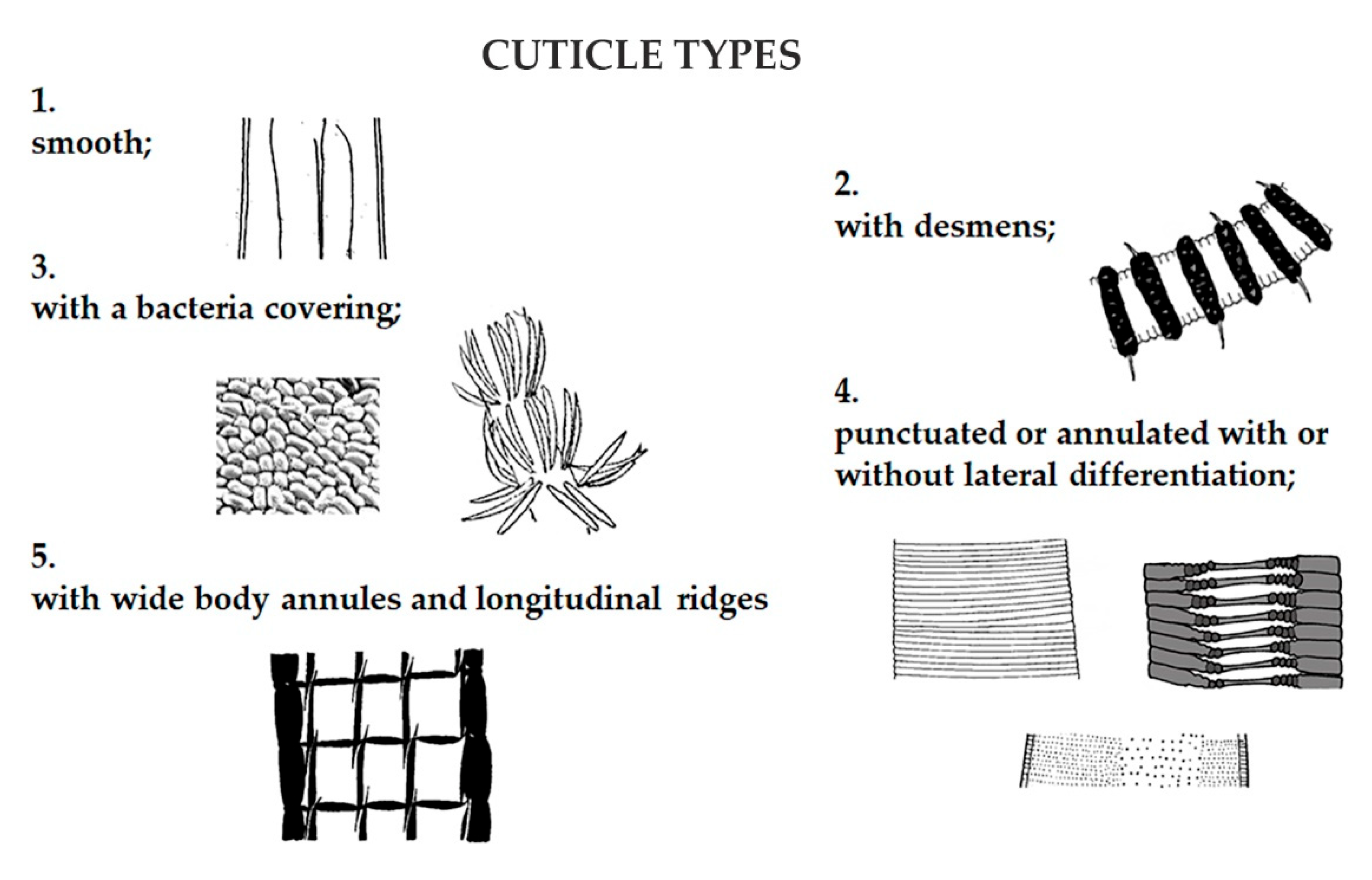
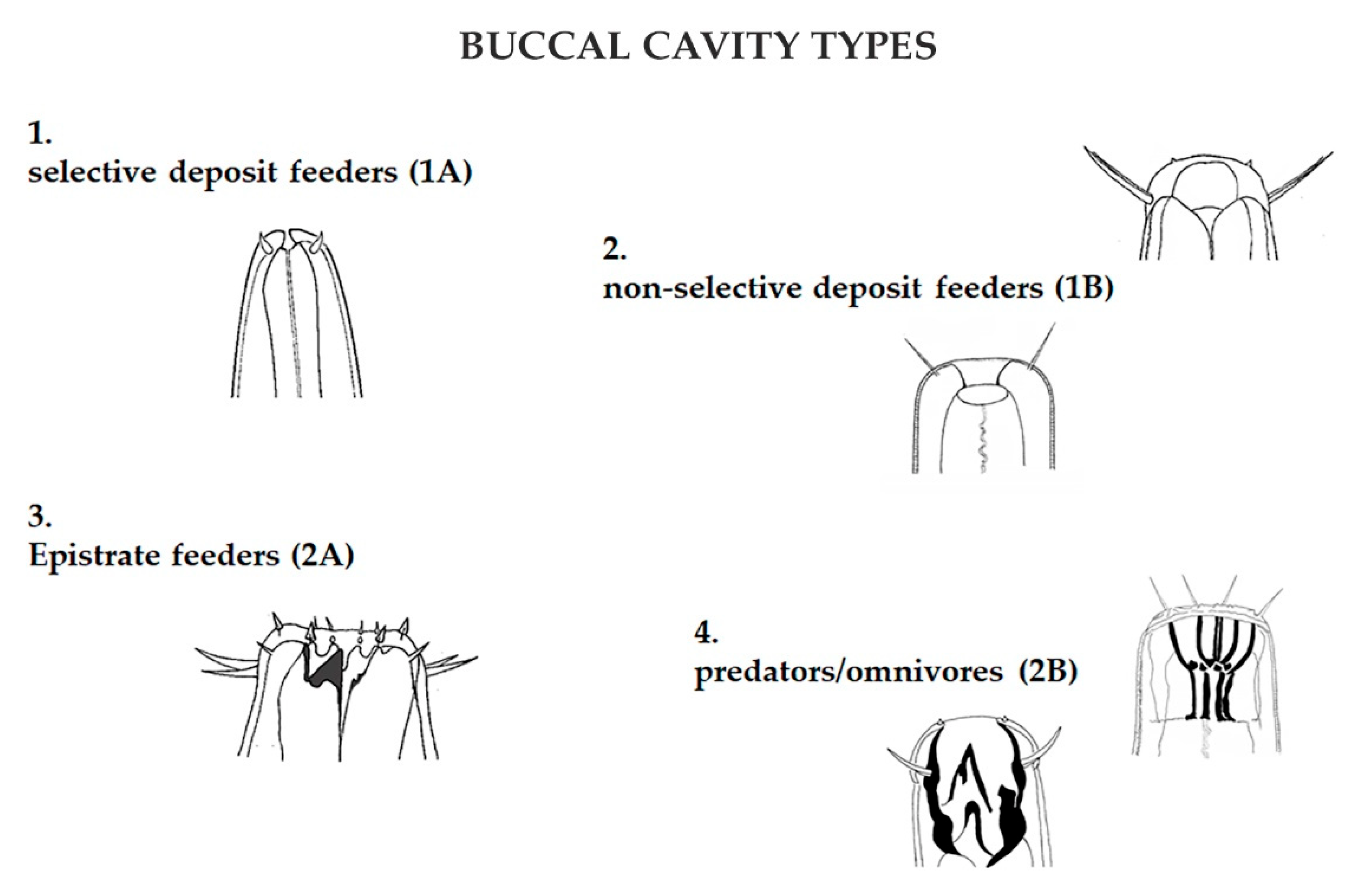
2.3. Data Analysis
3. Results and Discussion
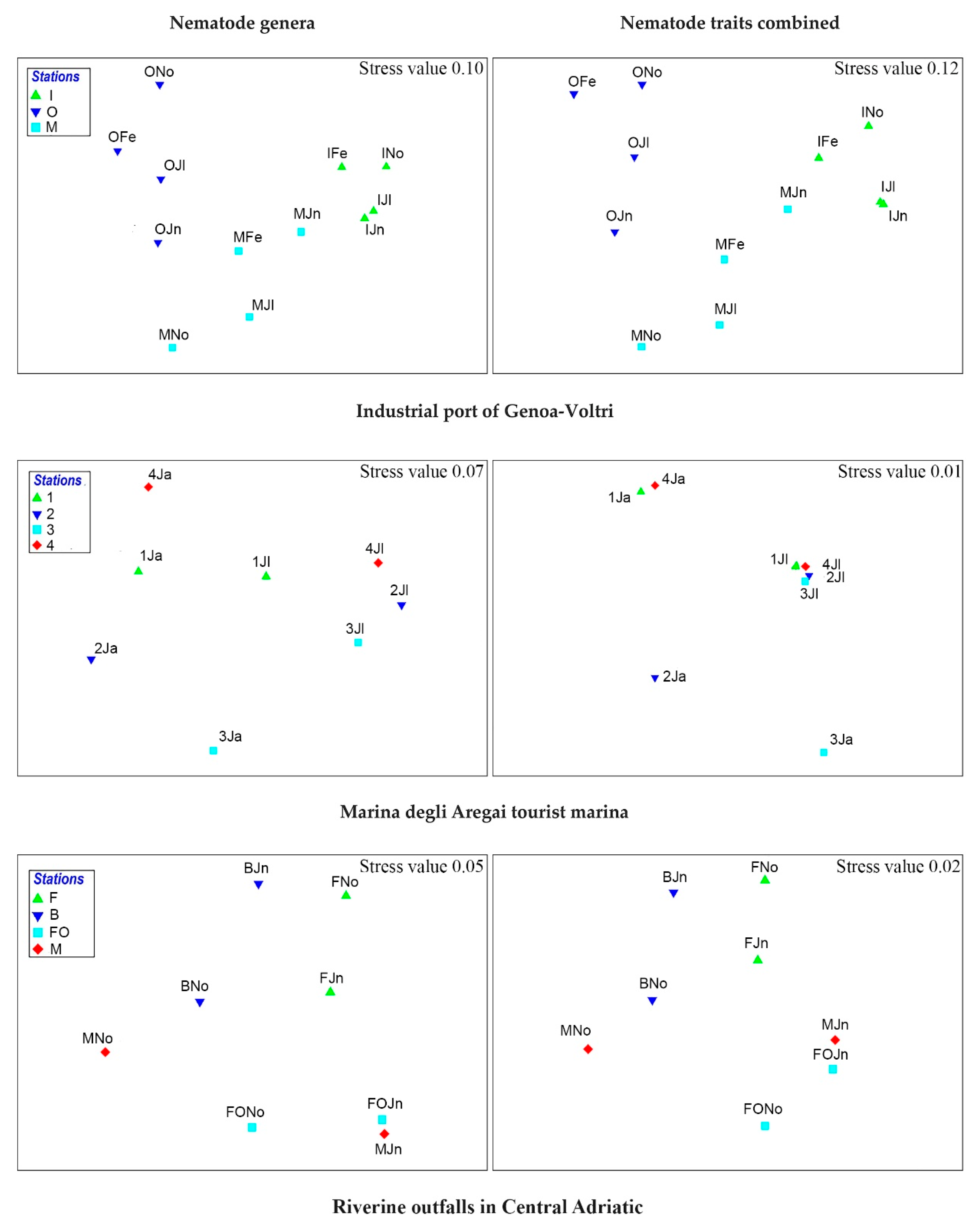
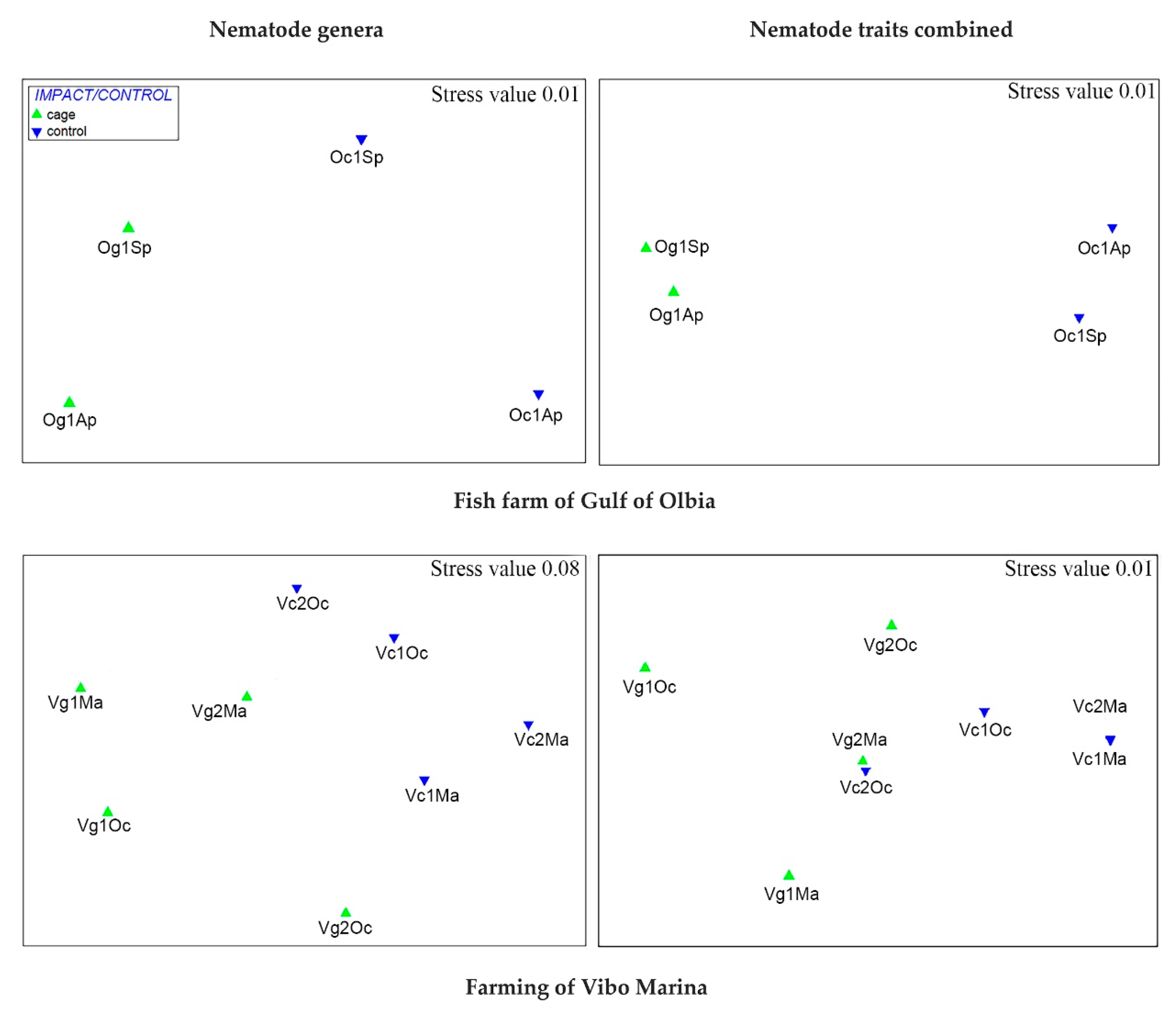
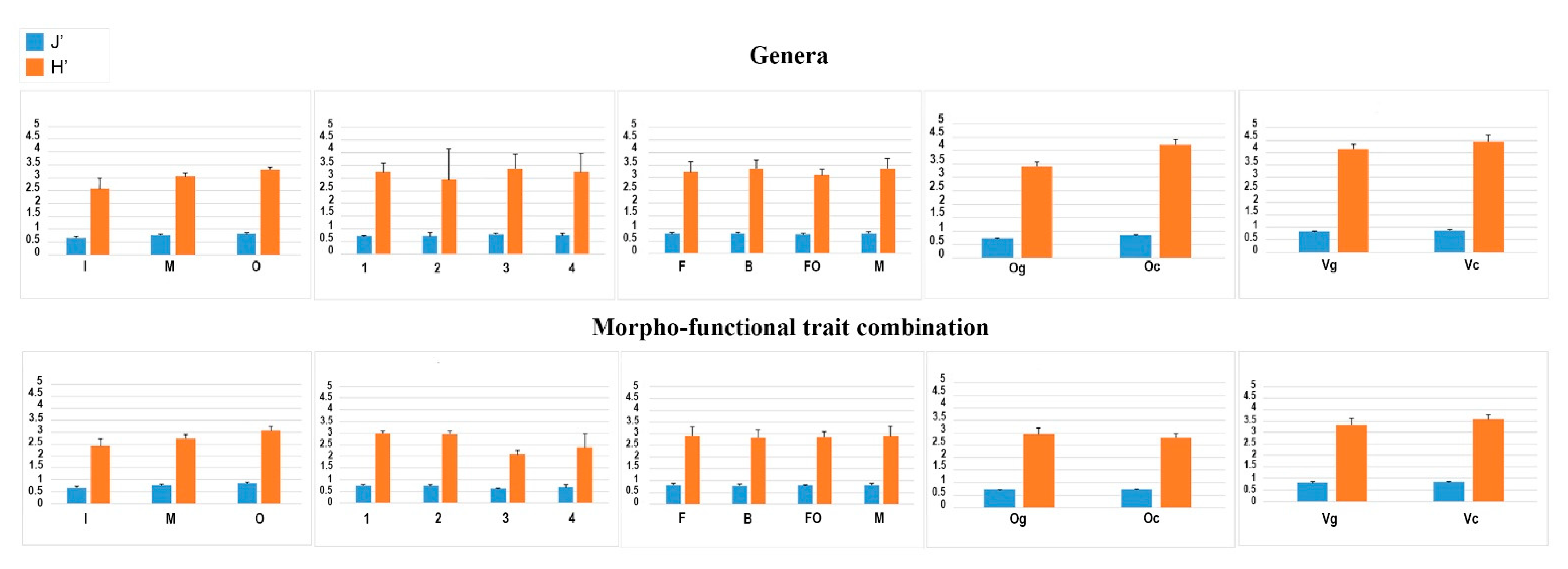
3.1. Does the Combination of Morpho-Functional Traits Reflect the Taxonomic Composition of Nematode Fauna?
3.2. Does the Combination of Morpho-Functional Traits Detect the Effect of a Different Human Pressure?
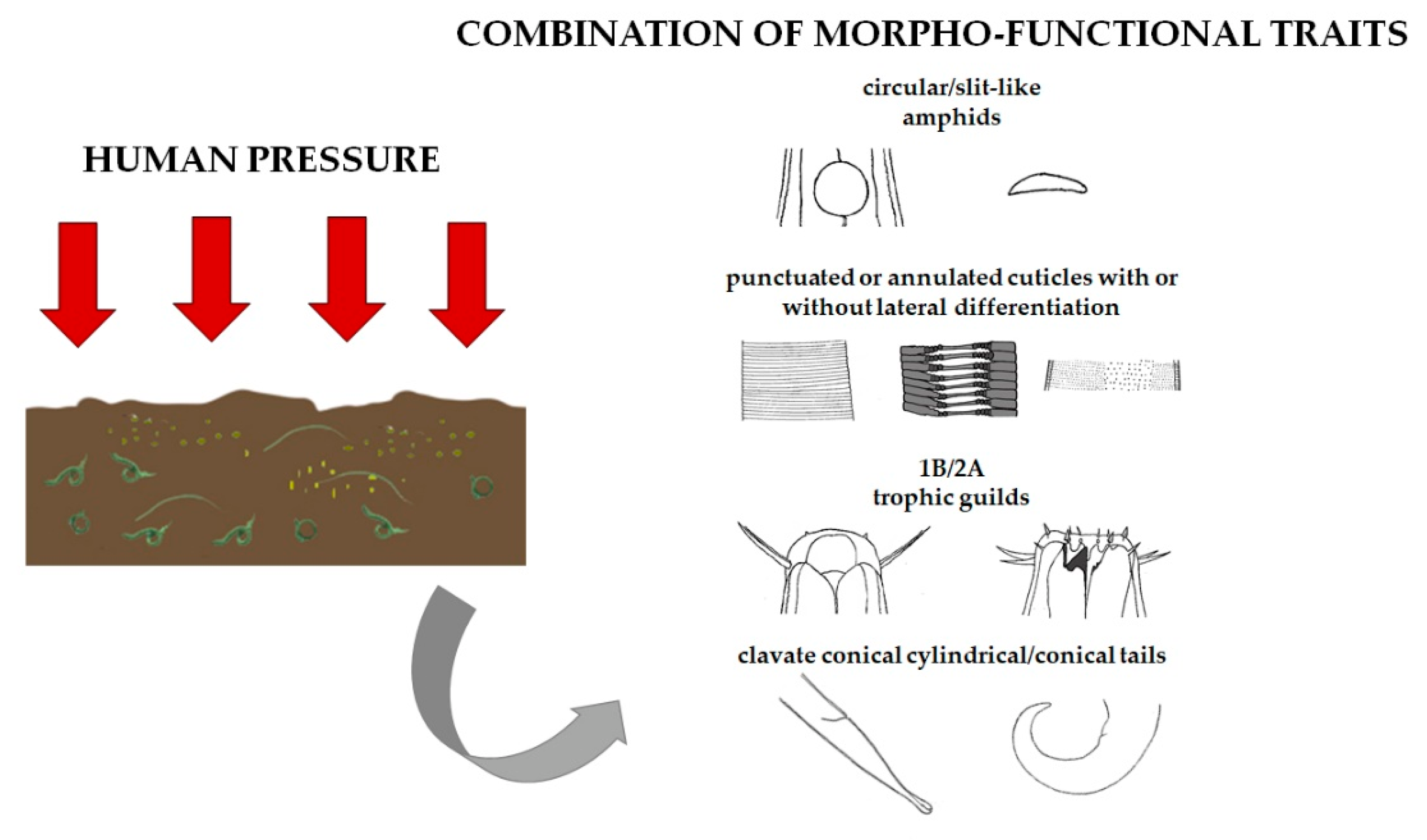
3.3. Are There Recurrent Combinations of Morpho-Functional Traits in Nematode Communities in Polluted Sediments?
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Matsuyama, A.; Yano, S.; Taninaka, T.; Kindaichi, M.; Sonoda, I.; Tada, A.; Akagi, H. Chemical characteristics of dissolved mercury in the pore water of Minamata Bay sediments. Mar. Pollut. Bull. 2018, 129, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Losi, V.; Grassi, E.; Balsamo, M.; Rocchi, M.; Gaozza, L.; Semprucci, F. Changes in taxonomic structure and functional traits of nematodes as tools in the assessment of port impact. Estuar. Coast. Shelf Sci. 2021, 260, 107524. [Google Scholar] [CrossRef]
- Pawlowski, J.; Bruce, K.; Panksep, K.; Aguirre, F.I.; Amalfitano, S.; Apothéloz-Perret-Gentil, L.; Baussant, T.; Bouchez, A.; Carugati, L.; Cermakova, K.; et al. Environmental DNA metabarcoding for benthic monitoring: A review of sediment sampling and DNA extraction methods. Sci. Total Environ. 2021, 19, 151783. [Google Scholar] [CrossRef]
- Armynot du Châtelet, E.; Bout-Roumazeilles, V.; Coccioni, R.; Frontalini, F.; Francescangeli, F.; Margaritelli, G.; Rettori, R.; Spagnoli, F.; Semprucci, F.; Trentesaux, A.; et al. Environmental control on a land-sea transitional setting–Integrated microfaunal, sedimentological, and geochemical approaches. Environ. Earth Sci. 2016, 75, 123. [Google Scholar] [CrossRef]
- Gambi, C.; Dell’Anno, A.; Corinaldesi, C.; Lo Martire, M.; Musco, L.; Da Ros, Z.; Armiento, G.; Danovaro, R. Impact of historical contamination on meiofaunal assemblages: The case study of Bagnoli-Coroglio Bay (southern Tyrrhenian Sea). Mar. Environ. Res. 2020, 156, 104907. [Google Scholar] [CrossRef] [PubMed]
- Schratzberger, M.; Somerfield, P.J. Effects of widespread human disturbances in the marine environment suggest a new agenda for meiofauna research is needed. Sci. Total Environ. 2020, 728, 138435. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Semprucci, F.; Vezzulli, L.; Balsamo, M.; Fabiano, M.; Albertelli, G. The use of nematodes in assessing ecological quality status in the Mediterranean coastal ecosystems. Ecol. Indic. 2011, 11, 328–336. [Google Scholar] [CrossRef]
- Hong, J.H.; Semprucci, F.; Raehyuk, J.; Kim, K.; Lee, S.; Jeon, D.; Yoo, H.; Kim, J.; Kim, J.; Yeom, J.; et al. Meiobenthic nematodes in the assessment of the relative impact of human activities on coastal marine ecosystem. Environ. Monit. Assess. 2020, 192, 81. [Google Scholar] [CrossRef]
- Sahraeian, N.; Sahafi, H.H.; Mosallanejad, H.; Ingels, J.; Semprucci, F. Temporal and spatial variability of free-living nematodes in a beach system characterized by domestic and industrial impacts (Bandar Abbas, Persian Gulf, Iran). Ecol. Indic. 2020, 118, 106697. [Google Scholar] [CrossRef]
- Baldrighi, E.; Semprucci, F.; Franzo, A.; Cvitkovic, I.; Bogner, D.; Despalatovic, M.; Berto, D.; Malgorzata Formalewicz, M.; Scarpato, A.; Frapiccini, E.; et al. Meiofaunal communities in four Adriatic ports: Baseline data for risk assessment in ballast water management. Mar. Pollut. Bull. 2019, 147, 171–184. [Google Scholar] [CrossRef]
- Nelson, W.G. A quantitative assessment of organic carbon content as a regional sediment-condition indicator. Ecol. Indic. 2020, 114, 106318. [Google Scholar] [CrossRef] [PubMed]
- Kiddon, J.; Sullivan, H.; Nelson, W.; Pelletier, M.; Harwell, L.; Nord, M.; Paulsen, S. Lessons learned from 30 years of assessing U.S. coastal water. In Water Quality–Science, Assessments and Policy; Summers, K., Ed.; InTechOpen: Rijeka, Croatia, 2020; pp. 1–22. [Google Scholar]
- Guerra-García, J.M.; Navarro-Barranco, C.; Ros, M.; Sedano, F.; Espinar, R.; Fernández-Romero, A.; Martínez-Laiz, G.; Cuesta, J.A.; Giráldez, I.; Morales, E.; et al. Ecological quality assessement of marinas: An integrative approach combining, biological and environmental data. J. Environ. Manag. 2021, 15, 286–112237. [Google Scholar] [CrossRef]
- Diaz, R.J.; Solan, M.; Valente, R.M. A review of approaches for classifying benthic habitats and evaluating habitat quality. J. Environ. Manag. 2004, 73, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Borja, A.; Barbone, E.; Basset, A.; Borgersen, G.; Brkljacic, M.; Elliott, M.; Garmendia, J.M.; Marques, J.C.; Mazik, K.; Muxika, I.; et al. Response of single benthic metrics and multi-metric methods to anthropogenic pressure gradients, in five distinct European coastal and transitional ecosystems. Mar. Pollut. Bull. 2011, 62, 499–513. [Google Scholar] [CrossRef]
- Vanaverbeke, J.; Merckx, B.; Degraer, S.; Vincx, M. Sediment-related distribution patterns of nematodes and macrofauna: Two sides of the benthic coin? Mar. Environ. Res. 2011, 71, 31–40. [Google Scholar] [CrossRef]
- Appeltans, W.; Ahyong, S.T.; Anderson, G.; Angel, M.V.; Artois, T.; Bailly, N.; Costello, M.J. The Magnitude of Global Marine Species Diversity. Curr. Biol. 2012, 22, 2189–2202. [Google Scholar] [CrossRef] [Green Version]
- Jouili, S.; Essid, N.; Semprucci, F.; Boufahja, F.; Nasri, A.; Beyrem, H. Environmental quality assessment of El Bibane lagoon (Tunisia) using taxonomical and functional diversity of meiofauna and nematodes. J. Mar. Biol. Assoc. UK 2017, 97, 1593–1603. [Google Scholar] [CrossRef]
- Bezerra, T.N.; Decraemer, W.; Eisendle-Flöckner, U.; Hodda, M.; Holovachov, O.; Leduc, D.; Miljutin, D.; Mokievsky, V.; Peña Santiago, R.; Sharma, J.; et al. Nemys: World Database of Nematodes. Available online: http://nemys.ugent.be/ (accessed on 19 December 2021).
- Coull, B.C.; Chandler, G.T. Pollution and meiofauna: Field, laboratory and mesocosm studies. Oceanogr. Mar. Biol. 1992, 30, 191–271. [Google Scholar]
- Giere, O. Meiobenthology: The Microscopic Motile Fauna of Aquatic Sediments, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2009; p. 527. [Google Scholar]
- Schmid-Araya, J.M.; Schmid, P.E. Trophic relationships: Integrating meiofauna into a realistic benthic food web. Freshw. Biol. 2000, 44, 149–163. [Google Scholar] [CrossRef]
- Balsamo, M.; Semprucci, F.; Frontalini, F.; Coccioni, R. Meiofauna as a tool for marine ecosystem biomonitoring. Mar. Ecosyst. 2012, 4, 77–104. [Google Scholar]
- Chen, C.A.; Soo, C.L.; Balsamo, M.; Semprucci, F. An approach based on nematode descriptors for the ecological quality (EcoQ) classification of the Malaysian coasts. Mar. Biodivers. 2018, 48, 117–126. [Google Scholar] [CrossRef]
- Semprucci, F.; Balsamo, M.; Appolloni, L.; Sandulli, R. Assessment of ecological quality status along the Apulian coasts (Eastern Mediterranean Sea) based on meiobenthic and nematode assemblages. Mar. Biodivers. 2018, 48, 105–115. [Google Scholar] [CrossRef]
- Boufahja, F.; Semprucci, F.; Beyrem, H.; Bhadury, P. Marine nematodes taxonomy in Africa: Promising prospects against scarcity of information. J. Nematic. 2015, 47, 198–206. [Google Scholar]
- Derycke, S.; Maes, S.; Van den Bulcke, L.; Vanhollebeke, J.; Wittoeck, J.; Hillewaert, H.; Ampe, B.; Haegeman, A.; Hostens, K.; De Backer, A. Optimisation of metabarcoding for monitoring marine macrobenthos: Primer choice and morphological traits determine species detection in bulkDNA and eDNA from the ethanol preservative. Front. Mar. Sci. 2021, 8, 675. [Google Scholar]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Sechi, V.; De Goede, R.G.M.; Rutgers, M.; Brussaard, L.; Mulder, C. Functional diversity in nematode communities across terrestrial ecosystems. Basic Appl. Ecol. 2018, 30, 76–86. [Google Scholar] [CrossRef]
- Chalcraft, D.R.; Resetarits, W.J. Mapping functional similarity of predators on the basis of trait similarities. Am. Nat. 2003, 162, 390–402. [Google Scholar] [CrossRef] [Green Version]
- Schratzberger, M.; Warr, K.; Rogers, S.I. Functional diversity of nematode community in the south-western North Sea. Mar. Environ. Res. 2007, 63, 368–389. [Google Scholar] [CrossRef] [Green Version]
- Wieser, W. Die beziehung zwischen mundhöhlengestalt, ernährungsweise und vorkommen bei freilebenden marinen nematoden. Eine ökologisch-morphologische studie. Ark. Zool. 1953, 4, 439–484. [Google Scholar]
- Wieser, W. Free-living Marine Nematodes IV. General Part. Acta Univ. Lund. 1959, 55, 1–111. [Google Scholar]
- Thistle, D.; Sherman, K.M. The nematode fauna of a deep-sea site exposed to strong near-bottom currents. Deep Sea Res. 1985, 32, 1077–1088. [Google Scholar] [CrossRef]
- Thistle, D.; Lambshead, P.J.D.; Sherman, K.M. Nematode tail-shape groups respond to environmental differences in the deep sea. Life Environ. 1995, 45, 107–115. [Google Scholar]
- Vanaverbeke, J.; Steyaert, M.; Vanreusel, A.; Vincx, M. Nematode biomass spectra as descriptors of functional changes due to human and natural impact. Mar. Ecol. Prog. Ser. 2003, 249, 157–170. [Google Scholar] [CrossRef]
- Losi, V.; Moreno, M.; Gaozza, L.; Vezzulli, L.; Fabiano, M.; Albertelli, G. Nematode biomass and allometric attributes as indicators of environmental quality in a Mediterranean harbour (Ligurian Sea, Italy). Ecol. Indic. 2013, 30, 80–89. [Google Scholar] [CrossRef]
- Franzo, A.; Del Negro, P. Functional diversity of free-living nematodes in river lagoons: Can biological traits analysis (BTA) integrate traditional taxonomic-based approaches as a monitoring tool? Mar. Environ. Res. 2019, 145, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Thiele, T.R.; Faumont, S.; Lockery, S.R. The neural network for chemotaxis to tastants in Caenorhabditis elegans is specialized for temporal differentiation. J. Neurosci. 2009, 29, 11904–11911. [Google Scholar] [CrossRef] [Green Version]
- Zullini, A.; Semprucci, F. Morphological differences between free-living soil and freshwater nematodes in relation to their environments. Nematology 2020, 22, 125–132. [Google Scholar] [CrossRef]
- Raes, M.; De Troch, M.; Ndaro, S.G.M.; Muthumbi, A.; Guilini, K.; Vanreusel, A. The structuring role of microhabitat type in coral degradation zones: A case study with marine nematodes from Kenya and Zanzibar. Coral Reefs. 2007, 26, 113–126. [Google Scholar] [CrossRef]
- Semprucci, F.; Cesaroni, L.; Guidi, L.; Balsamo, M. Do the morphological and functional traits of free-living marine nematodes mirror taxonomical diversity? Mar. Environ. Res. 2018, 135, 114–122. [Google Scholar] [CrossRef]
- Ichiishi, K.; Ekino, T.; Kanzaki, N.; Shinya, R. Thick cuticles as an anti-predator defense in nematodes. Nematology 2021, 24, 11–20. [Google Scholar] [CrossRef]
- Moreno, M.; Ferrero, T.J.; Gallizia, I.; Vezzulli, L.; Albertelli, G.; Fabiano, M. An assessment of the spatial heterogeneity of environmental disturbance within an enclosed harbour through the analysis of meiofauna and nematode assemblages. Estuar. Coast. Shelf. Sci. 2008, 77, 565–576. [Google Scholar] [CrossRef]
- Moreno, M.; Albertelli, G.; Fabiano, M. Nematode response to metal, PAHs and organic enrichment in tourist marinas of the Mediterranean Sea. Mar. Poll. Bull. 2009, 58, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Semprucci, F.; Boi, P.; Manti, A.; Covazzi Harriague, A.; Rocchi, M.; Colantoni, P.; Papa, S.; Balsamo, M. Benthic communities along a littoral of the Central Adriatic Sea (Italy). Helgol. Mar. Res. 2010, 64, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Semprucci, F.; Sbrocca, C.; Rocchi, M.; Balsamo, M. Temporal changes of the meiofaunal assemblage as a tool for the assessment of the ecological quality status. J. Mar. Biolog. Assoc. UK 2015, 95, 247–254. [Google Scholar] [CrossRef]
- Semprucci, F.; Frontalini, F.; Sbrocca, C.; Armynot du Châtelet, E.; Bout-Roumazeilles, V.; Coccioni, R.; Balsamo, M. Meiobenthos and free-living nematodes as tools for biomonitoring environments affected by riverine impact. Environ. Monit. Assess. 2015, 187, 251. [Google Scholar] [CrossRef] [PubMed]
- Vezzulli, L.; Moreno, M.; Marin, V.; Pezzati, E.; Bartoli, M.; Fabiano, M. Organic waste impact of capture- based Atlantic bluefin tuna aquaculture at an exposed site in the Mediterranean Sea. Estuar. Coast. Shelf. Sci. 2008, 78, 369–384. [Google Scholar] [CrossRef]
- Danovaro, R.; Gambi, C.; Mirto, S.; Sandulli, R.; Ceccherelli, V.M. Mediterranean Marine Benthos: A Manual of Methods for Its Sampling and Study; Gambi, M.C., Dappiano, M., Eds.; S.I.B.M-Società Italiana di Biologia Marina: Genova, Italy, 2004; pp. 55–97. [Google Scholar]
- Seinhorst, J.W. A rapid method for the transfer of nematodes from fixative to unhydrous glycerine. Nematologica 1959, 4, 67–69. [Google Scholar] [CrossRef] [Green Version]
- Platt, H.M.; Warwick, R.M. Free-living Marine Nematodes. Part, I. British Enoplids. In Synopses of the British Fauna (New Series); Cambridge University Press: Cambridge, UK, 1983; Volume 28, p. 307. [Google Scholar]
- Platt, H.M.; Warwick, R.M. Free-living Marine Nematodes. Part II. British Chromadorids. In Synopses of the British Fauna (New Series); Brill: Leiden, The Netherlands, 1988; Volume 38, p. 502. [Google Scholar]
- Warwick, R.M.; Platt, H.M.; Somerfield, P.J. Free-living Marine Nematodes. Part III. British Monhysterids. In Synopses of the British Fauna (New Series); Field Studies Council: Shrewsbury, UK, 1998; Volume 53, p. 296. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, MA, USA, 2006; p. 192. [Google Scholar]
- Semprucci, F.; Losi, V.; Moreno, M. A review of Italian research on free-living marine nematodes and the future perspectives on their use as Ecological Indicators (EcoInds). Mediterr. Mar. Sci. 2015, 16, 352–365. [Google Scholar] [CrossRef] [Green Version]
- Armenteros, M.; Pérez-García, J.A.; Marzo-Pérez, D.; Rodríguez-García, P. The Influential Role of the Habitat on the Diversity Patterns of Free-Living Aquatic Nematode Assemblages in the Cuban Archipelago. Diversity 2019, 11, 166. [Google Scholar] [CrossRef] [Green Version]
- Inglis, W.G. The structure of the nematode cuticle. Proc. Zool. Soc. Lond. 1964, 143, 465–502. [Google Scholar] [CrossRef]
- Mirto, S.; La Rosa, T.; Gambi, C.; Danovaro, R.; Mazzola, A. Nematode community response to fish-farm impact in the western Mediterranean. Environ. Pollut. 2002, 116, 203–214. [Google Scholar] [CrossRef]
- Schratzberger, M.; Wall, C.M.; Reynolds, W.J.; Reed, J.; Waldock, M.J. Effects of paint-derived tributyltin on structure of estuarine nematode assemblages in experimental microcosms. J. Exp. Mar. Biol. Ecol. 2002, 272, 217–235. [Google Scholar] [CrossRef]
- Boufahja, F.; Semprucci, F. Stress-induced selection of a single species from an entire meiobenthic nematode assemblage: Is this possible using iron enrichment and does pre-exposure affect the ease of the process? Environ. Sci. Pollut. Res. 2015, 22, 1979–1998. [Google Scholar] [CrossRef]
- Cibic, T.; Franzo, A.; Nasi, F.; Auriemma, R.; Del Negro, P. The port of Trieste (northern Adriatic Sea)—A case study of the “ecosystem approach to management”. Front. Mar. Sci. 2017, 4, 336. [Google Scholar] [CrossRef] [Green Version]
- Bellec, L.; Cambon-Bonavita, M.-A.; Durand, L.; Aube, J.; Gayet, N.; Sandulli, R.; Brandily, C.; Zeppilli, D. Microbial communities of the shallow-water hydrothermal vent near Naples, Italy, and chemosynthetic symbionts associated With a free-living marine nematode. Front. Microbiol. 2020, 11, 2023. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, S.; Prein, M.; Valentin, C. Mass aggregations of the free-living marine nematode Pontonema vulgare (Oncholaimidae) in organically polluted fjords. Mar. Ecol. Prog. Ser. 1987, 37, 27–34. [Google Scholar] [CrossRef]
- Thiermann, F.; Vismann, B.; Giere, O. Sulphide tolerance of the marine nematode Oncholaimus campylocercoides-a result of internal sulphur formation? Mar. Ecol. Prog. Ser. 2000, 193, 251–259. [Google Scholar] [CrossRef]
- Bargmann, C.I. Chemosensation in C. elegans; WormBook, 2006; pp. 1–29. Available online: https://www.ncbi.nlm.nih.gov/books/NBK19746/ (accessed on 13 February 2022).
- Guden, R.M.; Derycke, S.; Moens, T. A Multi-Faceted Approach to Understand How Resource Diversity Can Mediate the Coexistence of Cryptic Marine Nematode Species. Front. Mar. Sci. 2021, 8, 777425. [Google Scholar] [CrossRef]
- Wakkaf, T.; Allouche, M.; Harrath, A.H.; Mansour, L.; Alwasel, S.; Mohamed Thameemul Ansari, K.G.; Beyrem, H.; Sellami, B.; Boufahja, F. The individual and combined effects of cadmium, polyvinyl chloride (PVC) microplastics and their polyalkylamines modified forms on meiobenthic features in a microcosm. Environ. Pollut. 2020, 266, 115263. [Google Scholar] [CrossRef]
- Page, A.P.; Johnstone, I.L. The cuticle. WormBook. 2007. Available online: http://wormbook.org/chapters/www_cuticle/cuticle.pdf (accessed on 13 February 2022).
- Boufahja, F.; Semprucci, F.; Beyrem, H. An experimental protocol to select nematode species from an entire community using progressive sedimentary enrichment. Ecol. Indic. 2016, 60, 292–309. [Google Scholar] [CrossRef]
- Armenteros, M.; Ruiz-Abierno, A.; Fernández-Garcés, R.; Pérez-García, J.A.; Díaz-Asencio, L.; Vincx, M.; Decraemer, W. Biodiversity patterns of free-living marine nematodes in a tropical bay: Cienfuegos, Caribbean Sea. Estuar. Coast. Shelf Sci. 2009, 85, 179–189. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Case | Structure at Genus Level | Structure at Morpho-Functional Level | ||
|---|---|---|---|---|
| Sample Statistic | Pairwise Tests | Sample Statistic | Pairwise Tests | |
| Genoa Voltri port | Stations R = 0.80; p = 0.001 | St. I vs. St. O: R = 0.99; p = 0.03 | Stations R = 0.78; p = 0.001 | St. I vs. St. O: R = 0,99; p = 0.03 |
| St. I vs. St. M: R = 0.65; p = 0.03 | St. I vs. St. M: R = 0.60; p = 0.03 | |||
| St. O vs. St. M: R = 0.75; p = 0.03 | St. O vs. St. M: R = 0.70; p = 0.03 | |||
| Periods p = n.s. | Periods p = n.s. | |||
| Aregai marina | Stations: p = n.s. | Stations p = n.s. | ||
| Periods R = 0.81; p = 0.029 | Periods p = n.s. | |||
| Riverine outfall areas | Stations R = 0.38; p = 0.001 | F vs. B: R = 0.26; p = 0.05 | Stations R = 0.35; p = 0.001 | F vs. B: R = 0.25; p = 0.05 |
| F vs. FO: R = 0.60; p = 0.002 | F vs. FO: R = 0.59; p = 0.004 | |||
| F vs. M: R = 0.48; p = 0.002 | F vs. M: R = 0.34; p = 0.004 | |||
| B vs. FO: R = 0.67; p = 0.002 | B vs. FO: R = 0.70; p = 0.002 | |||
| B vs. M: R = 0.30; p = 0.017 | B vs. M: R = 0.28; p = 0.017 | |||
| FO vs. M: n.s. | FO vs. M: n.s. | |||
| Periods R = 0.24; p = 0.001 | Periods R = 0.17; p = 0.016 | |||
| Olbia Fish-farm | Stations p = n.s. | Stations p = n.s. | ||
| Periods p = n.s. | Periods p = n.s. | |||
| Vibo Fish-farm | Stations p = n.s. | Stations p = n.s. | ||
| Periods p = n.s. | Periods p = n.s. | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semprucci, F.; Grassi, E.; Balsamo, M. Simple Is the Best: An Alternative Method for the Analysis of Free-Living Nematode Assemblage Structure. Water 2022, 14, 1114. https://doi.org/10.3390/w14071114
Semprucci F, Grassi E, Balsamo M. Simple Is the Best: An Alternative Method for the Analysis of Free-Living Nematode Assemblage Structure. Water. 2022; 14(7):1114. https://doi.org/10.3390/w14071114
Chicago/Turabian StyleSemprucci, Federica, Eleonora Grassi, and Maria Balsamo. 2022. "Simple Is the Best: An Alternative Method for the Analysis of Free-Living Nematode Assemblage Structure" Water 14, no. 7: 1114. https://doi.org/10.3390/w14071114
APA StyleSemprucci, F., Grassi, E., & Balsamo, M. (2022). Simple Is the Best: An Alternative Method for the Analysis of Free-Living Nematode Assemblage Structure. Water, 14(7), 1114. https://doi.org/10.3390/w14071114