
Relationships between Physico-Chemical Parameters and Taxonomic Structure of Benthic Macroinvertebrate Assemblages in Streams of West Cameroon



Abstract
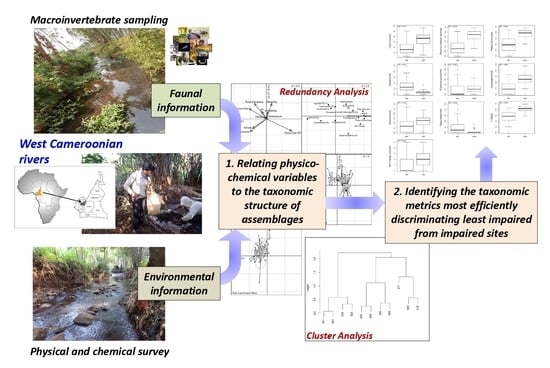
:
1. Introduction
2. Material and Methods
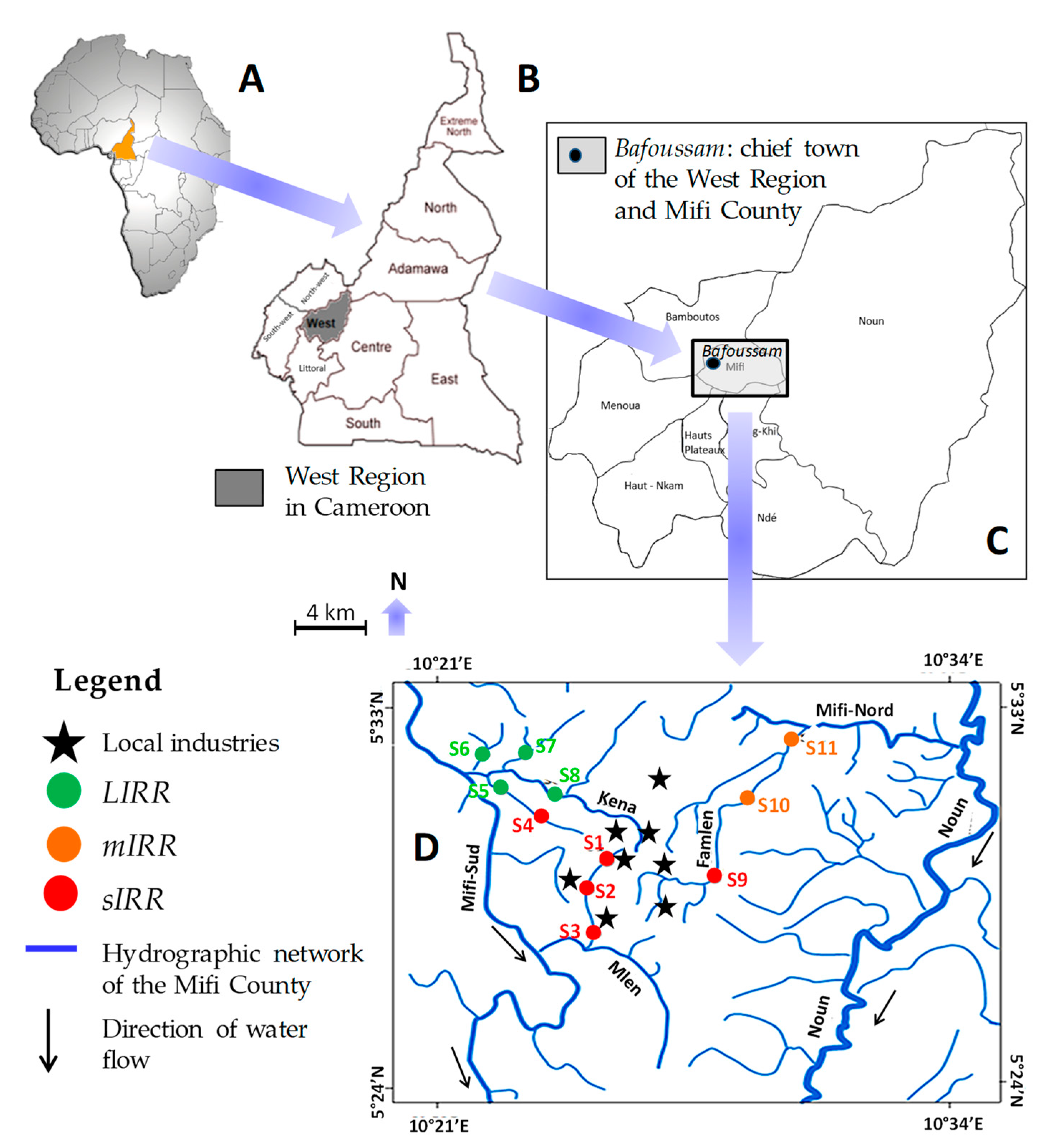
2.1. Study Area and Human Pressure-Based Typology of Sites
2.2. Benthic Macroinvertebrate Sampling and Community Structure Description
2.3. Physico-Chemical Parameters
2.4. Environmental and Faunal Data Analyses
3. Results
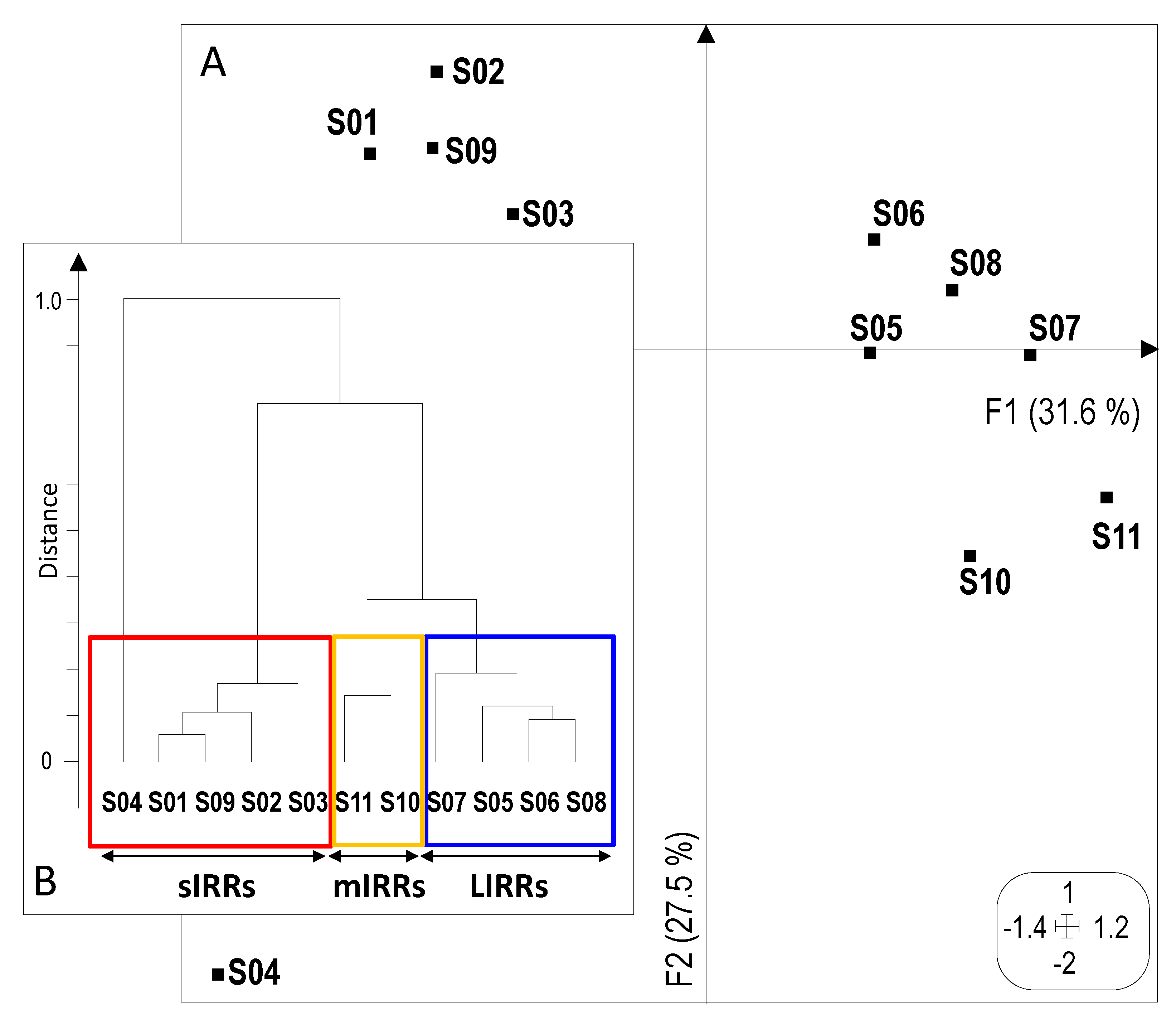
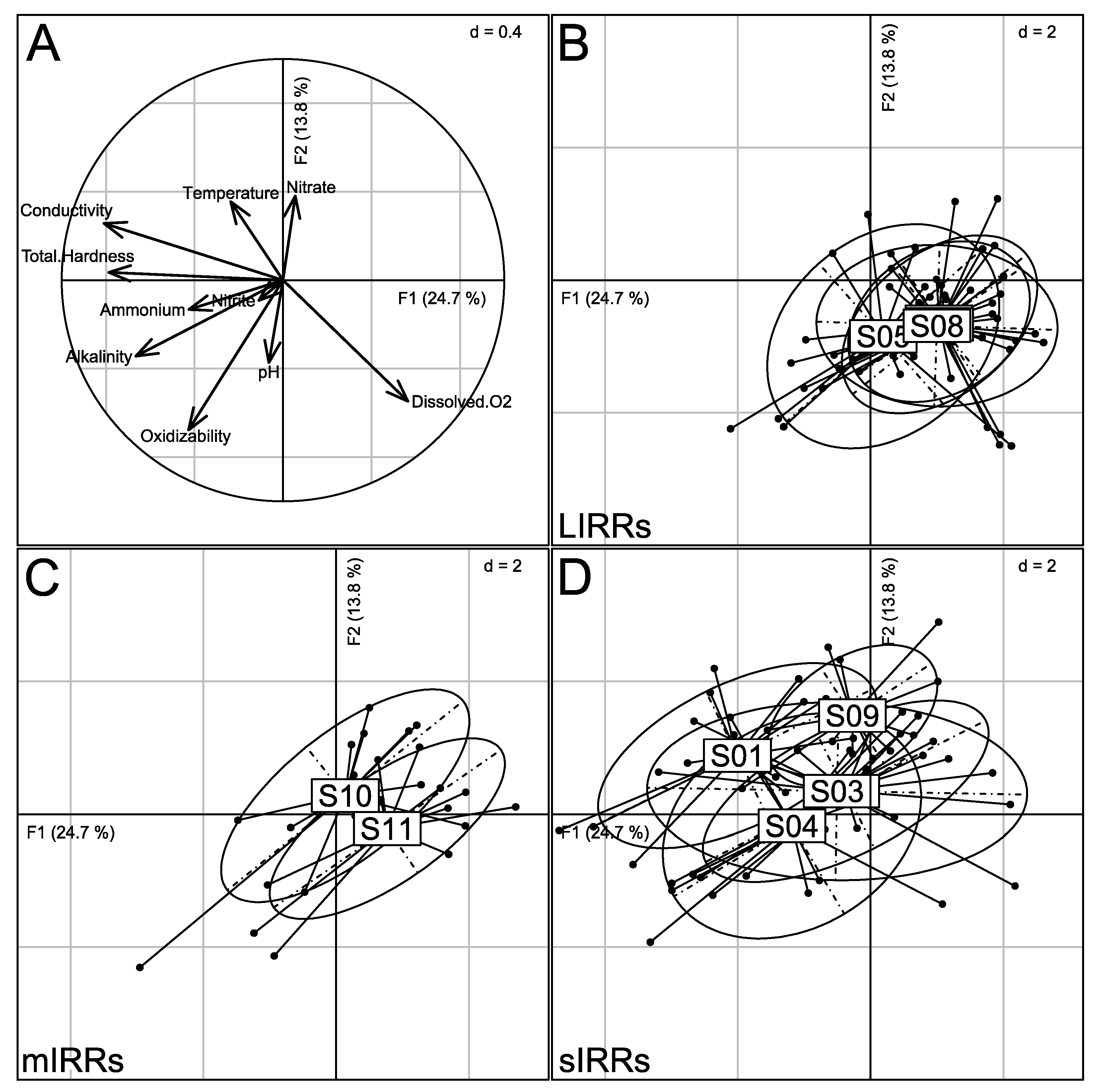
3.1. Human Pressure-Based Typology of Sites
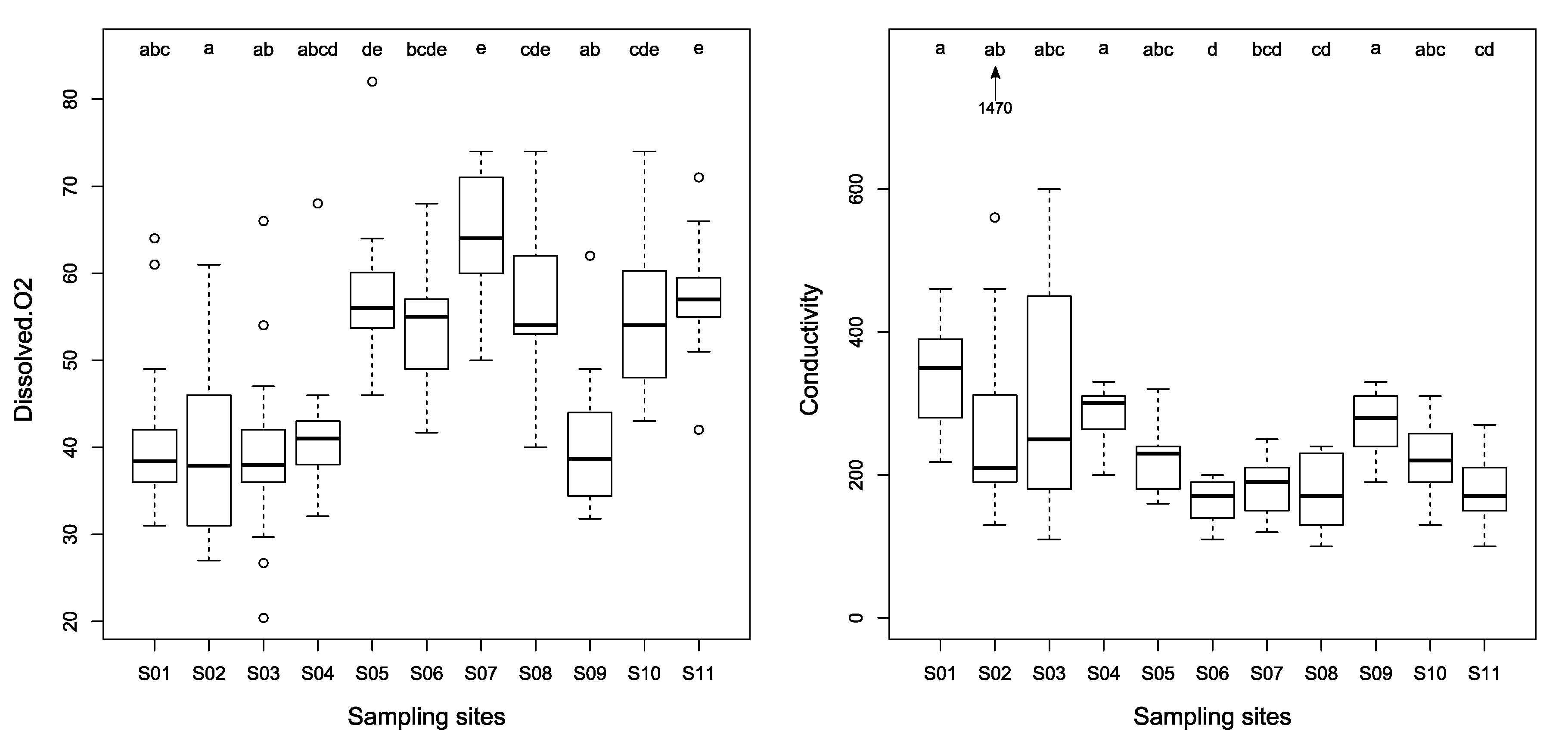
3.2. Physico-Chemical Characteristics of Streams
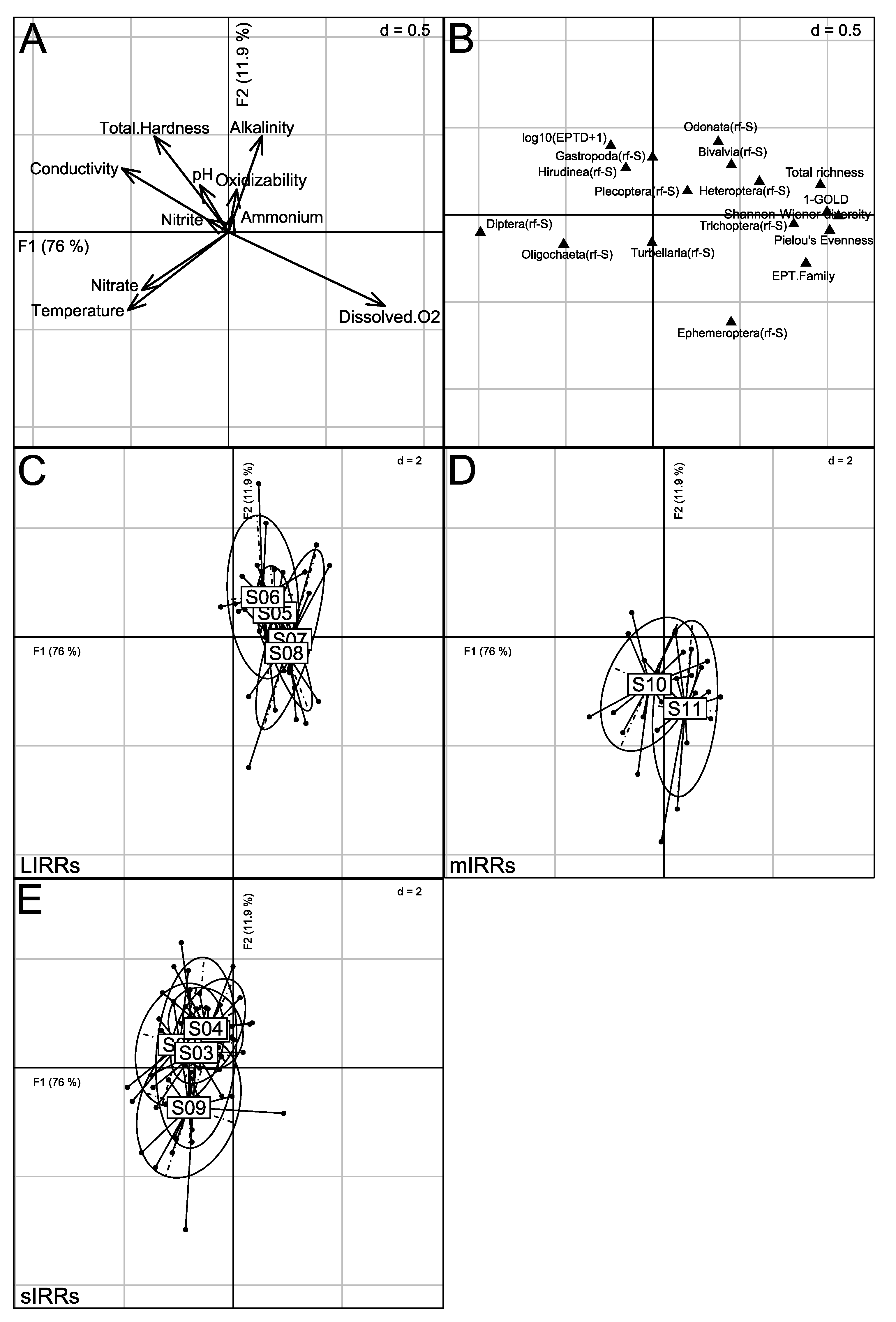
3.3. Abiotic Typology of Sampling Sites
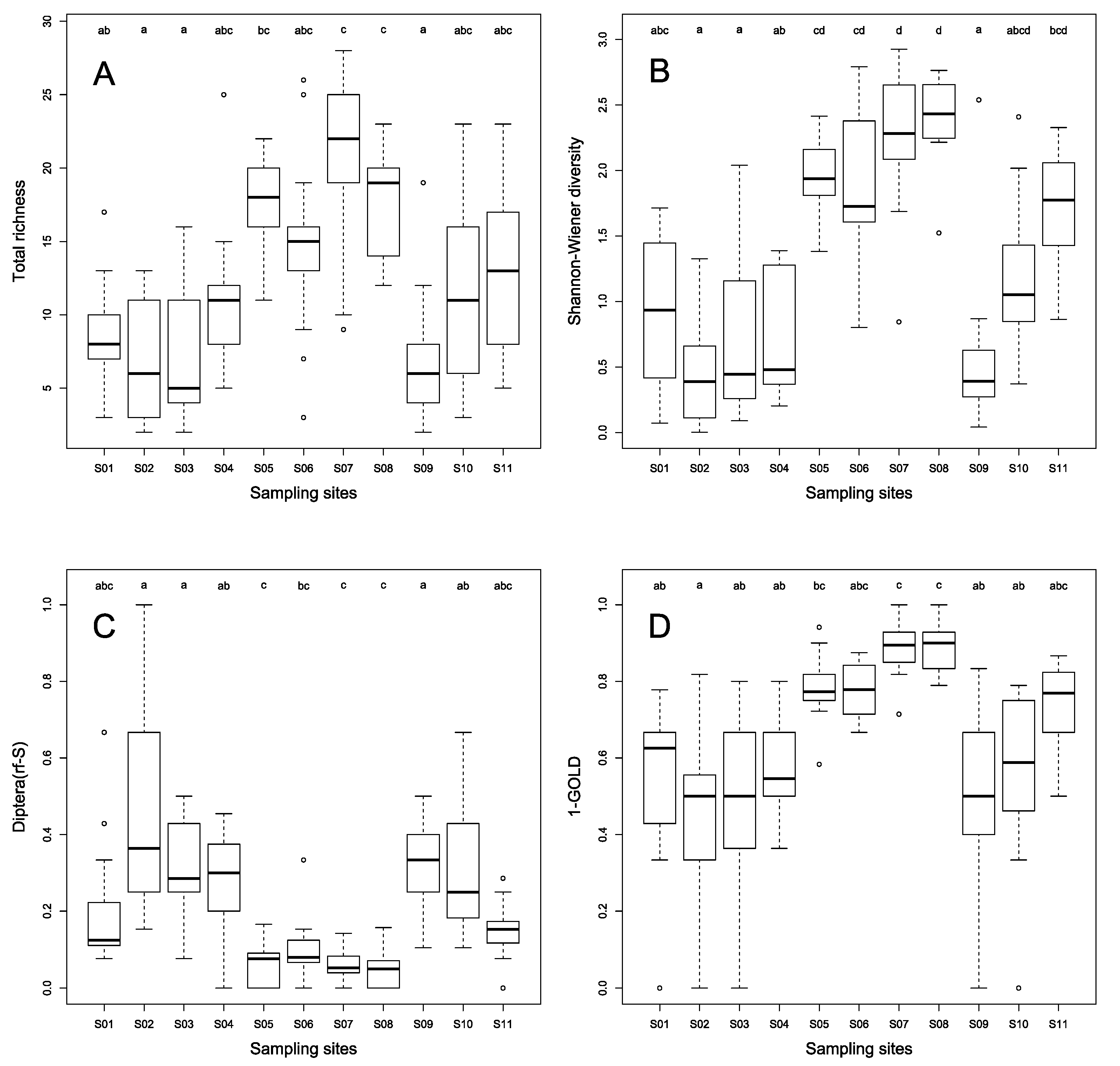
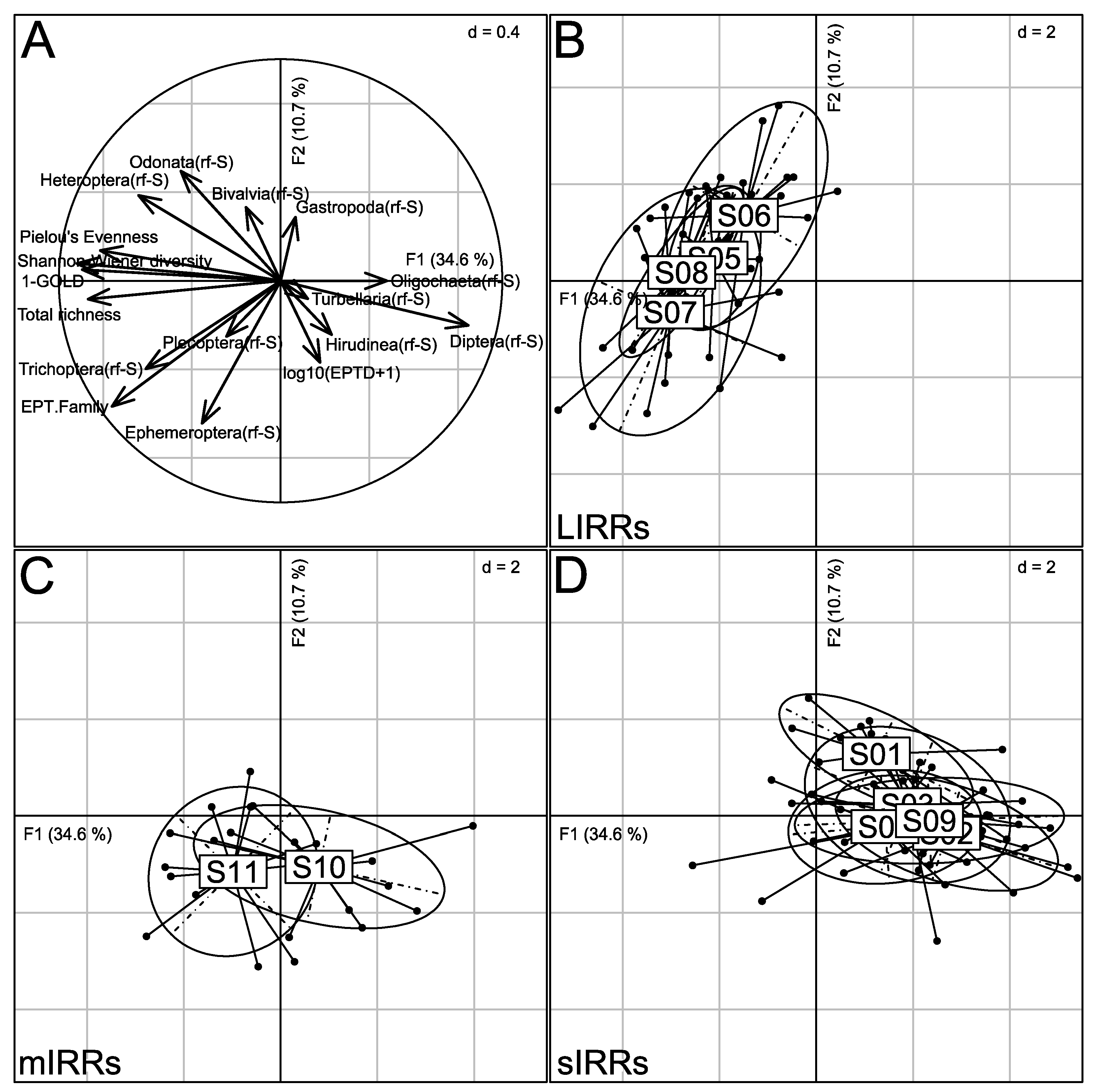
3.4. Taxonomic Structure of Stream Benthic Macroinvertebrate Assemblages
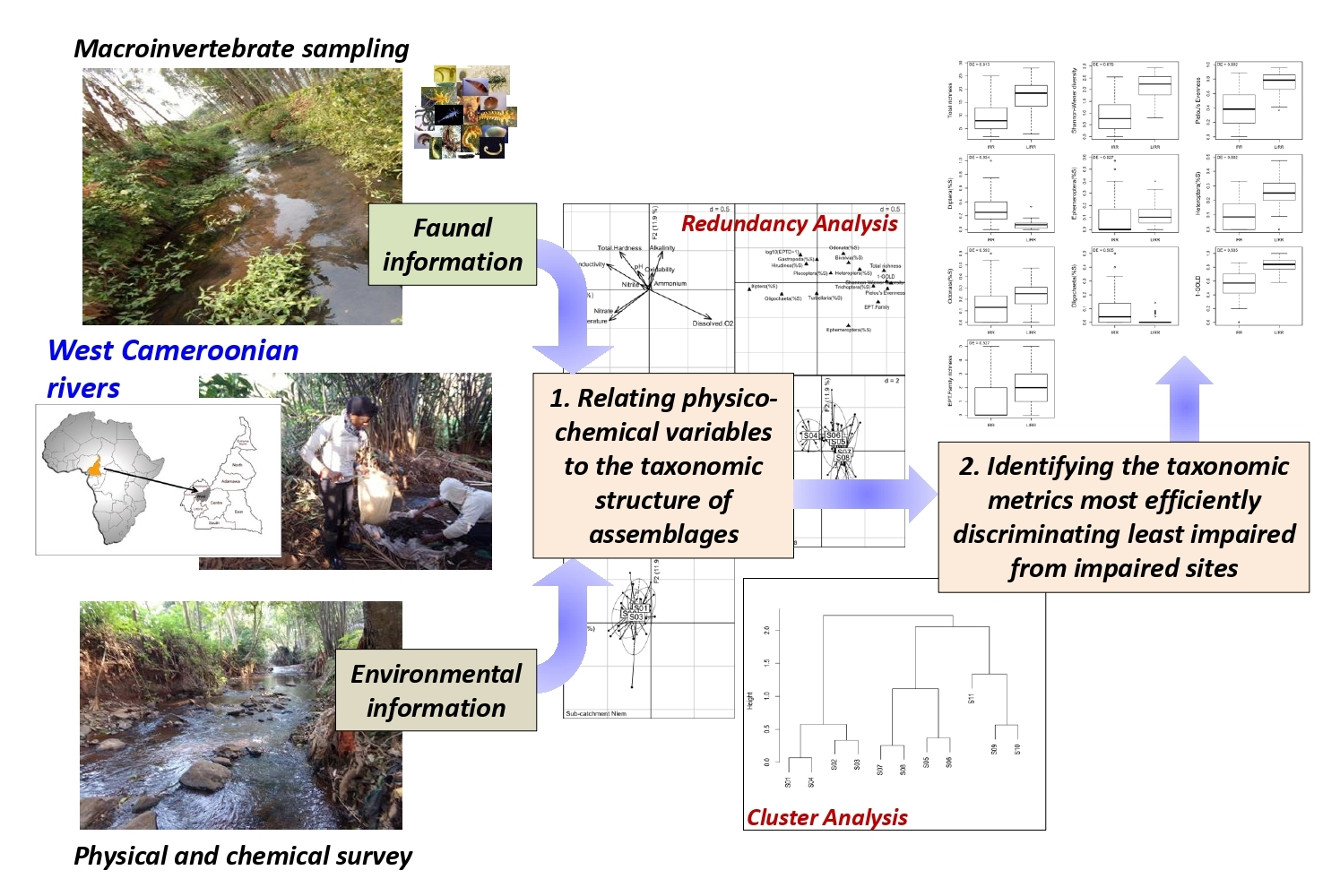
3.5. Relationships between Physico-Chemical Parameters and the Taxonomic Metrics of Benthic Macroinvertebrate Assemblages
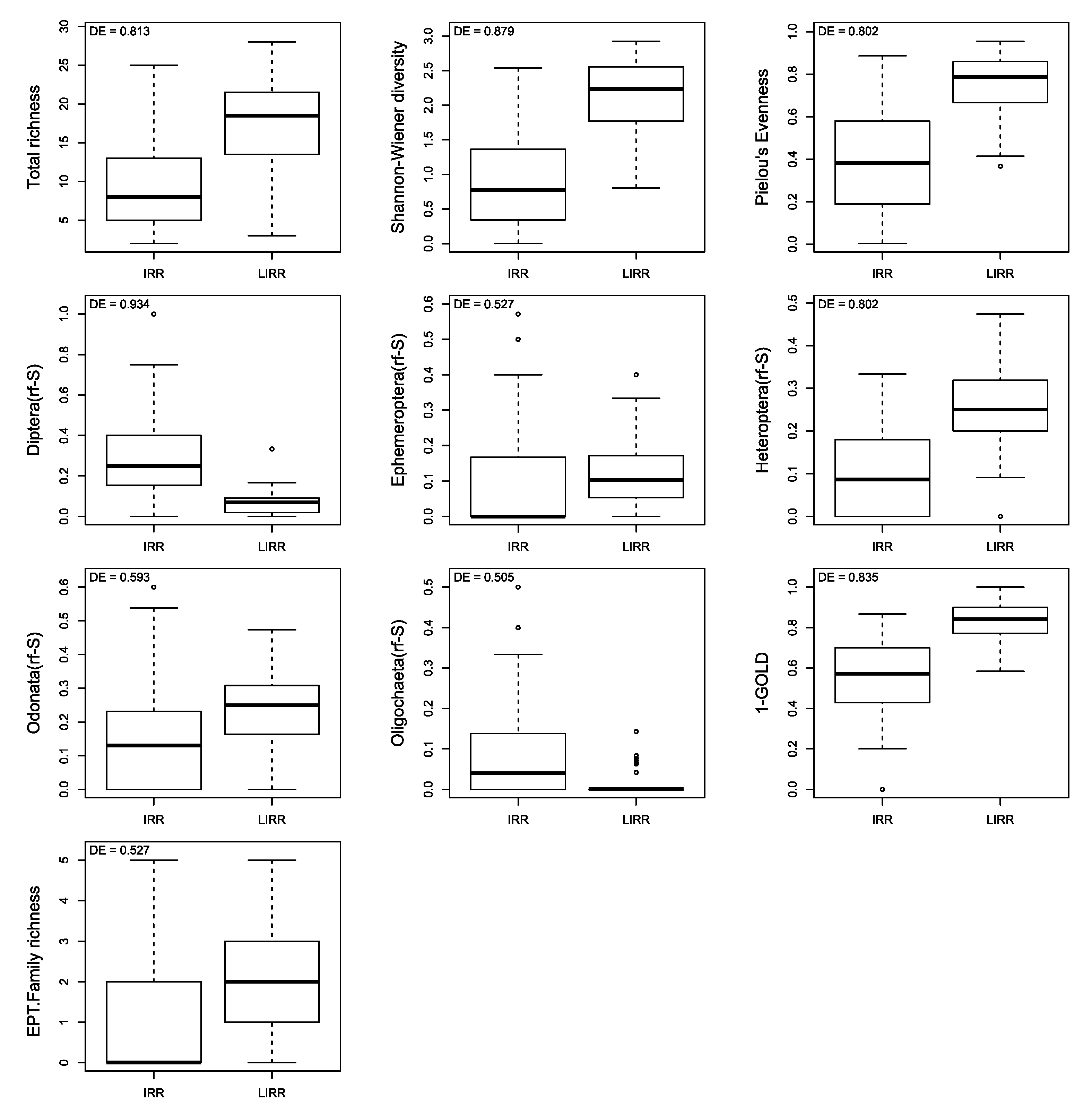
3.6. The Discrimination Efficiency (DE) of the Metrics Describing Benthic Invertebrate Assemblages
4. Discussion
4.1. Physico-Chemical Characteristics of Streams
4.2. Invertebrate-Based Metrics and Their Relationships with Physico-Chemical Parameters
4.3. Metrics of Interest for Future Biomonitoring Tool
4.4. Seasonal Effect on Macroinvertebrate-Based Metrics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Townsend, C.R.; Uhlmann, S.S.; Matthaei, C.D. Individual and combined responses of stream ecosystems to multiple stressors. J. Appl. Ecol. 2008, 45, 1810–1819. [Google Scholar] [CrossRef]
- Ormerod, S.J.; Dobson, M.; Hildrew, A.G.; Townsend, C.R. Multiple stressors in freshwater ecosystems. Freshw. Biol. 2010, 55, 1–4. [Google Scholar] [CrossRef]
- Nõges, P.; Argillier, C.; Borja, A.; Garmendia, J.M.; Hanganu, J.; Kodes, V.; Pletterbauer, F.; Sagouis, A.; Birk, S. Quantified biotic and abiotic responses to multiple stress in freshwater, marine and ground waters. Sci. Total Environ. 2015, 540, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Castro, D.M.P.; de Carvalho, D.R.; dos Santos Pompeu, P.; Moreira, M.Z.; Nardoto, G.B.; Callisto, M. Land use influences niche size and the assimilation of resources by benthic macroinvertebrates in tropical headwater streams. PLoS ONE 2016, 11, e0150527. [Google Scholar]
- Christensen, M.R.; Graham, M.D.; Vinebrooke, R.D.; Findlay, D.L.; Paterson, M.J.; Turner, M.A. Multiple anthropogenic stressors cause ecological surprises in boreal lakes. Glob. Chang. Biol. 2006, 12, 2316–2322. [Google Scholar] [CrossRef]
- Sirombra, M.G.; Mesa, L.M. A method for assessing the ecological quality of riparian forests in subtropical Andean streams: QBRy index. Ecol. Indic. 2012, 20, 324–331. [Google Scholar] [CrossRef]
- Taniwaki, R.H.; Piggott, J.J.; Ferraz, S.F.; Matthaei, C.D. Climate change and multiple stressors in small tropical streams. Hydrobiologia 2017, 793, 41–53. [Google Scholar] [CrossRef]
- Meyer, J.L.; Paul, M.J.; Taulbee, W.K. Stream ecosystem function in urbanizing landscapes. J. N. Am. Benthol. Soc. 2005, 24, 602–612. [Google Scholar] [CrossRef]
- Walsh, C.J.; Roy, A.H.; Feminella, J.W.; Cottingham, P.D.; Groffman, P.M.; Morgan II, R.P. The urban stream syndrome: Current knowledge and the search for a cure. J. N. Am. Benthol. Soc. 2005, 24, 706–723. [Google Scholar] [CrossRef]
- Cuffney, T.F.; Brightbill, R.A.; May, J.T.; Waite, I.R. Responses of benthic macroinvertebrates to environmental changes associated with urbanization in nine metropolitan areas. Ecol. Appl. 2010, 20, 1384–1401. [Google Scholar] [CrossRef]
- Holland-Clift, S.; O’Dowd, D.J.; Mac Nally, R. Impacts of an invasive willow (Salix × rubens) on riparian bird assemblages in south-eastern Australia. Austral Ecol. 2011, 36, 511–520. [Google Scholar] [CrossRef]
- Kouam Kenmogne, G.R.; Rosillon, F.; Nono, A.; Nzeukou Nzeugang, A.; Mpakam, H.G. Les maladies hydriques à l’épreuve de la gestion des ressources en eau dans une zone urbaine d’un pays en développement: Cas de la ville de Yaoundé (Centre-Cameroun). Eur. J. Water Qual. 2011, 42, 35–49. [Google Scholar] [CrossRef]
- Aazami, J.; Esmaili-Sari, A.; Abdoli, A.; Sohrabi, H.; Van den Brink, P.J. Monitoring and assessment of water health quality in the Tajan River, Iran using physicochemical, fish and macroinvertebrates indices. J. Environ. Health Sci. Eng. 2015, 13, 260–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aweng, E.R.; Ismid, M.S.; Maketab, M. The effect of land uses on physicochemical water quality at three rivers in Sungai Endau watershed, Kluang, Johor, Malaysia. Aust. J. Basic Appl. Sci. 2011, 5, 923–932. [Google Scholar]
- Zarei, H.; Pourreza Bilondi, M. Factor analysis of chemical composition in the Karoon River basin, southwest of Iran. Appl. Water Sci. 2013, 3, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Bere, T.; Dalu, T.; Mwedzi, T. Detecting the impact of heavy metal contaminated sediment on benthic macroinvertebrate communities in tropical streams. Sci. Total Environ. 2016, 572, 147–156. [Google Scholar] [CrossRef]
- Unyimadu, J.P.; Osibanjo, O.; Babayemi, J.O. Selected persistent organic pollutants (POPs) in water of River Niger: Occurrence and distribution. Environ. Monit. Assess. 2018, 190, 6. [Google Scholar] [CrossRef]
- Heatherly, T.; Whiles, M.R.; Royer, T.V.; David, M.B. Relationships between water quality, habitat quality, and macroinvertebrate assemblages in Illinois streams. J. Environ. Qual. 2007, 36, 1653–1660. [Google Scholar] [CrossRef] [Green Version]
- Deborde, D.D.D.; Hernandez, M.B.M.; Magbanua, F.S. Benthic macroinvertebrate community as an indicator of stream health: The effects of land use on stream benthic macroinvertebrates. Sci. Diliman 2016, 28, 5–26. [Google Scholar]
- Borisko, J.P.; Kilgour, B.; Stanfield, L.W.; Jones, F.C. An evaluation of rapid bioassessment protocols for stream benthic invertebrates in Southern Ontario, Canada. Water Qual. Res. J. Can. 2007, 42, 184–193. [Google Scholar] [CrossRef]
- Buss, D.F.; Vitorino, A.S. Rapid bioassessment protocols using benthic macroinvertebrates in Brazil: Evaluation of taxonomic sufficiency. J. N. Am. Benthol. Soc. 2010, 29, 562–571. [Google Scholar] [CrossRef]
- Gupta, M.; Paliwal, A. Role of aquatic insects of water quality in related to physico-chemical parameters in Yamuna River at district Firozabad (U.P.). Adv. Biores. 2010, 1, 70–73. [Google Scholar]
- Mackintosh, T.J.; Davis, J.A.; Thompson, R.M. The influence of urbanisation on macroinvertebrate biodiversity in constructed stormwater wetlands. Sci. Total Environ. 2015, 536, 527–537. [Google Scholar] [CrossRef]
- Mermillod-Blondin, F. The functional significance of bioturbation and biodeposition on biogeochemical processes at the water–sediment interface in freshwater and marine ecosystems. J. N. Am. Benthol. Soc. 2011, 30, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Mereta, S.T.; Boets, P.; Bayih, A.A.; Malu, A.; Ephrem, Z.; Sisay, A.; Endale, H.; Yitbarek, M.; Jemal, A.; De Meester, L.; et al. Analysis of environmental factors determining the abundance and diversity of macroinvertebrate taxa in natural wetlands of Southwest Ethiopia. Ecol. Inform. 2012, 7, 52–61. [Google Scholar] [CrossRef]
- Floury, M.; Usseglio-Polatera, P.; Férréol, M.; Delattre, C.; Souchon, Y. Global climate change in large European rivers: Long-term effects on macroinvertebrate communities and potential local confounding factors. Glob. Chang. Biol. 2013, 19, 1085–1099. [Google Scholar] [CrossRef]
- Friberg, N.; Bonada, N.; Bradley, D.C.; Dunbar, M.J.; Edwards, F.K.; Grey, J.; Hayes, R.B.; Hildrew, A.G.; Lamouroux, N.; Trimmer, M.; et al. Biomonitoring of human impacts in freshwater ecosystems: The good, the bad and the ugly. Adv. Ecol. Res. 2011, 44, 1–68. [Google Scholar]
- Colas, F.; Archaimbault, V.; Férard, J.-F.; Bouquerel, J.; Roger, M.-C.; Devin, S. Benthic indicators of sediment quality associated with run-of-river reservoirs. Hydrobiologia 2013, 703, 149–164. [Google Scholar]
- Ferreira, A.; Cyrino, J.E.P.; Duarte-Neto, P.J.; Martinelli, L.A. Permeability of riparian forest strips in agricultural, small subtropical watersheds in south-eastern Brazil. Mar. Freshw. Res. 2013, 63, 1272–1282. [Google Scholar] [CrossRef]
- Dallas, H.F.; Rivers-Moore, N. Ecological consequences of global climate change for freshwater ecosystems in South Africa. S. Afr. J. Sci. 2014, 110, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Wang, Z.; Duan, X.; Pan, B. Effects of pollution on macroinvertebrates and water quality bio-assessment. Hydrobiologia 2014, 729, 247–259. [Google Scholar] [CrossRef]
- Graeber, D.; Jensen, T.M.; Rasmussen, J.J.; Riis, T.; Wiberg-Larsen, P.; Baattrup-Pedersen, A. Multiple stress response of lowland stream benthic macroinvertebrates depends on habitat type. Sci. Total Environ. 2017, 599–600, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- Tafangenyasha, C.; Dzinomwa, T. Land-use impacts on river water quality in lowveld sand river systems in south-east Zimbabwe. Land Use Water Resour. Res. 2005, 5, 3.1–3.10. [Google Scholar]
- Adandedjan, D.; Lalèyè, P.; Ouattara, A.; Gourene, G. Distribution of benthic insect fauna in a West African lagoon: The Porto-Novo Lagoon in Bénin. Asian J. Biol. Sci. 2011, 4, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Kilonzo, F.; Masese, F.O.; Van Griensven, A.; Bauwens, W.; Obando, J.; Lens, P.N.L. Spatial–temporal variability in water quality and macro-invertebrate assemblages in the Upper Mara River basin, Kenya. Phys. Chem. Earth Parts A/B/C 2014, 67–69, 93–104. [Google Scholar] [CrossRef]
- Mutonkole Senga, P.; Tshitenge Mbuebue, J.-M.; Masamba Lulendo, N. Benthic macroinvertebrates as indicators of water quality: A case-study of urban Funa stream (in Kinshasa, Democratic Republic of Congo). Open J. Water Pollut. Treat. 2015, 2, 8–24. [Google Scholar]
- Onana, F.M.; Zébazé Togouet, S.H.; Koji, E.; Nyamsi Tchatcho, N.L.; Tchakonté, S. Influence of municipal and industrial pollution on the diversity and the structure of benthic macro-invertebrates community of an urban river in Douala, Cameroon. J. Biodivers. Environ. Sci. 2016, 8, 120–133. [Google Scholar]
- Koji, E.; Noah Ewoti, O.V.; Onana, F.M.; Tchakonté, S.; Lontsi Djimeli, C.; Tamsa Arfao, A.; Bricheux, G.; Sime-Ngando, T.; Nola, M. Influence of anthropogenic pollution on the abundance dynamics of some freshwater invertebrates in the coastal area of Cameroon. J. Environ. Prot. 2017, 8, 810–829. [Google Scholar] [CrossRef] [Green Version]
- Edia, O.E.; Bony, K.Y.; Konan, K.F.; Ouattara, A.; Gourène, G. Distribution of aquatic insects among four costal river habitats (Côte d’Ivoire, West-Africa). Bull. Env. Pharmacol. Life Sci. 2013, 2, 68–77. [Google Scholar]
- Kaboré, I.; Moog, O.; Alp, M.; Guenda, W.; Koblinger, T.; Mano, K.; Ouéda, A.; Ouédraogo, R.; Trauner, D.; Melcher, A.H. Using macroinvertebrates for ecosystem health assessment in semi-arid streams of Burkina Faso. Hydrobiologia 2016, 766, 57–74. [Google Scholar] [CrossRef]
- Foto Menbohan, S.; Tchakonté, S.; Ajeagah, G.A.; Zébazé Togouet, S.H.; Bilong Bilong, C.F.; Njiné, T. Water quality assessment using benthic macroinvertebrates in a peri-urban stream (Cameroon). Int. J. Biotechnol. 2013, 2, 91–104. [Google Scholar]
- Tchakonté, S.; Ajeagah, G.A.; Camara, A.I.; Diomandé, D.; Nyamsi Tchatcho, N.L.; Ngassam, P. Impact of urbanization on aquatic insect assemblages in the coastal zone of Cameroon: The use of biotraits and indicator taxa to assess environmental pollution. Hydrobiologia 2015, 755, 123–144. [Google Scholar] [CrossRef]
- Heino, J.; Virkkala, R.; Toivonen, H. Climate change and freshwater biodiversity: Detected patterns, future trends and adaptations in northern regions. Biol. Rev. 2009, 84, 39–54. [Google Scholar] [CrossRef]
- Poulton, B.C.; Graham, J.L.; Rasmussen, T.J.; Stone, M.L. Responses of macroinvertebrate community metrics to a wastewater discharge in the Upper Blue River of Kansas and Missouri, USA. J. Water Resour. Prot. 2015, 7, 1195–1220. [Google Scholar] [CrossRef] [Green Version]
- Parent-Raoult, C.; Boisson, J.C. Impacts des rejets urbains de temps de pluie (RUTP) sur les milieux aquatiques: État des connaissances. Rev. Sci. Eau 2007, 20, 229–239. [Google Scholar]
- Olivry, J.C. Fleuves et Rivières du Cameroun; Edition Mesres-Orstom: Paris, France, 1986; 733p. [Google Scholar]
- Abdi, H.; Valentin, D. Multiple Correspondence Analysis. In Encyclopedia of Measurement and Statistics; Salkind, N., Ed.; Sage: Thousand Oaks, CA, USA, 2007. [Google Scholar]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011; 319p. [Google Scholar]
- Stark, J.D.; Boothroyd, K.G.; Harding, J.S.; Maxted, J.R.; Scarsbrook, M.R. Protocols for Sampling Macroinvertebrates in Wadeable Streams. New Zealand Macro-Invertebrates Working Group, Report No.1, Ministry for the Environment and Sustainable Management, Fund Project n° 5103. 2001. Available online: https://riversgroup.org.nz/wp-content/uploads/2018/06/4.1.3-macroinvertebrate-sampling.pdf (accessed on 5 September 2015).
- Mpakam, H.G. Water Resource Vulnerability to Pollution in the Bafoussam Region (West Cameroon) and Socio-Economic and Health Effects: Sanitation Procedures. Ph.D. Thesis, University of Yaoundé I, Yaoundé, Cameroon, 2009. [Google Scholar]
- Durand, J.R.; Lévêque, C. Flore et Faune Aquatiques de l’Afrique Sahélo-Soudanienne. Tome II; Edition de l’ORSTOM: Paris, France, 1981; 517p. [Google Scholar]
- Day, J.A.; Harrison, A.D.; de Moor, I.J. Guides to the freshwater invertebrates of Southern Africa, Volume 9: Diptera. In Water Research Commission Report; No. TT 201/02; Water Research Commission: Pretoria, South Africa, 2002; 210p. [Google Scholar]
- De Moor, I.J.; Day, J.A. Guides to the freshwater invertebrates of Southern Africa, Volume 6: Arachnida & Mollusca. Chapter 3: Mollusca. In Water Research Commission Report; No. TT 182/02; Water Research Commission: Pretoria, South Africa, 2002; pp. 42–125. [Google Scholar]
- De Moor, I.J.; Day, J.A.; De Moor, F.C. Guides to the freshwater invertebrates of Southern Africa, Volume 7: Insecta I. Ephemeroptera, Odonata & Plecoptera. In Water Research Commission Report; No. TT 207/03; Water Research Commission: Pretoria, South Africa, 2003; 301p. [Google Scholar]
- De Moor, I.J.; Day, J.A.; De Moor, F.C. Guides to the freshwater invertebrates of Southern Africa, Volume 8: Insecta II. Hemiptera, Megaloptera, Neuroptera, Trichoptera & Lepidoptera. In Water Research Commission Report; No. TT 214/03; Water Research Commission: Pretoria, South Africa, 2003; 219p. [Google Scholar]
- Tachet, H.; Richoux, P.; Bournaud, M.; Usseglio-Polatera, P. Invertébrés d’Eau Douce. Systématique, Biologie, Écologie; CNRS Éditions: Paris, France, 2010; 600p. [Google Scholar]
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish (Volume 339); US Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999.
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Feld, C.K.; de Bello, F.; Dolédec, S. Biodiversity of traits and species both show weak responses to hydromorphological alteration in lowland river macroinvertebrates. Freshw. Biol. 2014, 59, 233–248. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Buffagni, A.; Erba, S.; Cazzola, M.; Murray-Bligh, J.; Soszka, H.; Genoni, P. The STAR common metrics approach to the WFD intercalibration process: Full application for small, lowland rivers in three European countries. Hydrobiologia 2006, 566, 379–399. [Google Scholar] [CrossRef]
- APHA Standard Method for Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998; 1150p.
- Rodier, J.; Legube, B.; Marlet, N.; Brunet, R. L’Analyse de l’Eau, 9th ed.; Dunod: Paris, France, 2009; 1579p. [Google Scholar]
- Siegel, S.; Castellan, N.J. Non Parametric Statistics for the Behavioral Sciences; McGraw-Hill: New York, NY, USA, 1988; 399p. [Google Scholar]
- Dolédec, S.; Chessel, D. Rythmes saisonniers et composantes stationnelles en milieu aquatique. II. Prise en compte et élimination d’effets dans un tableau faunistique. Acta Oecol. Oecol. Gen. 1989, 10, 207–232. [Google Scholar]
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Dray, S.; Dufour, A.B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Chevenet, F.; Dolédec, S.; Chessel, D. A fuzzy coding approach for the analysis of long-term ecological data. Freshw. Biol. 1994, 31, 295–309. [Google Scholar] [CrossRef]
- Galani, J.H.; Houbraken, M.; Wumbei, A.; Djeugap, J.F.; Fotio, D.; Spanoghe, P. Evaluation of 99 pesticide residues in major agricultural products from the Western Highlands Zone of Cameroon using QuEChERS method extraction and LC-MS/MS and GC-ECD analyses. Foods 2018, 7, 184. [Google Scholar] [CrossRef] [Green Version]
- Oscoz, J.; Galicia, D.; Miranda, R. (Eds.) Identification Guide of Freshwater Macroinvertebrates of Spain; Springer: Dordrecht, The Netherlands, 2011; 45p. [Google Scholar]
- Morrissey, C.A.; Boldt, A.; Mapstone, A.; Newton, J.; Ormerod, S.J. Stable isotopes as indicators of wastewater effects on the macroinvertebrates of urban rivers. Hydrobiologia 2013, 700, 231–244. [Google Scholar] [CrossRef]
- Varandas, S.G.; Cortes, R.M.V. Evaluating macroinvertebrate biological metrics for ecological assessment of streams in northern Portugal. Environ. Monit. Assess. 2010, 166, 201–221. [Google Scholar] [CrossRef]
- Hepp, L.U.; Restello, R.M.; Milesi, S.V.; Biasi, C.; Molozzi, J. Distribution of aquatic insects in urban headwater streams. Acta Limnol. Bras. 2013, 25, 1–9. [Google Scholar] [CrossRef]
- Prommi, T.; Payakka, A. Aquatic insect biodiversity and water quality parameters of streams in Northern Thailand. Sains Malays. 2015, 44, 707–717. [Google Scholar] [CrossRef]
- Qu, X.; Wu, N.; Tang, T.; Cai, Q.; Park, Y.S. Effects of heavy metals on benthic macroinvertebrate communities in high mountain streams. Ann. Limnol. 2010, 46, 291–302. [Google Scholar] [CrossRef]
- Rawson, C.A.; Lim, R.P.; Tremblay, L.A.; Warne, M.S.J.; Ying, G.G.; Laginestra, E.; Chapman, J.C. Benthic macroinvertebrate assemblages in remediated wetlands around Sydney, Australia. Ecotoxicology 2010, 19, 1589–1600. [Google Scholar] [CrossRef] [PubMed]
- Arimoro, F.O.; Muller, W.J. Mayfly (Insecta: Ephemeroptera) community structure as an indicator of the ecological status of a stream in the Niger Delta area of Nigeria. Environ. Monit. Assess. 2010, 166, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Shelly, S.Y.; Mirza, Z.B.; Bashir, S. Comparative ecological study of aquatic macroinvertebrates of Mangla dam and Chashma barrage wetland areas. J. Anim. Plant Sci. 2011, 21, 340–350. [Google Scholar]
- Myers, L.W.; Kondratieff, B.C.; Mihuc, T.B.; Ruiter, D.E. The mayflies (Ephemeroptera), stoneflies (Plecoptera), and caddisflies (Trichoptera) of the Adirondack Park (New York State). Trans. Am. Entomol. Soc. 2011, 137, 63–140. [Google Scholar] [CrossRef]
- Kaboré, I.; Ouédraogo, I.; Tampo, L.; Ouéda, A.; Moog, O.; Guenda, W.; Melcher, A.H. Composition and dynamic of benthic macroinvertebrates community in semi-arid area rivers of Burkina Faso (West Africa). Int. J. Biol. Chem. Sci. 2016, 10, 1542–1561. [Google Scholar] [CrossRef] [Green Version]
- Duka, S.; Pepa, B.; Keci, E.; Paparisto, A.; Lazo, P. Biomonitoring of water quality of the Osumi, Devolli, and Shkumbini rivers through benthic macroinvertebrates and chemical parameters. J. Environ. Sci. Health Part A 2017, 52, 471–478. [Google Scholar] [CrossRef]
- Nyamsi Tchatcho, N.L.; Foto Menbohan, S.; Zébazé Togouet, S.H.; Onana, F.M.; Adandedjan, D.; Tchakonté, S.; Yémélé Tsago, C.; Koji, E.; Njiné, T. Indice multimétrique des macroinvertébrés benthiques yaoundéens (IMMY) pour l’évaluation biologique de la qualité des eaux des cours d’eau de la région du Centre Sud Forestier du Cameroun. Eur. J. Sci. Res. 2014, 123, 412–430. [Google Scholar]
- Cook, S.E.; Fisher, M.J.; Andersson, M.S.; Rubiano, J.; Giordano, M. Water, food and livelihoods in river basins. Water Int. 2009, 34, 13–29. [Google Scholar] [CrossRef]
- Dar, P.A.; Reshi, Z.A. Components, processes and consequences of biotic homogenization: A review. Contemp. Probl. Ecol. 2014, 7, 123–136. [Google Scholar] [CrossRef]
- Doretto, A.; Piano, E.; Bona, F.; Fenoglio, S. How to assess the impact of fine sediments on the macroinvertebrate communities of alpine streams? A selection of the best metrics. Ecol. Indic. 2018, 84, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Mondy, C.P.; Villeneuve, B.; Archaimbault, V.; Usseglio-Polatera, P. A new macroinvertebrate-based multimetric index (I2M2) to evaluate ecological quality of French wadeable streams fulfilling the WFD demands: A taxonomical and trait approach. Ecol. Indic. 2012, 18, 452–467. [Google Scholar] [CrossRef]
- Warwick, R.M.; Clarke, K.R. New ‘biodiversity’ measures reveal a decrease in taxonomic distinctness with increasing stress. Mar. Ecol. Prog. Ser. 1995, 129, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Böhmer, J.; Rawer-Jost, C.; Zenker, A. Multimetric assessment of data provided by water managers from Germany: Assessment of several different types of stressors with macrozoobenthos communities. Hydrobiologia 2004, 516, 215–228. [Google Scholar] [CrossRef]
- Dedieu, N.; Clavier, S.; Vigouroux, R.; Cerdan, P.; Céréghino, R. A multimetric macroinvertebrate index for the implementation of the European Water Framework Directive in French Guiana, East Amazonia. River Res. Appl. 2016, 32, 501–515. [Google Scholar] [CrossRef] [Green Version]
- Stoddard, J.L.; Herlihy, A.T.; Peck, D.V.; Hughes, R.M.; Whittier, T.R.; Tarquinio, E. A process for creating multimetric indices for large-scale aquatic surveys. J. N. Am. Benthol. Soc. 2008, 27, 878–891. [Google Scholar] [CrossRef]
- Pereira, P.S.; Souza, N.F.; Baptista, D.F.; Oliveira, J.L.M.; Buss, D.F. Incorporating natural variability in the bioassessment of stream condition in the Atlantic Forest biome, Brazil. Ecol. Indic. 2016, 69, 606–616. [Google Scholar] [CrossRef]
- Brabec, K.; Zahrádková, S.; Nĕmejcová, D.; Pařil, P.; Kokeš, J.; Jarkovský, J. Assessment of organic pollution effect considering differences between lotic and lentic stream habitats. Hydrobiologia 2004, 516, 331–346. [Google Scholar] [CrossRef]
- Ofenböck, T.; Moog, O.; Gerritsen, J.; Barbour, M. A stressor specific multimetric approach for monitoring running waters in Austria using benthic macroinvertebrates. Hydrobiologia 2004, 516, 251–268. [Google Scholar] [CrossRef]
- Hering, D.; Feld, C.K.; Moog, O.; Ofenböck, T. Cook book for the development of a multimetic index for biological condition of aquatic ecosystems: Experiences from the European AQEM and STAR projects and related initiatives. Hydrobiologia 2006, 566, 311–324. [Google Scholar] [CrossRef]
- Chen, K.; Hughes, R.M.; Xu, S.; Zhang, J.; Cai, D.; Wang, B. Evaluating performance of macroinvertebrate-based adjusted and unadjusted multi-metric indices (MMI) using multi-season and multi-year samples. Ecol. Indic. 2014, 36, 142–151. [Google Scholar] [CrossRef]
- Chessman, B.C.; Thurtell, L.A.; Royal, M.J. Bioassessment in a harsh environment: A comparison of macroinvertebrate assemblages at reference and assessment sites in an Australian inland river system. Environ. Monit. Assess. 2006, 119, 303–330. [Google Scholar] [CrossRef]
- Pond, G.J. Biodiversity loss in Appalachian headwater streams (Kentucky, USA): Plecoptera and Trichoptera communities. Hydrobiologia 2012, 679, 97–117. [Google Scholar] [CrossRef]
- Buffagni, A.; Erba, S.; Cazzola, M.; Kemp, J.L. The AQEM multimetric system for the southern Italian Apennines: Assessing the impact of water quality and habitat degradation on pool macroinvertebrates in Mediterranean rivers. Hydrobiologia 2004, 516, 313–329. [Google Scholar] [CrossRef]
- Dias-Silva, K.; Cabette, H.S.R.; Juen, L.; De Marco, P., Jr. The influence of habitat integrity and physical-chemical water variables on the structure of aquatic and semi-aquatic Heteroptera. Zoologia (Curitiba) 2010, 27, 918–930. [Google Scholar] [CrossRef] [Green Version]
- Vieira, T.B.; Dias-Silva, K.; Pacífico, E.S. Effects of riparian vegetation integrity on fish and Heteroptera communities. Appl. Ecol. Environ. Res. 2014, 13, 53–65. [Google Scholar]
- Verdonschot, R.C.M.; Keizer-Vlek, H.E.; Verdonschot, P.F.M. Development of a multimetric index based on macroinvertebrates for drainage ditch networks in agricultural areas. Ecol. Indic. 2012, 13, 232–242. [Google Scholar] [CrossRef]
- Pinto, P.; Rosado, J.; Morais, M.; Antunes, I. Assessment methodology for southern siliceous basins in Portugal. Hydrobiologia 2004, 516, 191–214. [Google Scholar] [CrossRef]
- Lafont, M.; Camus, J.C.; Rosso, A. Superficial and hyporheic oligochaete communities as indicators of pollution and water exchange in the River Moselle, France. Hydrobiologia 1996, 334, 147–155. [Google Scholar] [CrossRef]
- Prygiel, J.; Rosso-Darmet, A.; Laffont, M.; Lesniak, C.; Durbec, A.; Ouddane, B. Use of oligochaete communities for assessment of ecotoxicological risk in fine sediment of rivers and canals of the Artois-Picardie water basin (France). Hydrobiologia 1999, 410, 25–37. [Google Scholar] [CrossRef]
- Vivien, R.; Tixier, G.; Lafont, M. Use of oligochaete communities for assessing the quality of sediments in watercourses of the Geneva area (Switzerland) and Artois-Picardie basin (France): Proposition of heavy metal toxicity thresholds. Ecohydrol. Hydrobiol. 2014, 14, 142–151. [Google Scholar] [CrossRef]
- Failla, A.J.; Vasquez, A.A.; Fujimoto, M.; Ram, J.L. The ecological, economic and public health impacts of nuisance chironomids and their potential as aquatic invaders. Aquat. Invasions 2015, 10, 1–15. [Google Scholar] [CrossRef]
- Machado, N.G.; Nassarden, D.C.S.; dos Santos, F.; Boaventura, I.C.G.; Perrier, G.; de Souza, F.S.C.; de Lima Martins, E.; Biudes, M.S. Chironomus larvae (Chironomidae: Diptera) as water quality indicators along an environmental gradient in a neotropical urban stream. Rev. Ambient. Agua 2015, 10, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Odume, O.N.; Muller, W.J. Diversity and structure of Chironomidae communities in relation to water quality differences in the Swartkops River. Phys. Chem. Earth Parts A/B/C 2011, 36, 929–938. [Google Scholar] [CrossRef]
- Masese, F.O.; Kitaka, N.; Kipkemboi, J.; Gettel, G.M.; Irvine, K.; McClain, M.E. Macroinvertebrate functional feeding groups in Kenyan highland streams: Evidence for a diverse shredder guild. Freshw. Sci. 2014, 33, 435–450. [Google Scholar] [CrossRef] [Green Version]
- Kalkman, V.J.; Clausnitzer, V.; Dijkstra, K.D.B.; Orr, A.G.; Paulson, D.R.; van Tol, J. Global diversity of dragonflies (Odonata) in freshwater. Hydrobiologia 2008, 595, 351–363. [Google Scholar] [CrossRef]
- Samways, M.J.; Steytler, N.S. Dragonfly (Odonata) distribution patterns in urban and forest landscapes, and recommendations for riparian management. Biol. Conserv. 1996, 78, 279–288. [Google Scholar] [CrossRef]
- Miguel, T.B.; Oliveira-Junior, J.M.B.; Ligeiro, R.; Juen, L. Odonata (Insecta) as a tool for the biomonitoring of environmental quality. Ecol. Indic. 2017, 81, 555–566. [Google Scholar] [CrossRef]
- De Oliveira Junior, J.M.B.; Shimano, Y.; Gardner, T.A.; Hughes, R.M.; Marco Júnior, P.; Juen, L. Neotropical dragonflies (Insecta: Odonata) as indicators of ecological condition of small streams in the eastern Amazon. Austral Ecol. 2015, 40, 733–744. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Bogan, M.T.; Bonada, N.; Rios-Touma, B.; Lytle, D.A. Seasonality and predictability shape temporal species diversity. Ecology 2017, 98, 1201–1216. [Google Scholar] [CrossRef] [Green Version]
- Dalu, T.; Wasserman, R.J.; Tonkin, J.D.; Mwedzi, T.; Magoro, M.L.; Weyl, O.L.F. Water or sediment? Partitioning the role of water column and sediment chemistry as drivers of macroinvertebrate communities in an austral South African stream. Sci. Total Environ. 2017, 607–608, 317–325. [Google Scholar] [CrossRef]
- Callisto, M.; Goulart, M.; Medeiros, A.O.; Moreno, P.; Rosa, C.A. Diversity assessment of benthic macroinvertebrates, yeasts, and microbiological indicators along a longitudinal gradient in Serra Do Cipó, Brazil. Braz. J. Biol. 2004, 64, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Tonkin, J.D.; Arimoro, F.O.; Haase, P. Exploring stream communities in a tropical biodiversity hotspot: Biodiversity, regional occupancy, niche characteristics and environmental correlates. Biodivers. Conserv. 2016, 25, 975–993. [Google Scholar] [CrossRef]
- Dalu, T.; Wasserman, R.J.; Tonkin, J.D.; Alexander, M.E.; Dalu, M.T.B.; Motitsoe, S.N.; Manungo, K.I.; Bepe, O.; Dube, T. Assessing drivers of benthic macroinvertebrate community structure in African highlands: An exploration using multivariate analysis. Sci. Total Environ. 2017, 601–602, 1340–1348. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Catchment Water Courses | Sampling Sites | GPS Coordinates (Latitude/Longitude) | Population Density | Anthropogenic Activities in the Watershed and/or Directly in the Stream | Nature of the Stream Bed | Expected Ecological Status |
|---|---|---|---|---|---|---|
| Nlem | S01 | 05°28′10″85 N 10°24′47″91 E | high | food crops, agricultural cooperative of planters, waste deposits (d, m, i) and cattle farming | waste and mud | sIRR |
| S02 | 05°27′58″16 N 10°24′13″80 E | high | soap factory, brewery of Cameroon, waste deposits (d, m, i) | boulders, sand and waste | sIRR | |
| S03 | 05°27′17″08 N 10°24′08″08 E | high | construction of a retaining wall, food crop and society of coffee company | mud and sand | sIRR | |
| Kena | S04 | 05°30′38″03 N 10°23′10″60 E | low | rock extraction, vegetable farming, waste deposits (d, a) | gravel and sand | sIRR |
| S05 | 05°30′43″85 N 10°22′16″62 E | low | food crops | root mats and sand | LIRR | |
| S06 | 05°31′33″01 N 10°21′52″79 E | low | swampy area | mud and sand | LIRR | |
| S07 | 05°31′33″69 N 10°23′08″77 E | low | food crops | sand | LIRR | |
| S08 | 05°30′54″46 N 10°24′23″54 E | low | food crops | sand | LIRR | |
| Famlem | S09 | 05°28′43″38 N 10°26′40″62 E | high | waste deposits (d, m) and food crops | waste deposits and mud | sIRR |
| S10 | 05°29′20″31 N 10°28′00″46 N | low | sand mining activities and food crops | sand | mIRR | |
| S11 | 05°30′40″15 N 10°28′43″38 E | low | sand mining activities | sand and boulders | mIRR |
| Metric Name | Short Description | Reference | Predicted Variation |
|---|---|---|---|
| Total richness | The number of taxa within an assemblage | Barbour et al., 1999 [57] | decrease |
| Shannon-Wiener diversity (H) | H accounts for taxonomic richness and relative taxon abundance distribution | Shannon, 1948 [58] | decrease |
| Pielou evenness (J) | J = H/Hmax (Feld et al., 2014, [59]) | Pielou, 1966 [60] | decrease |
| Bivalvia(rf-S) | Relative richness of this taxonomic group in the invertebrate assemblage (relative frequency) | increase | |
| Coleoptera(rf-S) | idem | decrease | |
| Dictyoptera(rf-S) | idem | increase | |
| Diptera(rf-S) | idem | increase | |
| Ephemeroptera(rf-S) | idem | [57] | decrease |
| Gastropoda(rf-S) | idem | increase | |
| Heteroptera(rf-S) | idem | decrease | |
| Hirudinea(rf-S) | idem | increase | |
| Odonata(rf-S) | idem | decrease | |
| Oligochaeta(rf-S) | idem | increase | |
| Plecoptera(rf-S) | idem | [57] | decrease |
| Trichoptera(rf-S) | idem | [57] | decrease |
| Turbellaria(rf-S) | idem | decrease | |
| EPT Family richness | Number of Ephemeroptera, Plecoptera and Trichoptera families | [57] | decrease |
| log10(EPTD + 1) | Log-transformed sum of the abundances of a selection of insect families: Heptageniidae, Ephemeridae, Leptophlebiidae (E), Brachycentridae, Goeridae, Polycentropodidae, Limnephilidae, Odontoceridae (T), Dolichopodidae, Stratiomyidae, Dixidae, Empididae, Athericidae (D) and Nemouridae (P) | Buffagni et al., 2006 [61] | decrease |
| 1-GOLD | 1—relative abundance of Gastropoda, Oligochaeta and Diptera | [61] | decrease |
| Nlem Stream | Kena Stream | Famlem Stream | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sampling Sites | S01 | S02 | S03 | S04 | S05 | S06 | S07 | S08 | S09 | S10 | S11 |
| Physico-chemical parameters | |||||||||||
| T (°C) *** | 22.8 ± 1.9 | 21.9 ± 2.3 | 22.3 ±1.8 | 20.8 ±1.9 | 19.6 ±1.5 | 20.1 ±1.5 | 21.3 ±1.2 | 21.0 ±2.2 | 22.7 ±0.8 | 22.8 ± 1.3 | 21.9 ± 1.2 |
| pH *** | 7.0 ± 0.5 | 8.0 ± 1.7 | 7.9 ± 1.4 | 7.3 ± 0.6 | 7.0 ± 0.6 | 6.9 ± 0.6 | 7.4 ± 0.4 | 7.3 ± 0.3 | 6.5 ± 0.6 | 7.0 ± 0.6 | 7.6 ± 0.5 |
| Dissolved O2 (%) *** | 41.9 ± 10.2 | 38.8 ± 9.9 | 39.4 ± 11.6 | 42.2 ± 8.7 | 58.0 ± 8.7 | 53.6 ± 6.9 | 64.0 ± 7.8 | 57.0 ± 8.8 | 40.7 ± 8.4 | 55.6 ± 9.2 | 57.1 ± 6.9 |
| Conductivity (μS/cm) *** | 342.3 ± 75.7 | 358.3 ± 356.3 | 299.7 ± 168.9 | 281.9 ± 41.7 | 222.8 ± 45.6 | 164.8 ± 31.1 | 180.8 ± 40.7 | 176.9 ± 50.9 | 272.4 ± 43.6 | 225.0 ± 55.4 | 179.2 ± 54.7 |
| Total hardness (mg/L) *** | 76.1 ± 16.6 | 57.0 ± 19.2 | 56.4 ± 33.4 | 58.1 ± 17.7 | 51.4 ± 14.4 | 34.0 ± 9.9 | 42.5 ± 16.9 | 34.9 ± 15.2 | 47.9 ± 16.0 | 46.6 ± 9.8 | 40.4 ± 11.9 |
| Color (mg/L) | 100.8 ± 62.1 | 210.7 ± 394.7 | 167.5 ± 232.1 | 74.8 ± 85.9 | 102.4 ± 83.4 | 60.5 ± 41.0 | 84.1 ± 93.7 | 108.1 ± 131.4 | 92.5 ± 113.7 | 138.1 ± 142.5 | 190.0 ± 195.3 |
| Dissolved CO2 (mg/L) | 10.7 ± 7.8 | 9.3 ± 9.6 | 10.0 ± 8.3 | 10.0 ± 8.0 | 11.2 ± 7.6 | 10.4 ± 7.6 | 10.0 ± 7.3 | 9.9 ± 6.4 | 11.5 ± 6.2 | 8.7 ± 5.6 | 8.7 ± 5.7 |
| Alkalinity (mg/L) * | 32.2 ± 38.3 | 34.8 ± 43.5 | 30.5 ± 37.2 | 32.8 ± 42.8 | 30.2 ± 37.4 | 28.9 ± 37.5 | 24.3 ± 34.3 | 25.4 ± 33.6 | 20.2 ± 23.6 | 22.3 ± 27.7 | 23.1 ± 30.8 |
| Suspended matter (mg/L) | 24.2 ± 34.7 | 47.8 ± 44.2 | 41.3 ± 61.5 | 28.4 ± 36.6 | 34.3 ± 45.8 | 30.7 ± 46.7 | 39.3 ± 60.6 | 29.5 ± 44.2 | 30.4 ± 64.7 | 75.4 ± 103.0 | 51.1 ± 68.4 |
| Turbidity (mg/L) | 28.1 ± 26.0 | 60.3 ± 113.3 | 32.0 ± 26.9 | 26.3 ± 23.9 | 30.2 ± 38.3 | 27.5 ± 23.4 | 38.7 ± 53.5 | 41.6 ± 66.9 | 26.4 ± 37.2 | 39.2 ± 50.6 | 57.4 ± 71.0 |
| Nitrate (mg/L NO3−) ** | 2.3 ± 1.2 | 5.2 ± 8.4 | 3.2 ± 3.3 | 2.3 ± 1.2 | 1.8 ± 1.3 | 1.5 ± 1.1 | 2.5 ± 1.9 | 1.7 ± 0.9 | 3.0 ± 1.9 | 3.0 ± 1.6 | 3.0 ± 2.3 |
| Orthophophate (mg/L PO43−) | 1.9 ± 2.2 | 2.7 ± 3.6 | 2.5 ± 4.3 | 0.8 ± 1.0 | 1.1 ±1.3 | 0.5 ± 0.8 | 0.5 ± 0.5 | 0.7 ± 1.2 | 0.6 ± 1.0 | 1.6 ± 2.0 | 1.0 ± 1.4 |
| Nitrite (mg/L NO2−) ** | 1.2 ± 1.5 | 4.5 ± 7.7 | 5.7 ± 8.8 | 1.9 ± 2.5 | 1.6 ± 2.2 | 2.5 ± 4.3 | 1.2 ± 2.3 | 1.3 ± 3.3 | 1.0 ± 1.3 | 3.8 ± 4.9 | 3.0 ± 4.7 |
| Ammonium (mg/L NH4+) *** | 1.3 ± 1.4 | 0.7 ± 0.7 | 1.0 ± 1.2 | 1.3 ± 1.0 | 1.0 ± 0.8 | 0.6 ± 1.0 | 0.5 ± 0.4 | 0.5 ± 0.6 | 0.4 ± 0.4 | 1.1 ± 1.4 | 1.0 ± 1.3 |
| BOD5 (mg/L O2) | 39.2 ± 33.8 | 40.4 ± 27.8 | 32.4 ± 24.3 | 44.6 ± 32.5 | 51.9 ± 30.09 | 48.3 ± 23.0 | 26.2 ± 19.5 | 35.4 ± 23.9 | 53.8 ± 40.6 | 38.1 ± 21.4 | 57.3 ± 67.2 |
| Chlorophyll a (µg/L) | 0.2 ± 0.2 | 0.2 ± 0.2 | 0.2 ± 0.3 | 0.1 ± 0.3 | 0.2 ± 0.3 | 0.1 ± 0.2 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.1 ± 0.2 | 0.2 ± 0.2 | 0.1 ± 0.2 |
| Oxidizability (mg/L) * | 4.6 ± 3.6 | 5.1 ± 3.6 | 6.4 ± 6.0 | 6.5 ± 3.1 | 5.4 ± 3.7 | 4.6 ± 2.9 | 4.3 ± 2.7 | 4.7 ± 2.6 | 3.4 ± 1.9 | 4.0 ± 4.1 | 3.5 ± 2.5 |
| Nlem Stream | Kena Stream | Famlem Stream | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sampling Sites | S01 | S02 | S03 | S04 | S05 | S06 | S07 | S08 | S09 | S10 | S11 |
| Metrics | |||||||||||
| Total richness *** | 8.62 ± 3.75 | 6.69 ± 4.01 | 7.38 ± 4.75 | 11.31 ± 5.07 | 17.46 ± 3.71 | 14.69 ± 6.42 | 20.69 ± 6.36 | 17.38 ± 3.84 | 6.69 ± 4.55 | 11.23 ± 6.26 | 13.31 ± 5.41 |
| Shannon diversity *** | 0.92 ± 0.60 | 0.48 ± 0.44 | 0.69 ± 0.62 | 0.71 ± 0.47 | 1.94 ± 0.29 | 1.94 ± 0.61 | 2.27 ± 0.56 | 2.41 ± 0.33 | 0.58 ± 0.63 | 1.23 ± 0.59 | 1.72 ± 0.42 |
| Evenness (Pielou) *** | 0.42 ± 0.25 | 0.23 ± 0.17 | 0.35 ± 0.26 | 0.29 ± 0.16 | 0.68 ± 0.08 | 0.76 ± 0.15 | 0.76 ± 0.15 | 0.85 ± 0.08 | 0.30 ± 0.20 | 0.54 ± 0.17 | 0.69 ± 0.12 |
| Bivalvia(rf-S) * | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.02 | 0.01 ± 0.03 | 0.00 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Coleoptera(rf-S) | 0.13 ± 0.11 | 0.10 ± 0.16 | 0.09 ± 0.10 | 0.07 ± 0.06 | 0.07 ± 0.06 | 0.18 ± 0.12 | 0.14 ± 0.08 | 0.15 ± 0.09 | 0.19 ± 0.18 | 0.10 ± 0.12 | 0.10 ± 0.07 |
| Dictyoptera(rf-S) | 0.01 ± 0.03 | 0.00 ± 0.00 | 0.02 ± 0.03 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.01 | 0.00 ± 0.01 | 0.00 ± 0.01 | 0.00 ± 0.02 | 0.00 ± 0.02 |
| Diptera(rf-S) *** | 0.21 ± 0.17 | 0.46 ± 0.26 | 0.32 ± 0.14 | 0.28 ± 0.12 | 0.06 ± 0.06 | 0.11 ± 0.08 | 0.06 ± 0.04 | 0.05 ± 0.05 | 0.34 ± 0.12 | 0.31 ± 0.17 | 0.15 ± 0.07 |
| Ephemeroptera(rf-S) *** | 0.01 ± 0.04 | 0.03 ± 0.06 | 0.06 ± 0.14 | 0.07 ± 0.08 | 0.11 ± 0.05 | 0.05 ± 0.08 | 0.16 ± 0.10 | 0.14 ± 0.11 | 0.04 ± 0.07 | 0.20 ± 0.11 | 0.25 ± 0.16 |
| Gastropoda(rf-S) *** | 0.17 ± 0.17 | 0.03 ± 0.05 | 0.12 ± 0.11 | 0.12 ± 0.05 | 0.13 ± 0.05 | 0.09 ± 0.06 | 0.04 ± 0.04 | 0.06 ± 0.05 | 0.03 ± 0.07 | 0.06 ± 0.06 | 0.06 ± 0.08 |
| Heteroptera(rf-S) *** | 0.14 ± 0.12 | 0.05 ± 0.08 | 0.10 ± 0.11 | 0.14 ± 0.09 | 0.24 ± 0.08 | 0.23 ± 0.12 | 0.26 ± 0.08 | 0.29 ± 0.09 | 0.09 ± 0.12 | 0.05 ± 0.08 | 0.12 ± 0.10 |
| Hirudinea(rf-S) *** | 0.01 ± 0.03 | 0.08 ± 0.14 | 0.06 ± 0.07 | 0.13 ± 0.07 | 0.03 ± 0.03 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.03 | 0.01 ± 0.03 | 0.06 ± 0.07 | 0.04 ± 0.06 |
| Odonata(rf-S) *** | 0.23 ± 0.15 | 0.18 ± 0.14 | 0.16 ± 0.18 | 0.15 ± 0.09 | 0.27 ± 0.09 | 0.26 ± 0.10 | 0.23 ± 0.13 | 0.23 ± 0.08 | 0.07 ± 0.11 | 0.10 ± 0.10 | 0.13 ± 0.11 |
| Oligochaeta(rf-S) ** | 0.08 ± 0.09 | 0.07 ± 0.09 | 0.07 ± 0.12 | 0.03 ± 0.05 | 0.01 ± 0.03 | 0.02 ± 0.04 | 0.01 ± 0.02 | 0.01 ± 0.03 | 0.15 ± 0.15 | 0.07 ± 0.11 | 0.06 ± 0.05 |
| Plecoptera(rf-S) * | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Trichoptera(rf-S) *** | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.05 ± 0.05 | 0.03 ± 0.05 | 0.05 ± 0.05 | 0.04 ± 0.04 | 0.00 ± 0.01 | 0.00 ± 0.00 | 0.03 ± 0.04 |
| Turbellaria(rf-S) *** | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.04 | 0.00 ± 0.02 | 0.00 ± 0.01 | 0.02 ± 0.03 | 0.01 ± 0.03 | 0.07 ± 0.10 | 0.00 ± 0.01 | 0.05 ± 0.06 |
| EPT Family *** | 0.08 ± 0.28 | 0.31 ± 0.63 | 0.38 ± 0.65 | 0.92 ± 1.19 | 2.31 ± 0.85 | 1.00 ± 1.29 | 3.08 ± 1.44 | 2.62 ± 1.71 | 0.46 ± 1.13 | 1.69 ± 0.95 | 2.62 ± 1.12 |
| log([EPTD + 1]) *** | 0.02 ± 0.08 | 0.25 ± 0.24 | 0.11 ± 0.23 | 0.18 ± 0.25 | 0.00 ± 0.00 | 0.02 ± 0.08 | 0.13 ± 0.21 | 0.00 ± 0.00 | 0.04 ± 0.13 | 0.11 ± 0.23 | 0.02 ± 0.08 |
| 1–GOLD *** | 0.54 ± 0.21 | 0.44 ± 0.24 | 0.49 ± 0.24 | 0.56 ± 0.14 | 0.79 ± 0.09 | 0.78 ± 0.07 | 0.89 ± 0.08 | 0.89 ± 0.06 | 0.48 ± 0.23 | 0.56 ± 0.23 | 0.74 ± 0.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kengne Fotsing, J.; Foto Menbohan, S.; Meyer, A.; Leprêtre, A.; Usseglio-Polatera, P. Relationships between Physico-Chemical Parameters and Taxonomic Structure of Benthic Macroinvertebrate Assemblages in Streams of West Cameroon. Water 2022, 14, 1490. https://doi.org/10.3390/w14091490
Kengne Fotsing J, Foto Menbohan S, Meyer A, Leprêtre A, Usseglio-Polatera P. Relationships between Physico-Chemical Parameters and Taxonomic Structure of Benthic Macroinvertebrate Assemblages in Streams of West Cameroon. Water. 2022; 14(9):1490. https://doi.org/10.3390/w14091490
Chicago/Turabian StyleKengne Fotsing, Joséphine, Samuel Foto Menbohan, Albin Meyer, Alain Leprêtre, and Philippe Usseglio-Polatera. 2022. "Relationships between Physico-Chemical Parameters and Taxonomic Structure of Benthic Macroinvertebrate Assemblages in Streams of West Cameroon" Water 14, no. 9: 1490. https://doi.org/10.3390/w14091490
APA StyleKengne Fotsing, J., Foto Menbohan, S., Meyer, A., Leprêtre, A., & Usseglio-Polatera, P. (2022). Relationships between Physico-Chemical Parameters and Taxonomic Structure of Benthic Macroinvertebrate Assemblages in Streams of West Cameroon. Water, 14(9), 1490. https://doi.org/10.3390/w14091490