
Linking Micropollutants to Trait Syndromes across Freshwater Diatom, Macroinvertebrate, and Fish Assemblages


 ,
,  , and
, and 
Abstract
:1. Introduction
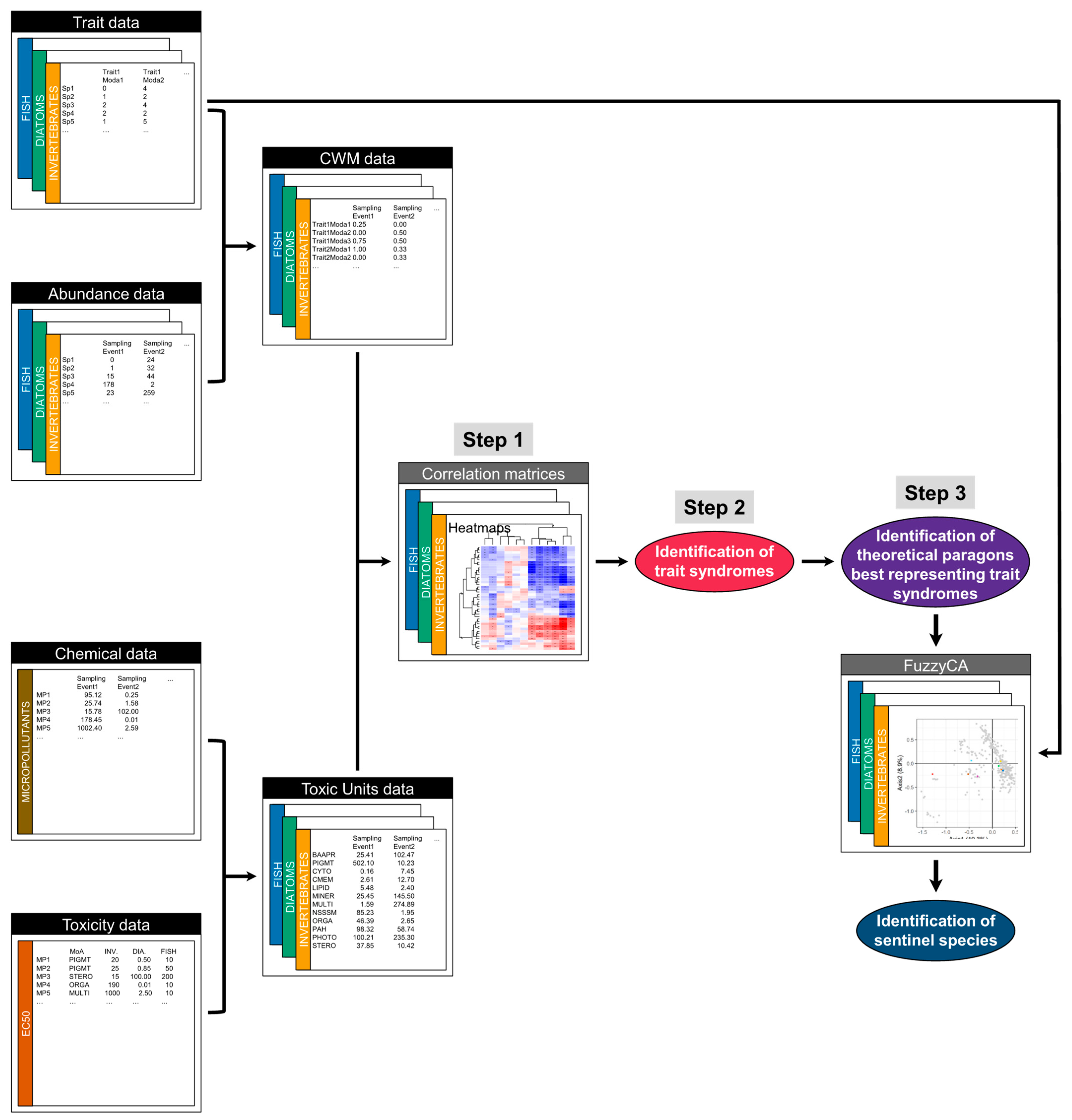
2. Material and Methods
2.1. Benthic Diatom, Macroinvertebrate, and Fish Data
2.2. Calculation of Micropollutant Toxicity in Water
2.3. Statistical Analyses
3. Results
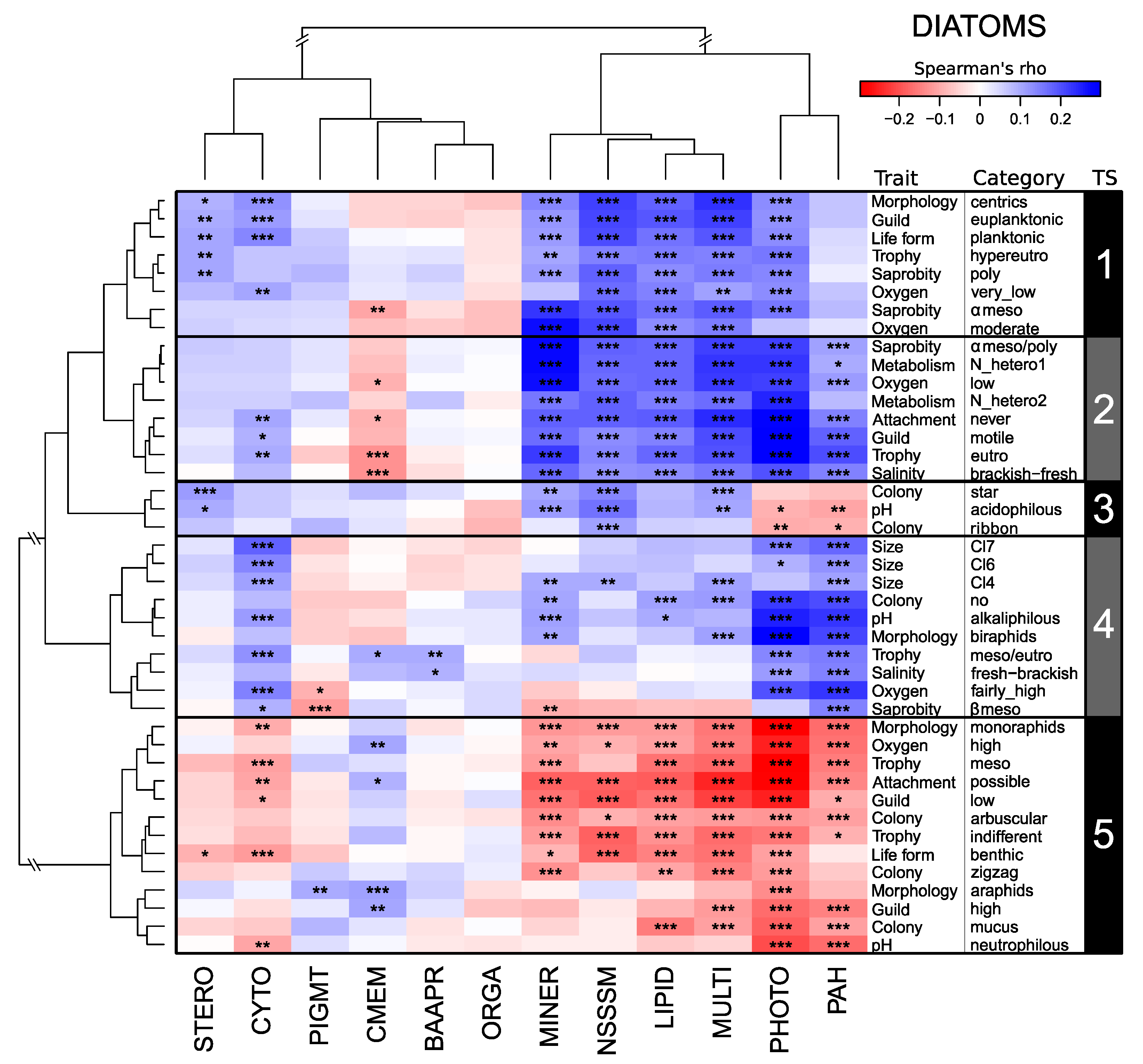
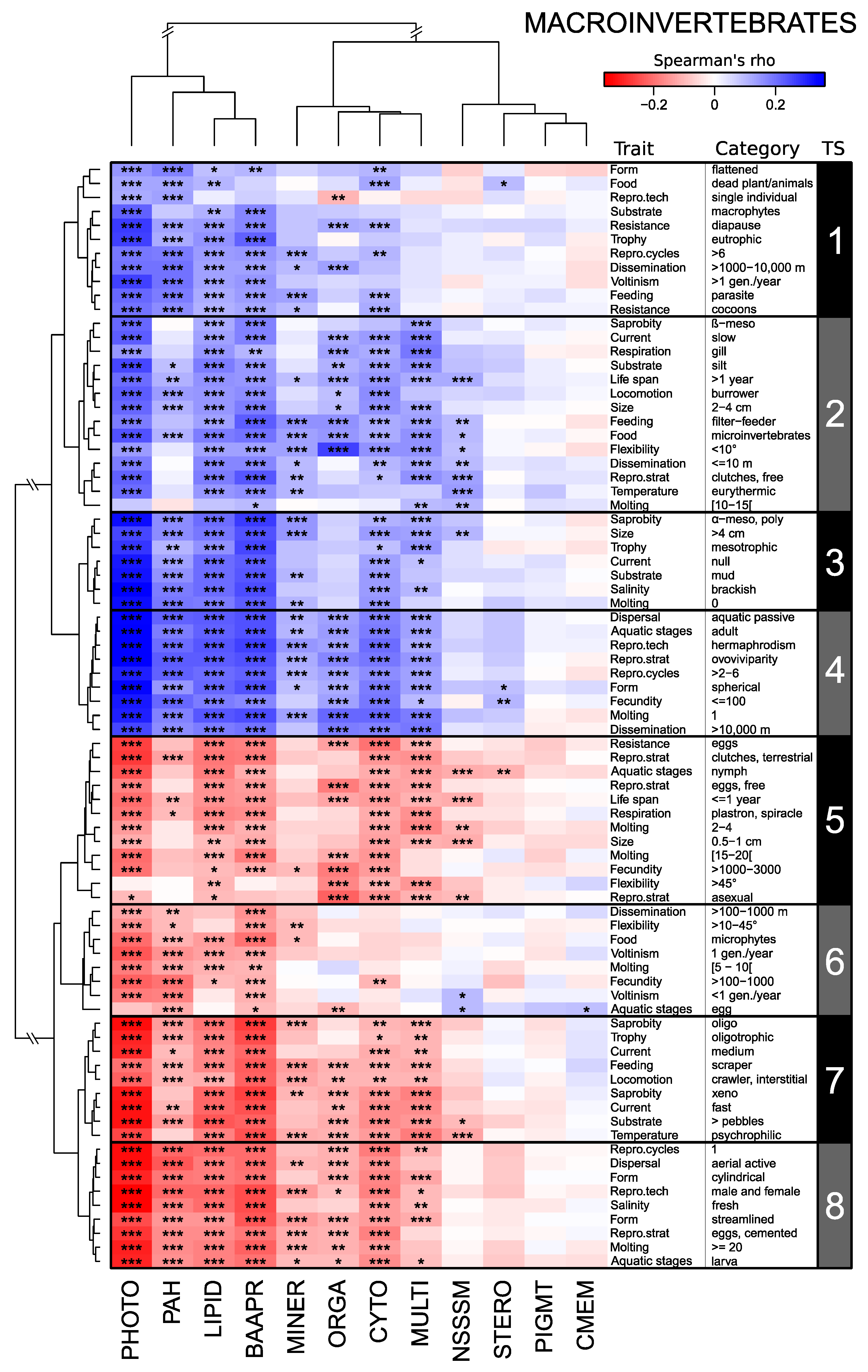
3.1. Links between Trait Syndromes and Modes of Action of Micropollutants
3.2. Trait Syndromes vs. Trait Categories in Explaining the Community–MoA Relationships
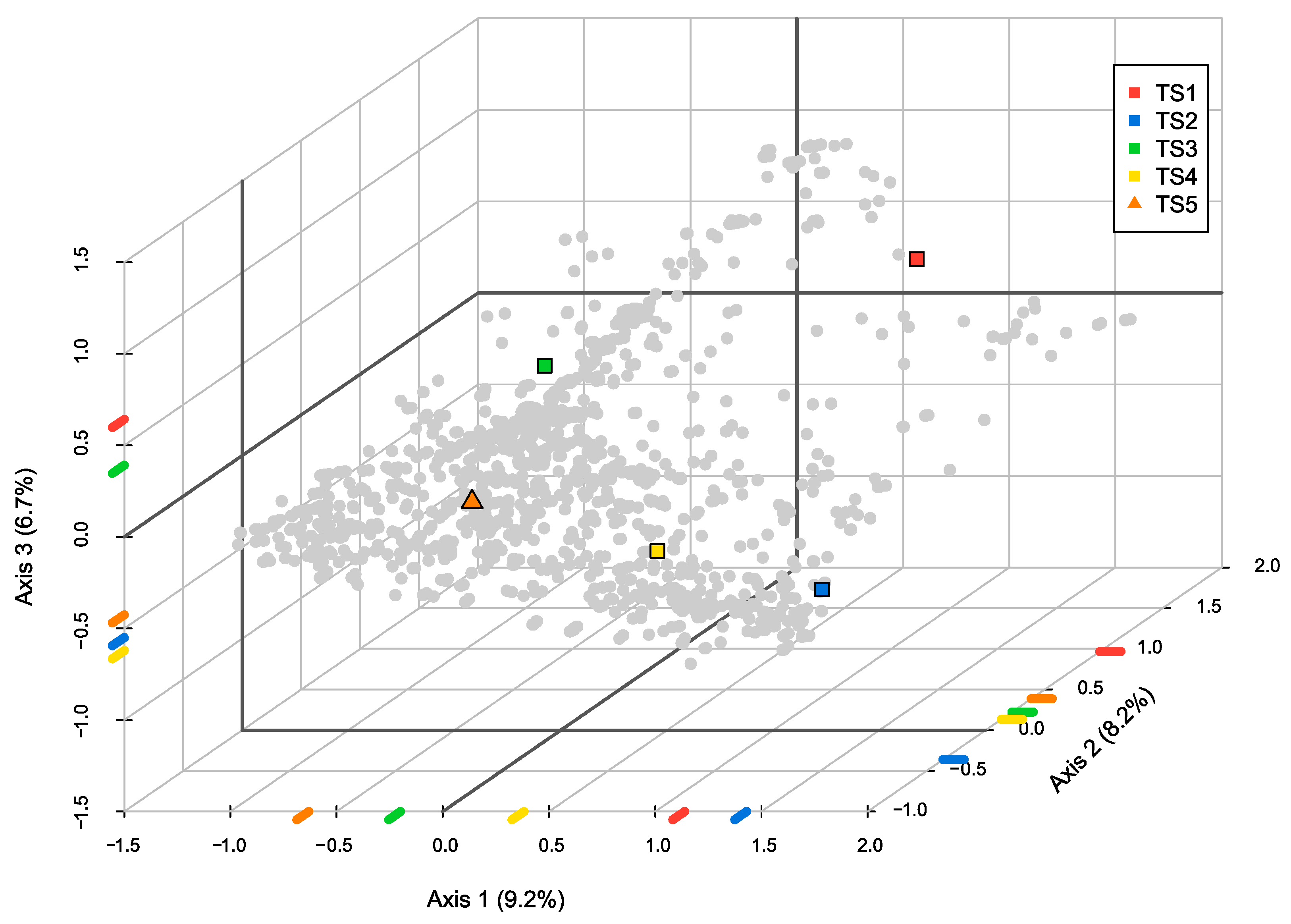
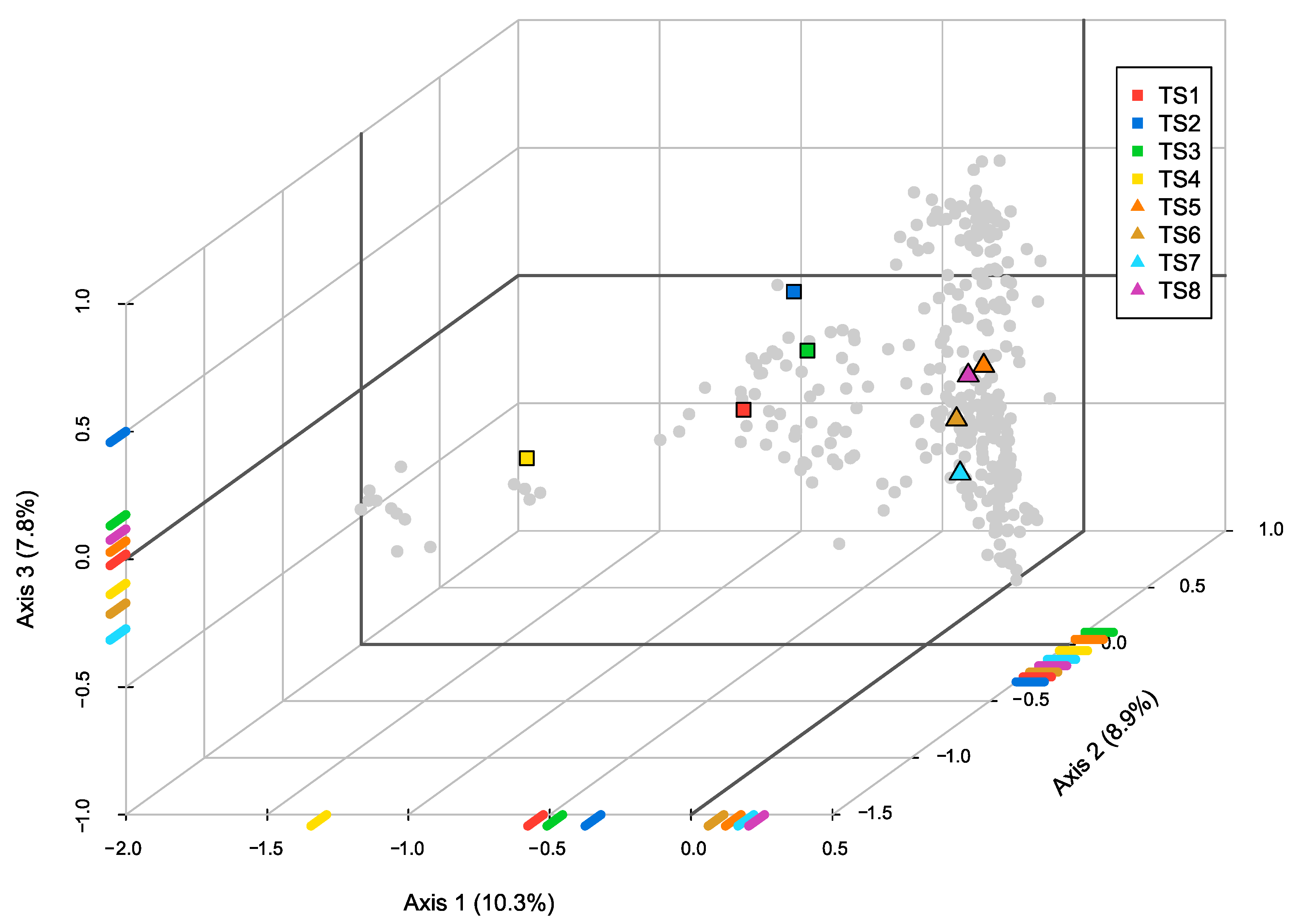
3.3. Paragons as Model Taxa Best Representing Trait Syndromes
4. Discussion
4.1. Trait Syndromes in Diatom, Macroinvertebrate, and Fish Assemblages
4.2. Relevance and Complementarity of Trait Syndromes for Bioassessment
4.3. Key Trait Categories Driving Trait Syndrome Responses
4.4. Paragons as Model Taxa Best Representing Trait Syndromes
5. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matthaei, C.D.; Piggott, J.J.; Townsend, C.R. Multiple stressors in agricultural streams: Interactions among sediment addition, nutrient enrichment and water abstraction. J. Appl. Ecol. 2010, 47, 639–649. [Google Scholar] [CrossRef]
- Luo, K.; Hu, X.; He, Q.; Wu, Z.; Cheng, H.; Hu, Z.; Mazumder, A. Impacts of rapid urbanization on the water quality and macroinvertebrate communities of streams: A case study in Liangjiang New Area, China. Sci. Total Environ. 2018, 621, 1601–1614. [Google Scholar] [CrossRef]
- Embke, H.S.; Rypel, A.L.; Carpenter, S.R.; Sass, G.G.; Ogle, D.; Cichosz, T.; Hennessy, J.; Essington, T.E.; Vander Zanden, M.J. Production dynamics reveal hidden overharvest of inland recreational fisheries. Proc. Natl. Acad. Sci. USA 2019, 116, 24676–24681. [Google Scholar] [CrossRef]
- Haase, P.; Pilotto, F.; Li, F.; Sundermann, A.; Lorenz, A.W.; Tonkin, J.D.; Stoll, S. Moderate warming over the past 25 years has already reorganized stream invertebrate communities. Sci. Total Environ. 2019, 658, 1531–1538. [Google Scholar] [CrossRef] [PubMed]
- Feld, C.K.; Hering, D. Community structure or function: Effects of environmental stress on benthic macroinvertebrates at different spatial scales. Freshw. Biol. 2007, 52, 1380–1399. [Google Scholar] [CrossRef]
- Tóth, R.; Czeglédi, I.; Kern, B.; Erős, T. Land use effects in riverscapes: Diversity and environmental drivers of stream fish communities in protected, agricultural and urban landscapes. Ecol. Indic. 2019, 101, 742–748. [Google Scholar] [CrossRef] [Green Version]
- Schwarzenbach, R.P.; Escher, B.I.; Fenner, K.; Hofstetter, T.B.; Johnson, C.A.; von Gunten, U.; Wehrli, B. The Challenge of Micropollutants in Aquatic Systems. Science 2006, 313, 1072–1077. [Google Scholar] [CrossRef]
- Persson, L.; Carney Almroth, B.M.; Collins, C.D.; Cornell, S.; de Wit, C.A.; Diamond, M.L.; Fantke, P.; Hassellöv, M.; MacLeod, M.; Ryberg, M.W.; et al. Outside the Safe Operating Space of the Planetary Boundary for Novel Entities. Environ. Sci. Technol. 2022, 56, 1510–1521. [Google Scholar] [CrossRef]
- Tang, J.Y.M.; McCarty, S.; Glenn, E.; Neale, P.A.; Warne, M.S.J.; Escher, B.I. Mixture effects of organic micropollutants present in water: Towards the development of effect-based water quality trigger values for baseline toxicity. Water Res. 2013, 47, 3300–3314. [Google Scholar] [CrossRef]
- Ankley, G.T.; Bennett, R.S.; Erickson, R.J.; Hoff, D.J.; Hornung, M.W.; Johnson, R.D.; Mount, D.R.; Nichols, J.W.; Russom, C.L.; Schmieder, P.K.; et al. Adverse outcome pathways: A conceptual framework to support ecotoxicology research and risk assessment. Environ. Toxicol. Chem. 2010, 29, 730–741. [Google Scholar] [CrossRef]
- Ankley, G.T.; Edwards, S.W. The adverse outcome pathway: A multifaceted framework supporting 21st century toxicology. Curr. Opin. Toxicol. 2018, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.E.; Vikstrom, S.; König, M.; Schlichting, R.; Zarfl, C.; Zwiener, C.; Escher, B.I. Mitochondrial Toxicity of Selected Micropollutants, Their Mixtures, and Surface Water Samples Measured by the Oxygen Consumption Rate in Cells. Environ. Toxicol. Chem. 2019, 38, 1000–1011. [Google Scholar] [CrossRef]
- Schäfer, R.B.; Gerner, N.; Kefford, B.J.; Rasmussen, J.J.; Beketov, M.A.; de Zwart, D.; Liess, M.; von der Ohe, P.C. How to characterize chemical exposure to predict ecologic effects on aquatic communities? Environ. Sci. Technol. 2013, 47, 7996–8004. [Google Scholar] [CrossRef] [PubMed]
- Neale, P.A.; Altenburger, R.; Aït-Aïssa, S.; Brion, F.; Busch, W.; de Aragão Umbuzeiro, G.; Denison, M.S.; Du Pasquier, D.; Hilscherová, K.; Hollert, H.; et al. Development of a bioanalytical test battery for water quality monitoring: Fingerprinting identified micropollutants and their contribution to effects in surface water. Water Res. 2017, 123, 734–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, N.P.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Novel understanding of toxicity in a life cycle perspective–The mechanisms that lead to population effect–The case of Ag (nano)materials. Environ. Pollut. 2020, 262, 114277. [Google Scholar] [CrossRef]
- Busch, W.; Schmidt, S.; Kühne, R.; Schulze, T.; Krauss, M.; Altenburger, R. Micropollutants in European rivers: A mode of action survey to support the development of effect-based tools for water monitoring. Environ. Toxicol. Chem. 2016, 35, 1887–1899. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Chen, Z.; Hollert, H.; Zhou, S.; Deutschmann, B.; Seiler, T.-B. Toxicity of 10 organic micropollutants and their mixture: Implications for aquatic risk assessment. Sci. Total Environ. 2019, 666, 1273–1282. [Google Scholar] [CrossRef]
- Baattrup-Pedersen, A.; Göthe, E.; Riis, T.; O’Hare, M.T. Functional trait composition of aquatic plants can serve to disentangle multiple interacting stressors in lowland streams. Sci. Total Environ. 2016, 543, 230–238. [Google Scholar] [CrossRef]
- Mondy, C.P.; Muñoz, I.; Dolédec, S. Life-history strategies constrain invertebrate community tolerance to multiple stressors: A case study in the Ebro basin. Sci. Total Environ. 2016, 572, 196–206. [Google Scholar] [CrossRef]
- Waite, I.R.; Munn, M.D.; Moran, P.W.; Konrad, C.P.; Nowell, L.H.; Meador, M.R.; Van Metre, P.C.; Carlisle, D.M. Effects of urban multi-stressors on three stream biotic assemblages. Sci. Total Environ. 2019, 660, 1472–1485. [Google Scholar] [CrossRef]
- Alric, B.; Dézerald, O.; Meyer, A.; Billoir, E.; Coulaud, R.; Larras, F.; Mondy, C.P.; Usseglio-Polatera, P. How diatom-, invertebrate- and fish-based diagnostic tools can support the ecological assessment of rivers in a multi-pressure context: Temporal trends over the past two decades in France. Sci. Total Environ. 2021, 762, 143915. [Google Scholar] [CrossRef] [PubMed]
- Mondy, C.P.; Usseglio-Polatera, P. Using conditional tree forests and life history traits to assess specific risks of stream degradation under multiple pressure scenario. Sci. Total Environ. 2013, 461–462, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Larras, F.; Coulaud, R.; Gautreau, E.; Billoir, E.; Rosebery, J.; Usseglio-Polatera, P. Assessing anthropogenic pressures on streams: A random forest approach based on benthic diatom communities. Sci. Total Environ. 2017, 586, 1101–1112. [Google Scholar] [CrossRef]
- Dézerald, O.; Mondy, C.P.; Dembski, S.; Kreutzenberger, K.; Reyjol, Y.; Chandesris, A.; Valette, L.; Brosse, S.; Toussaint, A.; Belliard, J.; et al. A diagnosis-based approach to assess specific risks of river degradation in a multiple pressure context: Insights from fish communities. Sci. Total Environ. 2020, 734, 139467. [Google Scholar] [CrossRef] [PubMed]
- Southwood, T.R.E. Habitat, the Templet for Ecological Strategies? J. Anim. Ecol. 1977, 46, 336. [Google Scholar] [CrossRef]
- Southwood, T.R.E. Tactics, Strategies and Templets. Oikos 1988, 52, 3–18. [Google Scholar] [CrossRef]
- Townsend, C.R.; Hildrew, A.G. Species traits in relation to a habitat templet for river systems. Freshw. Biol. 1994, 31, 265–275. [Google Scholar] [CrossRef]
- Keddy, P.A. Assembly and response rules: Two goals for predictive community ecology. J. Veg. Sci. 1992, 3, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Diamond, J.M. Assembly of species communities. In Ecology and Evolution of Communities; Cody, M.L., Diamond, J.M., Eds.; Harvard University Press: Cambridge, MA, USA, 1975; pp. 342–444. [Google Scholar]
- Resh, V.H.; Hildrew, A.G.; Statzner, B.; Townsend, C.R. Theoretical habitat templets, species traits, and species richness: A synthesis of long-term ecological research on the Upper Rhône River in the context of concurrently developed ecological theory. Freshw. Biol. 1994, 31, 539–554. [Google Scholar] [CrossRef]
- Statzner, B.; Bêche, L.A. Can biological invertebrate traits resolve effects of multiple stressors on running water ecosystems? Freshw. Biol. 2010, 55, 80–119. [Google Scholar]
- Verberk, W.C.E.P.; van Noordwijk, C.G.E.; Hildrew, A.G. Delivering on a promise: Integrating species traits to transform descriptive community ecology into a predictive science. Freshw. Sci. 2013, 32, 531–547. [Google Scholar] [CrossRef] [Green Version]
- Arce, E.; Archaimbault, V.; Mondy, C.P.; Usseglio-Polatera, P. Recovery dynamics in invertebrate communities following water-quality improvement: Taxonomy-vs trait-based assessment. Freshw. Sci. 2014, 33, 1060–1073. [Google Scholar] [CrossRef] [Green Version]
- Usseglio-Polatera, P.; Bournaud, M.; Richoux, P.; Tachet, H. Biological and ecological traits of benthic freshwater macroinvertebrates: Relationships and definition of groups with similar traits. Freshw. Biol. 2000, 43, 175–205. [Google Scholar] [CrossRef]
- Poff, N.L.; Olden, J.D.; Vieira, N.K.M.; Finn, D.S.; Simmons, M.P.; Kondratieff, B.C. Functional trait niches of North American lotic insects: Traits-based ecological applications in light of phylogenetic relationships. J. N. Am. Benthol. Soc. 2006, 25, 730–755. [Google Scholar] [CrossRef] [Green Version]
- Menezes, S.; Baird, D.J.; Soares, A.M.V.M. Beyond taxonomy: A review of macroinvertebrate trait-based community descriptors as tools for freshwater biomonitoring: Trait-based community descriptors. J. Appl. Ecol. 2010, 47, 711–719. [Google Scholar] [CrossRef]
- Statzner, B.; Hoppenhaus, K.; Arens, M.; Richoux, P. Reproductive traits, habitat use and templet theory: A synthesis of world-wide data on aquatic insects. Freshw. Biol. 1997, 38, 109–135. [Google Scholar] [CrossRef]
- Stearns, S.C. The Evolution of Life Histories; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Usseglio-Polatera, P.; Richoux, P.; Bournaud, M.; Tachet, H. A functional classification of benthic macroinvertebrates based on biological and ecological traits: Application to river condition assessment and stream management. Arch. Hydrobiol. 2001, 139, 53–83. [Google Scholar]
- Hering, D.; Johnson, R.K.; Kramm, S.; Schmutz, S.; Szoszkiewicz, K.; Verdonschot, P.F.M. Assessment of European streams with diatoms, macrophytes, macroinvertebrates and fish: A comparative metric-based analysis of organism response to stress. Freshw. Biol. 2006, 51, 1757–1785. [Google Scholar] [CrossRef]
- Johnson, R.K.; Hering, D.; Furse, M.T.; Clarke, R.T. Detection of ecological change using multiple organism groups: Metrics and uncertainty. Hydrobiologia 2006, 566, 115–137. [Google Scholar] [CrossRef]
- Hering, D.; Johnson, R.K.; Buffagni, A. Linking organism groups–major results and conclusions from the STAR project. Hydrobiologia 2006, 566, 109–113. [Google Scholar] [CrossRef]
- European Council. Directive 2000/60/EC. Establishing a Framework for Community Action in the Field of Water Policy; European Commission PE-CONS 3639/1/100Rev 1; European Council: Bruxelles, Belgium, 2000. [Google Scholar]
- AFNOR. Qualité de L’eau–Détermination de L’indice Biologique Diatomées (IBD); NF T90-354; AFNOR: Paris, France, 2000. [Google Scholar]
- AFNOR. Qualité de L’eau–Prélèvement des Macro-Invertébrés Aquatiques en Rivières peu Profondes; XP T90-333; AFNOR: Paris, France, 2009. [Google Scholar]
- AFNOR. Qualité de L’eau–Échantillonnage des Poissons à L’électricité; NF EN 14011; AFNOR: Paris, France, 2003. [Google Scholar]
- Mondy, C.P.; Villeneuve, B.; Archaimbault, V.; Usseglio-Polatera, P. A new macroinvertebrate-based multimetric index (I2M2) to evaluate ecological quality of French wadeable streams fulfilling the WFD demands: A taxonomical and trait approach. Ecol. Indic. 2012, 18, 452–467. [Google Scholar] [CrossRef]
- Tachet, H.; Richoux, P.; Bournaud, M.; Usseglio-Polatera, P. Invertébrés d’eau Douce: Systématique, Biologie, Ecologie, 2nd ed.; CNRS Éditions: Paris, France, 2010. [Google Scholar]
- Beck, M.; Mondy, C.P.; Danger, M.; Billoir, E.; Usseglio-Polatera, P. Extending the growth rate hypothesis to species development: Can stoichiometric traits help to explain the composition of macroinvertebrate communities? Oikos 2021, 130, 879–892. [Google Scholar] [CrossRef]
- Chevenet, F.; Dolédec, S.; Chessel, D. A fuzzy coding approach for the analysis of long-term ecological data. Freshw. Biol. 1994, 31, 295–309. [Google Scholar] [CrossRef]
- Malaj, E.; von der Ohe, P.C.; Grote, M.; Kühne, R.; Mondy, C.P.; Usseglio-Polatera, P.; Brack, W.; Schäfer, R.B. Organic chemicals jeopardize the health of freshwater ecosystems on the continental scale. Proc. Natl. Acad. Sci. USA 2014, 111, 9549–9554. [Google Scholar] [CrossRef] [Green Version]
- R4P. Universal Classification of PPPs/Classification Universelle des PPP. 2019. Available online: https://www.r4p-inra.fr/fr/r4p-propose-une-classification-universelle/ (accessed on 1 March 2021).
- Puth, M.-T.; Neuhäuser, M.; Ruxton, G.D. Effective use of Spearman’s and Kendall’s correlation coefficients for association between two measured traits. Anim. Behav. 2015, 102, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.H. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Sneath, P.H.A.; Sokal, R.R. Numerical Taxonomy: The Principles and Practice of Numerical Classification; W.H. Freeman Co.: London, UK, 1973. [Google Scholar]
- Hastie, T.; Tibshirani, R. Generalized Additive Models; Chapman and Hall: London, UK, 1990. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. gplots. Various R Programming Tools for Plotting Data. 2020. Available online: https://CRAN.R-project.org/package=gplots (accessed on 1 March 2021).
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman & Hall/CRC Press: London, UK, 2017. [Google Scholar]
- Dray, S.; Dufour, A.-B. The ade4 Package: Implementing the Duality Diagram for Ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Berger, E.; Haase, P.; Schäfer, R.B.; Sundermann, A. Towards stressor-specific macroinvertebrate indices: Which traits and taxonomic groups are associated with vulnerable and tolerant taxa? Sci. Total Environ. 2018, 619–620, 144–154. [Google Scholar] [CrossRef]
- Tison-Rosebery, J.; Leboucher, T.; Archaimbault, V.; Belliard, J.; Carayon, D.; Ferréol, M.; Floury, M.; Jeliazkov, A.; Tales, E.; Villeneuve, B.; et al. Decadal biodiversity trends in rivers reveal recent community rearrangements. Sci. Total Environ. 2022, 823, 153431. [Google Scholar]
- Marcel, R.; Berthon, V.; Castets, V.; Rimet, F.; Thiers, A.; Labat, F.; Fontan, B. Modelling diatom life forms and ecological guilds for river biomonitoring. Knowl. Manag. Aquat. Ecosyst. 2017, 418, 1. [Google Scholar] [CrossRef] [Green Version]
- Wood, R.J.; Mitrovic, S.M.; Lim, R.P.; Warne, M.S.J.; Dunlop, J.; Kefford, B.J. Benthic diatoms as indicators of herbicide toxicity in rivers–A new SPEcies at Risk (SPEARherbicides) index. Ecol. Indic. 2019, 99, 203–213. [Google Scholar] [CrossRef]
- Larras, F.; Keck, F.; Montuelle, B.; Rimet, F.; Bouchez, A. Linking Diatom Sensitivity to Herbicides to Phylogeny: A Step Forward for Biomonitoring? Environ. Sci. Technol. 2014, 48, 1921–1930. [Google Scholar] [CrossRef] [PubMed]
- Marcel, R.; Bouchez, A.; Rimet, F. Influence of Herbicide Contamination on Diversity and Ecological Guilds of River Diatoms. Cryptogam. Algol. 2013, 34, 169–183. [Google Scholar] [CrossRef]
- Garali, S.M.B.; Sahraoui, I.; Othman, H.B.; Kouki, A.; de la Iglesia, P.; Diogène, J.; Lafabrie, C.; Andree, K.B.; Fernández-Tejedor, M.; Mejri, K.; et al. Capacity of the potentially toxic diatoms Pseudo-nitzschia mannii and Pseudo-nitzschia hasleana to tolerate polycyclic aromatic hydrocarbons. Ecotoxicol. Environ. Saf. 2021, 214, 112082. [Google Scholar] [CrossRef] [PubMed]
- Usseglio-Polatera, P.; Beisel, J.-N. Longitudinal changes in macroinvertebrate assemblages in the Meuse River: Anthropogenic effects versus natural change. River Res. Appl. 2002, 18, 197–211. [Google Scholar] [CrossRef]
- Winemiller, K.O.; Fitzgerald, D.B.; Bower, L.M.; Pianka, E.R. Functional traits, convergent evolution, and periodic tables of niches. Ecol. Lett. 2015, 18, 737–751. [Google Scholar] [CrossRef]
- Comte, L.; Olden, J.D. Evidence for dispersal syndromes in freshwater fishes. Proc. R. Soc. B Biol. Sci. 2018, 285, 20172214. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, R.B.; van den Brink, P.J.; Liess, M. Impacts of pesticides on freshwater ecosystems. In Ecological Impacts of Toxic Chemicals; Sánchez-Bayo, F., van den Brink, P.J., Mann, R.M., Eds.; Bentham Science Publishers: Charjah, United Arab Emirates, 2011; pp. 111–137. [Google Scholar]
- Shahid, N.; Becker, J.M.; Krauss, M.; Brack, W.; Liess, M. Pesticide Body Burden of the Crustacean Gammarus pulex as a Measure of Toxic Pressure in Agricultural Streams. Environ. Sci. Technol. 2018, 52, 7823–7832. [Google Scholar] [CrossRef]
- Liess, M.; Henz, S.; Knillmann, S. Predicting low-concentration effects of pesticides. Sci. Rep. 2019, 9, 15248. [Google Scholar] [CrossRef] [Green Version]
- Liess, M.; Henz, S.; Shahid, N. Modeling the synergistic effects of toxicant mixtures. Environ. Sci. Eur. 2020, 32, 119. [Google Scholar] [CrossRef]
- Toussaint, A.; Charpin, N.; Brosse, S.; Villéger, S. Global functional diversity of freshwater fish is concentrated in the Neotropics while functional vulnerability is widespread. Sci. Rep. 2016, 6, 22125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamsi, O.; Pinelli, E.; Faucon, B.; Perrault, A.; Lacroix, L.; Sánchez-Pérez, J.-M.; Charcosset, J.-Y. Effects of herbicide mixtures on freshwater microalgae with the potential effect of a safener. Int. J. Limnol. 2019, 55, 3. [Google Scholar] [CrossRef] [Green Version]
- Larras, F.; Bouchez, A.; Rimet, F.; Montuelle, B. Using bioassays and species sensitivity distributions to assess herbicide toxicity towards benthic diatoms. PLoS ONE 2012, 7, e44458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reavie, E.D.; Cai, M. Consideration of species-specific diatom indicators of anthropogenic stress in the Great Lakes. PLoS ONE 2019, 14, e0210927. [Google Scholar] [CrossRef] [Green Version]
- Vidal, T.; Santos, M.; Santos, J.I.; Luís, T.; Pereira, M.J.; Abrantes, N.; Gonçalves, F.J.M.; Pereira, J.L. Testing the response of benthic diatom assemblages to common riverine contaminants. Sci. Total Environ. 2021, 755, 142534. [Google Scholar] [CrossRef]
- Esteves, S.M.; Keck, F.; Almeida, S.F.P.; Figueira, E.; Bouchez, A.; Rimet, F. Can we predict diatoms herbicide sensitivities with phylogeny? Influence of intraspecific and interspecific variability. Ecotoxicology 2017, 26, 1065–1077. [Google Scholar] [CrossRef]
- Cantonati, M.; Lange-Bertalot, H. Diatom monitors of close-to-pristine, very-low alkalinity habitats: Three new Eunotia species from springs in Nature Parks of the south-eastern Alps. J. Limnol. 2011, 70, 209–221. [Google Scholar] [CrossRef]
- Amorim, J.; Abreu, I.; Rodrigues, P.; Peixoto, D.; Pinheiro, C.; Saraiva, A.; Carvalho, A.P.; Guimarães, L.; Oliva-Teles, L. Lymnaea stagnalis as a freshwater model invertebrate for ecotoxicological studies. Sci. Total Environ. 2019, 669, 11–28. [Google Scholar] [CrossRef]
- Kunz, P.Y.; Kienle, C.; Gerhardt, A. Gammarus spp. in Aquatic Ecotoxicology and Water Quality Assessment: Toward Integrated Multilevel Tests. In Reviews of Environmental Contamination and Toxicology; Whitacre, D.M., Ed.; Springer: New York, NY, USA, 2010; Volume 205, pp. 1–76. [Google Scholar]
- Chaumot, A.; Geffard, O.; Armengaud, J.; Maltby, L. Gammarids as Reference Species for Freshwater Monitoring. In Aquatic Ecotoxicology; Elsevier: Amsterdam, The Netherlands, 2015; pp. 253–280. [Google Scholar]
- Binelli, A.; Della Torre, C.; Magni, S.; Parolini, M. Does zebra mussel (Dreissena polymorpha) represent the freshwater counterpart of Mytilus in ecotoxicological studies? A critical review. Environ. Pollut. 2015, 196, 386–403. [Google Scholar] [CrossRef]
- Palos-Ladeiro, M.; Barjhoux, I.; Bigot-Clivot, A.; Bonnard, M.; David, E.; Dedourge-Geffard, O.; Geba, E.; Lance, E.; Lepretre, M.; Magniez, G.; et al. Mussel as a Tool to Define Continental Watershed Quality. In Organismal and Molecular Malacology; Ray, S., Ed.; InTechOpen: London, UK, 2017. [Google Scholar]
- Guo, X.; Feng, C. Biological toxicity response of Asian Clam (Corbicula fluminea) to pollutants in surface water and sediment. Sci. Total Environ. 2018, 631, 56–70. [Google Scholar] [CrossRef]
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish, 2nd ed.; EPA 841-B-99-002; Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999.
- Ducrot, V.; Usseglio-Polatera, P.; Péry, A.R.R.; Mouthon, J.; Lafont, M.; Roger, M.-C.; Garric, J.; Férard, J.-F. Using aquatic macroinvertebrate species traits to build test batteries for sediment toxicity assessment: Accounting for the diversity of potential biological responses to toxicants. Environ. Toxicol. Chem. 2005, 24, 2306–2315. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Huerfano, F.; Gómez-Oliván, L.M.; Luja-Mondragón, M.; SanJuan-Reyes, N.; Islas-Flores, H.; Hernández-Navarro, M.-D. Embryotoxic and teratogenic profile of tretracycline at environmentally relevant concentrations on Cyprinus carpio. Chemosphere 2020, 240, 124969. [Google Scholar] [CrossRef] [PubMed]
- Rigaud, C.; Härme, J.; Vehniäinen, E.-R. Salmo trutta is more sensitive than Oncorhynchus mykiss to early-life stage exposure to retene. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 252, 109219. [Google Scholar] [CrossRef] [PubMed]
- Crowley, D.; Penk, M.R.; Macaulay, S.J.; Piggott, J.J. Acute toxicity of the insecticide cypermethrin to three common European mayfly and stonefly nymphs. Limnologica 2021, 88, 125871. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Target Function or System (or Name of Pollutants) | Number of Compounds Involved | Code |
|---|---|---|
| Pesticides | ||
| Biosynthesis of amino acids and proteins | 5 | BAAPR |
| Biosynthesis of nucleic acids and precursors | 1 | * |
| Biosynthesis of pigments | 3 | PIGMT |
| Cell division or cytoskeleton | 3 | CYTO |
| Cell membrane integrity | 2 | CMEM |
| Cell signaling | 3 | * |
| Hormonal regulation | 4 | * |
| Lipid metabolism | 4 | LIPID |
| Multi-sites, multi-targets | 4 | MULTI |
| Nervous system, sensory system or muscles | 18 | NSSSM |
| Photosynthesis | 15 | PHOTO |
| Plant defense stimulation | 1 | * |
| Sterol metabolism | 6 | STERO |
| Other micropollutants | ||
| (other) Organic micropollutants | 47 | ORGA |
| PAH | 8 | PAH |
| PCB | 1 | * |
| Mineral micropollutants | 5 | MINER |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meyer, A.; Alric, B.; Dézerald, O.; Billoir, E.; Coulaud, R.; Larras, F.; Mondy, C.P.; Usseglio-Polatera, P. Linking Micropollutants to Trait Syndromes across Freshwater Diatom, Macroinvertebrate, and Fish Assemblages. Water 2022, 14, 1184. https://doi.org/10.3390/w14081184
Meyer A, Alric B, Dézerald O, Billoir E, Coulaud R, Larras F, Mondy CP, Usseglio-Polatera P. Linking Micropollutants to Trait Syndromes across Freshwater Diatom, Macroinvertebrate, and Fish Assemblages. Water. 2022; 14(8):1184. https://doi.org/10.3390/w14081184
Chicago/Turabian StyleMeyer, Albin, Benjamin Alric, Olivier Dézerald, Elise Billoir, Romain Coulaud, Floriane Larras, Cédric P. Mondy, and Philippe Usseglio-Polatera. 2022. "Linking Micropollutants to Trait Syndromes across Freshwater Diatom, Macroinvertebrate, and Fish Assemblages" Water 14, no. 8: 1184. https://doi.org/10.3390/w14081184
APA StyleMeyer, A., Alric, B., Dézerald, O., Billoir, E., Coulaud, R., Larras, F., Mondy, C. P., & Usseglio-Polatera, P. (2022). Linking Micropollutants to Trait Syndromes across Freshwater Diatom, Macroinvertebrate, and Fish Assemblages. Water, 14(8), 1184. https://doi.org/10.3390/w14081184