1. Introduction
Selenium contamination in soil and water is a growing environmental concern due to selenium’s potential toxicity to humans and wildlife. Selenium is a trace element essential for the proper functioning of many living organisms, including humans. However, excess selenium (greater than 400 µg/d) can be toxic to humans and wildlife [
1]. The effects of selenium toxicity from long-term exposure include fingernail loss, numbness of fingers or toes, and circulatory problems [
2]. This has led health and environmental organizations to set maximum selenium concentrations in drinking water (WHO—40 μg/L, USEPA—50 μg/L) and natural water (USEPA—5 μg/L). Selenium exhibits several oxidation states, including −II, 0, +IV, and +VI. The predominant forms of selenium in water are the oxyanions selenite and selenate, corresponding to the oxidation states +IV and +VI, respectively. In contrast, elemental selenium, with an oxidation number of zero, is highly insoluble and not very toxic [
3].
High selenium concentrations can originate naturally from weathering of seleniferous rocks, whereas anthropogenic selenium contamination is caused mainly by industries, including mining, coal combustion, oil refineries, semiconductors, photoelectric devices, and agriculture [
4,
5]. In mining wastewater, selenium could potentially be found in concentrations ranging from 3–12,000 μg/L [
6]. Besides selenium oxyanions, these waters contain nitrates, especially in mining water; nitrates can be present as residuals from blasting and in agricultural water because of fertilizers [
7,
8]. The availability of nitrate in selenium-contaminated water is a threat, as nitrate anions compete with selenium oxyanions.
Because of toxicity concerns, selenium removal from water using different techniques has attracted considerable attention over the last few decades. Based on the removal mechanism involved, current treatment techniques can be categorized as physical, chemical, and biological. However, physicochemical processes such as chemical precipitation, adsorption, and ion exchange can primarily remove selenite while leaving the more toxic selenate in the effluent. Moreover, many physicochemical methods are not economical [
9]. Reduction methods are more attractive because they treat both selenate and selenite anions with higher removal efficiency. Biological reduction could be considered an economical variation of chemical reduction, because both methods follow similar key reactions to convert selenium oxyanions to elemental selenium. Bioremediation using microorganisms has shown promise as a sustainable and effective approach to removing selenium from the environment [
10]. However, the success of microbial bioremediation can be influenced by various factors, including the presence of competing electron acceptors and donors [
11].
Biological selenium reduction can only occur in an anoxic environment in the presence of a carbon source as an electron donor. The electron donor supplies electrons to the redox reaction, while selenium oxyanions act as electron acceptors and are reduced to elemental selenium. The presence of nitrate affects this reduction reaction because nitrate can also accept electrons. The competition from nitrate is a concern because high levels of nitrates often occur in selenium-contaminated water [
7]. Therefore, biological selenium reduction research in the presence of nitrate anions is important. Selenium oxyanion removal through biological reduction in the presence of nitrate ions has been studied via laboratory treatability experiments, such as batch and kinetic experiments. The microbial communities identified as selenium and nitrate reducers based on previous studies are summarized in
Table 1. Experiments conducted by Lai et al. [
7] with H
2-based membrane biofilm reactors showed that selenate is reduced from 1 mg-Se/L to 0.4 mg-Se/L in the presence of 3 mg-N/L nitrate. Hydrogen gas was the electron donor for the respiratory reduction reaction. The beta proteobacteria phylum has been identified as the most dominant phylum with more than 90% relative abundance among the biofilm microbial community [
7]. Another study conducted on selenate and nitrate reduction in sequencing batch reactor shows that both anions were reduced to below detection level within six hours. The initial selenate and nitrate concentrations were 0.1 mM and 0.96 mM NO
3−—N, respectively. For that study, acetate was the electron donor, and seed sludge was activated sludge from a wastewater treatment plant [
12]. However, most biological selenium reduction experiments have been limited to either pure culture or anaerobic sludge blanket methods. A relatively limited number of studies are available with a mixed microbial community and competitive anions [
7,
12,
13].
To further enhance removal efficiency, the current study focuses on developing and applying simultaneous adsorption and biological reduction treatment processes, with motivation stemming from economic and environmental considerations. A biofiltration system with modified zeolite as the filter medium was investigated for its performance in selenium removal when nitrate is present. Due to their abundance, cost-effectiveness, high porosity, thermal stability, and high surface area, zeolites have been used in recent decades in water treatment. Zeolites offer many advantages in adsorption and attached growth processes because of their high surface area. However, although zeolites can have very high cation exchange capacities, their anion exchange capacity is very limited.
Table 1.
Selenium and nitrate-reducing bacteria reported in the literature.
Table 1.
Selenium and nitrate-reducing bacteria reported in the literature.
| Organism | Taxonomy Level | Species Reduce | Reference |
|---|
| Chloroflexi | Phylum | Nitrate | [14,15] |
| Sulfurospirillum | Genus | Nitrate, selenate | [13,16] |
| Clostridium | Genus | Nitrate | [17] |
| Pseudomonas | Genus | Selenite, selenate | [18] |
| Dechloromonas | Genus | Nitrate, selenate | [19] |
| Methyloversatilis | Genus | Selenate, selenite | [7] |
| Aeromonas hydrophilla | Genus | Selenate, nitrate | [20] |
| Pseudomonas stutzeri pn1 | Genus | Selenate, nitrate | [20] |
| Geobacter sulfurreducens PCA | Genus | Selenite | [20] |
| Veillonella atypica | Genus | Selenite | [20] |
| Seleniivibrio woodruffii S4 | Genus | Selenate | [20] |
| Azospirillum | Genus | Nitrate, selenite | [21,22] |
| Exiguobacterium | Genus | Selenate, nitrate | [23] |
| Synergistia | Genus | Selenate | [24] |
| Lentimicrobium | Genus | Nitrate | [25] |
Thus, different surface modifications have been used to increase zeolite capacity for anions, such as selenite and selenate [
26]. In this study, iron-modified zeolite was used for both oxyanion adsorption and bacterial attachment.
The objective of this study was to investigate selenium removal efficiency in the presence of nitrate anions in chemically modified zeolite columns, as well as to elucidate the feasibility of using a secondary sludge microbial community for selenium removal. We studied the removal patterns of selenate and selenite oxyanions with and without nitrates. We also analyzed the microbial community structure based on the bioreactor bed composition and nitrate concentration in the influent. Based on the findings, we are able to explain how nitrate affects selenate and selenite reduction, as well as the changes in microbial community composition in the biofilm in the presence and absence of nitrate.
2. Materials and Methods
2.1. Zeolite Modification
The natural clinoptilolite zeolite was obtained from the St. Cloud mining company in Winston, NM, USA. The natural zeolite was chemically modified by pre-treating it with sodium and then coating its surface with iron. The final product was termed a sodium pre-treated, iron-modified zeolite, or SPIMZ. Pre-treatment with sodium has been previously proven to increase the iron content on the zeolite surface [
27]. In turn, iron modification increases the zeolite’s capacity for anions. For the pre-treating process, 200 g of natural zeolite was first sieved through a 14–40 mesh to separate the particle size fraction of 0.42–1.41 mm. Details of the zeolite modification process are described elsewhere [
28].
The properties of zeolites used in the column experiments were used to interpret and assess the effect of natural zeolite modification on selenium oxyanion sorption capacity. These properties included zeolite size and morphology, specific surface area, elemental composition, elemental distribution, and iron content. The morphology and elemental chemical composition of natural (NZ) and sodium pre-treated iron-modified zeolite (SPIMZ) were analyzed using a S-3400N II scanning electron microscope (SEM) equipped with an energy-dispersive X-ray spectrometer (EDX). The X-ray diffraction pattern of powder natural zeolite and chemically modified zeolite was measured using an X-ray diffractometer (MiniFlex II, PANalytical Empyrean X-ray Diffractometer) with Cu Kα radiation and 1.540 Ǻ wavelength. Nitrogen adsorption/desorption isotherms were used to determine the Brunauer, Emmett, and Teller (BET) specific surface area and Barrett–Joyner–Halenda (BJH) pore size distribution of sodium pre-treated iron-modified zeolites and natural zeolites, using an ASAP 2050 micropore analyzer (Micromeritrics, Norcross, GA, USA). The specific surface area of NZ was 9.01 m
2/g, while the specific surface area of SPIMZ was 9.62 m
2/g. The nitrogen adsorption was carried out at 77 K. The details of zeolite characterization can be found elsewhere [
28].
2.2. Synthetic Wastewater Preparation
Each liter of influent solution was prepared by mixing either 1 mL of 0.01 M of sodium selenite (Na2SeO3) or sodium selenate (Na2SeO4) stock solution with 1 L of DI water, to prepare a solution with a total Se concentration of 790 µg/L. The nutrients needed for microbial growth were also added to the influent solution. The nutrients for the selenium-reducing bacteria consisted of the following elements (concentrations in mg/L in parentheses): MgSO4 (44), CaCl2·2H2O (140), FeCl2·4H2O (2), MnSO4·H2O (3.4), (NH4)6Mo7O24·4H2O (1.2), CuSO4 (0.8), Zn(NO3)2·6H2O (1.8), and Ni(NO3)2·6H2O (0.3). In addition, 1 g/L of yeast extract and 2 mL/L of MEM vitamin solution were used to enhance the growth of the selenium-reducing bacteria. Lactate, 2.24 g/L, was used as carbon source and electron donor, while a final concentration of 114 mg/L NH4Cl and 28 mg/L Na2HPO4 were added as nitrogen and phosphorous sources. The pH was adjusted to 7.5 by adding sodium hydroxide.
2.3. Biofilter Setup and Operation
The biofiltration experiment included four filtration columns with an internal diameter of 50 mm and a filling length of 450 mm. The bed composition of each column is shown in
Table 2. Natural zeolites and SPIMZ were well mixed when packing the latter columns to distribute SPIMZ evenly throughout the entire column. Approximately 200 mL of anaerobic sludge from the Las Cruces, NM, USA, Wastewater Treatment Plant was added as the microbial inoculum. The sludge was added while packing the columns with the medium to ensure the sludge spread all over the column. Before the addition of sludge, the zeolite medium was hydrated by injecting nutrient solution prepared without selenium species or lactate. The upper and lower ends of the columns were packed with gravel layers of approximately 1-inch thickness to retain and avoid flushing out the zeolite medium. The fully packed columns were positioned vertically. Then, the prepared influent was pumped vertically upwards, continuously through the column, using a peristaltic pump at a flow rate of 3 mL per minute, and the empty bed contact time of the biofilter was 300 min. This flow direction was selected to ensure that the columns remained fully saturated throughout the experiment.
2.4. Effect of Presence of Nitrate Anions
Compared with selenite and selenate, nitrate is a better electron acceptor. The presence of nitrate may interfere with the bioreduction and adsorption of the zeolite biofiltration process [
29]. Therefore, selenium oxyanion removal efficiencies were studied in the presence of nitrate ions in the selenium-contaminated water. The initial selenium (selenate or selenite) concentration (790 µg/L), flow rate (3 mL/min), pH (7.5), and bed composition remained constant. Sodium nitrate was added to the synthetic selenium solution, so that the final nitrate concentration was 620 mg/L. The nitrate concentration, therefore, was about 1000 times higher than the selenium oxyanion concentration.
2.5. Analytical Procedures
Samples from columns A, B, C, and D were collected twice a week. These samples were filtered through 0.45 µm filters to exclude biomass. Of the filtrate, 10 mL was preserved for total dissolved selenium analysis by adding 100 µL of analytical-grade nitric acid. The total soluble selenium concentrations in the filtrate were analyzed using a Perkin Elmer ELAN DRC-e inductively coupled plasma mass spectrometer (ICP-MS).
2.6. DNA Extraction Method and Bacterial Community Analysis
Bacterial community analysis was conducted before and after adding nitrate to identify the microorganisms responsible for the selenium reduction within the columns. The DNA extraction process was performed using a DNeasy PowerSoil Kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol for the collected zeolite samples. Zeolite samples were collected approximately one inch inside from each top and bottom end of the columns to test the availability of microorganisms throughout each column. After DNA extraction, DNA quantity and quality were analyzed using a microplate spectrophotometer with a Take3™ multi-volume plate (Epoch, BioTek, Winooski, VT, USA), before 16S rRNA sequencing.
Bacterial community analysis was performed via next-generation sequencing in the MiSeq Illumina platform. Amplicon sequencing of the V4 region of the 16S rRNA gene is performed with the barcoded primer set 515f/806r designed by [
30] and following the protocol by the Earth Microbiome Project (EMP)
https://earthmicrobiome.org/protocols-and-standards/16s/ (retrieved on 10 May 2023) for the library preparation. PCR amplifications for each sample were performed in duplicate, then pooled and quantified using an accublue kit. A no-template control sample was included during the library preparation as a control for extraneous nucleic acid contamination. Some 240 ng of DNA per sample were pooled and then cleaned using a QIA quick PCR purification kit (QIAGEN). The pool was quantified using the qubit. Then, the DNA pool was diluted to a final concentration of 4 nM and denatured and diluted to a final concentration of 4 pM with a 25% of PhiX. Finally, the DNA library was loaded in the MiSeq Illumina and run using the version 2 module, 2 × 250 paired-end, following the manufacturer’s directions.
Qiime2 software [
31] version 2020.8 was used to analyze the microbial population of the sample based on 16S sequences. The DADA2 method [
32] was used to denoise reads and identify ASVs in each sample. ASVs were compared to various taxonomic classification references (Greengenes, Silva, and EZBioCloud) to construct multiple taxonomy predictions for each sample. ASVs were also used to compute alpha and beta diversity metrics for comparison of the microbiome between samples.
3. Results and Discussion
3.1. Effect of Nitrate Anions and Overall Performance of Columns
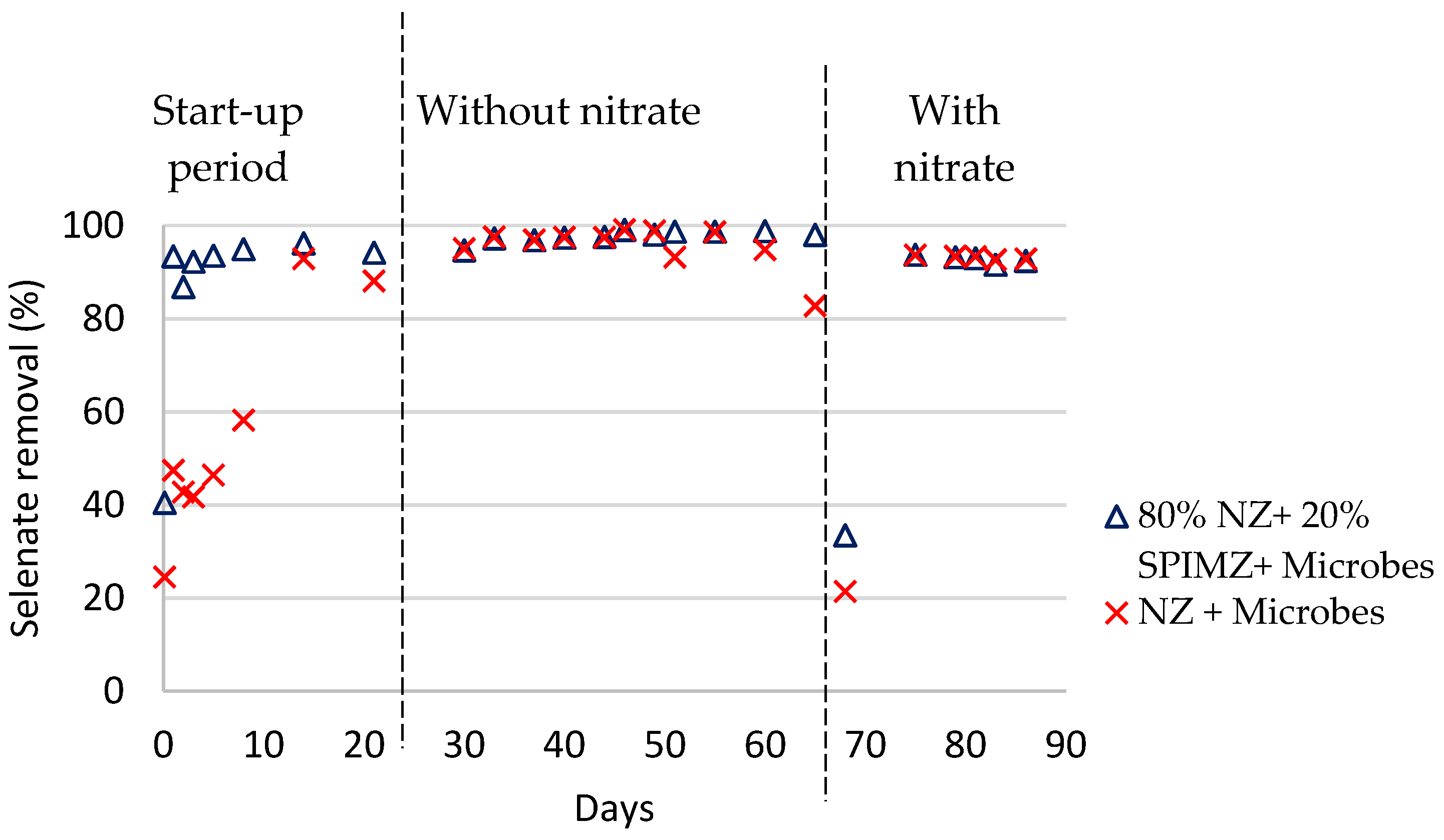
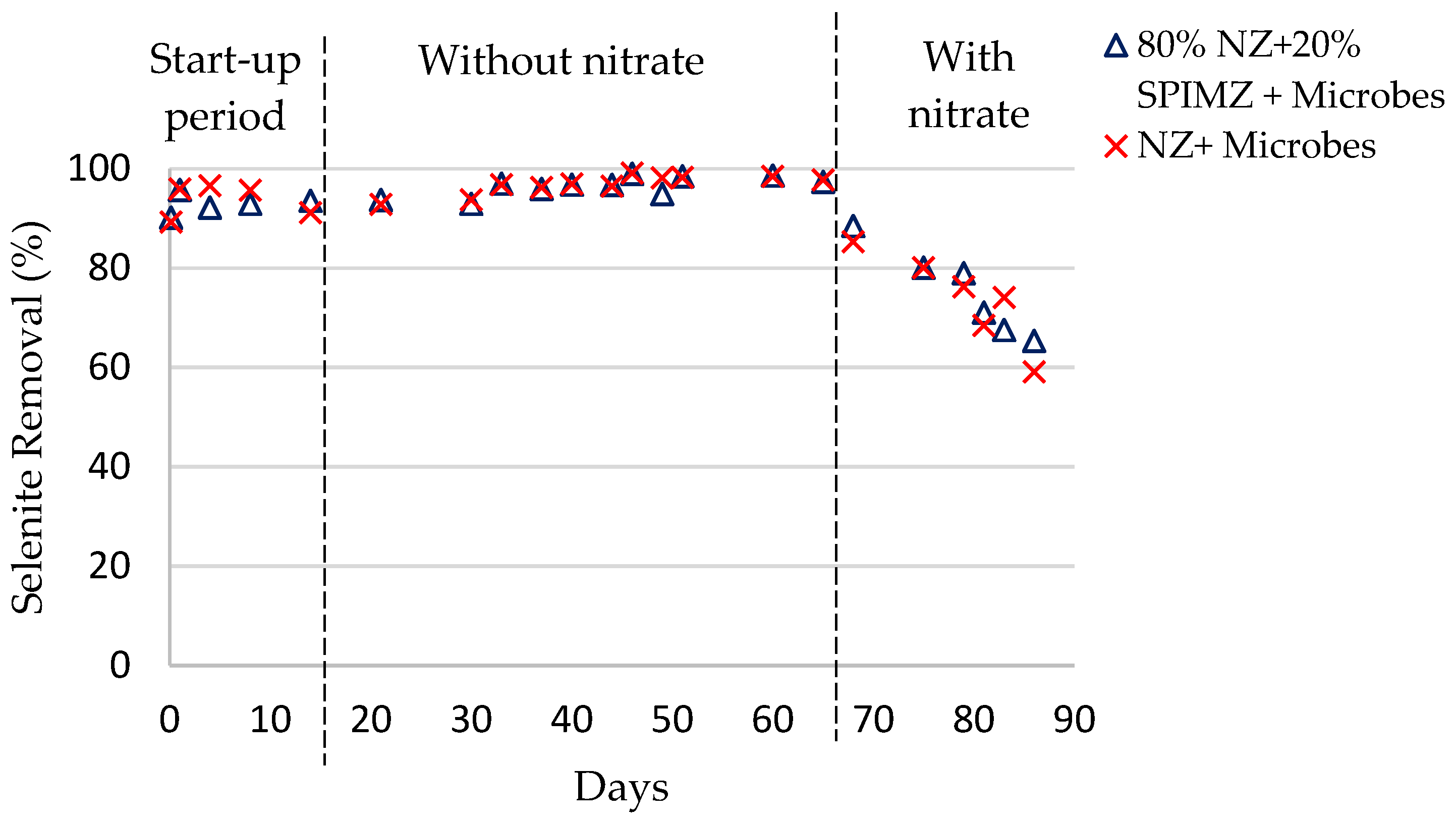
Selenium removal efficiencies in each column are shown in
Figure 1 and
Figure 2. The reactors were operated for 86 days. During the start-up period (first 14 days), feed solution was recirculated until the system reached a steady state. During the first 14 days, there was a higher removal of selenite than selenate, including the columns containing iron-coated zeolites. This suggests that during the start-up period, the dominant removal process is adsorption by zeolites and the iron coating of zeolites. This trend is consistent with previous research conducted using different adsorbents to remove selenium species from water. Compared to selenate, a higher extent of selenite uptake and higher adsorption capacities were also reported by others [
33,
34]. This is likely due to the adsorption of selenite and selenate onto surface groups of zeolites to form inner-sphere and outer-sphere surface complexes, respectively [
35]. Additionally, selenate removal during the first 14 days in the column containing modified zeolite was twice as high as in the column containing natural zeolite only. In addition, during the first 14 days, the maximum selenite removal was 89% in the column with modified zeolite. This result is evidence that the iron coating on the zeolite surface significantly increases the removal of selenium oxyanions. The role of surface complexation in the sorption of selenium oxyanions onto the zeolite surface, size exclusion effects, and impacts of zeolite modification on selenium removal are discussed in detail elsewhere [
28].
During days 14–65, bioreactors operated only with selenium oxyanions as electron acceptors. Regardless of the bed composition, selenium removal in all reactors stabilized around 99%. This suggests that a specialized selenium-reducing microbial population developed inside the columns within 14 days. The results reveal the dominant removal mechanism in the early stage was zeolite adsorption, and later, microbial reduction with the growth of microorganisms. So, in the beginning, the chemical modification has highly affected efficiencies, especially for selenate removal, despite the columns containing a relatively small amount (20%) of coated zeolites. In the beginning, the microbial reduction is low because aerobic conditions still prevail inside the columns, and time is required for the system to become anaerobic. This condition highly affects selenium reduction, as microbial selenium reduction only happens under anaerobic conditions.
After 65 days, the bioreactors were operated with nitrate (620 mg/L) in addition to the selenium oxyanions. The experiments were conducted at a mole ratio of approximately 1:1000 influent SeOx2−: NO3−. Selenate appears to be reduced more effectively compared to selenite. In addition, the system appears to have reached a steady state in the selenate columns, but not in the selenite columns.
When comparing the overall removal in each column, both selenate and selenite removal efficiencies decreased compared to the runs without nitrate. In the presence of nitrate anions, selenate removal decreased from 99 to 93%, and selenite removal decreased from 99 to 60%. A possible explanation hinges on the mechanisms of selenium removal in the current treatment system. These mechanisms are adsorption by zeolite media and biological reduction by selenium-respiring bacteria. The availability of nitrate anions influences both processes. In adsorption, the presence of other anions, such as sulfates, nitrates, and chromates, creates competition and reduces selenium oxyanion adsorption.
During biological reduction, anions act as competing electron acceptors, causing a decrease in selenium reduction, depending on the redox potential of anions present. The following Equations (1) through (5) summarize the overall reduction reactions involving selenate, selenite, nitrate, and nitrite:
Indeed, the reduction potential for the SeO
42−/Se
0, SeO
32−/Se
0, and NO
3−/NO
2− couples are +690 mV, +210 mV and +433 mV, respectively, and therefore selenate reduction occurs prior to nitrate reduction [
3,
29]. Hence, selenate reduction is not affected significantly by nitrate. However, selenite reduction is considerably negatively affected. This is because the SeO
32−/Se
0 redox potential is lower than the NO
3−/NO
2− redox potential; thus, NO
3− reduction takes place prior to selenite reduction. After 2 days from the introduction of nitrate, selenate removal in both columns initially was reduced to approximately 25%, but then increased again over the next 7 days and stabilized at approximately 93%. These results suggest that it takes about 9 days of acclimation for selenate-reducing bacteria to adapt to competitive inhibition, in agreement with previous studies [
12]. The initial pH of the effluent was kept at 7.5, because water with acidic or alkaline pH values tends to be toxic to microorganisms. Moreover, pH 7–8 is favorable in terms of the rate of the selenium reduction reaction [
36]. However, the pH values in the treated water were reduced, ranging from 6.0 to 6.8. As microorganisms consume the organic compound, which is lactate in the current study, using it as their energy source, they produce metabolic byproducts including volatile fatty acids [
37]. These fatty acids are weak acids that can dissociate in water and release hydrogen ions (H
+), thereby lowering the pH of the water.
3.2. Microbial Community Analysis
The results indicate that introducing nitrate increased the diversity of the microbial community and negatively correlated with Se removal efficiencies in both selenate and selenite reduction, which suggests that certain functional bacteria might mediate selenium reduction instead of all microorganisms present. Compared to selenate removal, selenite removal was affected more adversely. Overall, the similarity of the microbial community composition is highly influenced by the selenium speciation in the influent, and is independent of bed composition in the absence of nitrate. When nitrate is present in the influent with selenium oxyanions, the microbial community composition drastically changes, regardless of selenium speciation. This is likely because the nitrate concentration is 1000 times higher than the selenium concentration, and the high nitrate concentration shapes the community structure. However, the difference in removal efficiencies among selenium oxidation states shows that the selenium oxidation state highly influences the treatment efficiency in the presence of nitrate anions.
This study reveals that anaerobic sludge from the Las Cruces, NM, USA, wastewater treatment plant contains numerous bacteria that can play an active role in reducing oxidized selenium compounds. Before introducing nitrate, bacterial communities in the bioreactors display phylogenetic diversity within three major phyla (Firmicutes, Bacteroidota, and Proteobacteria) of the bacterial domain. Before introducing nitrate, members of the classes Bacteroidia and Negativicutes dominated the columns, regardless of the selenium speciation and bed composition. Most of the selenium-reducing bacteria belong to negativicutes [
18]. After introducing nitrate, Clostridia became significantly dominant.
The genus
Megasphaera (
Veillonellales-Selenomonadales) represented about 13–22% of the total bacteria in all columns when nitrate was absent, and it disappeared after introducing nitrate. Unsurprisingly, the genus
Megasphaera has not shown denitrification abilities in previous studies [
38]. However, this genus has not been previously reported as a selenium-reducing bacterium. The possible reason that affected the disappearance of
Megasphaera is that nitrate became toxic, because we used a higher concentration (1000 mg/L). On the other hand, nitrate can be converted to nitrite in anaerobic respiration, which has been identified as a toxin for ruminal microorganisms. The toxicity of nitrite is proven by an experiment conducted with a ruminal microorganism,
Selenomonas ruminantium, which belongs to the same order (
Selenomonadales) as
Megaspheara [
39]. In the absence of nitrate,
Veillonella (
Veillonellales-Selenomonadales) is the most abundant genus among the selenium-reducing microbial community. These two genera belong to the family
Veillonellaceae, the order
Selenomonadales class of negativicutes, and the phylum of Firmicutes. Hence, a significant reduction in the respective taxa could be observed with nitrates’ presence. The
Veillonella family contains six genera, including
Megasphaera and
Veillonella (type genus). Bacteria belonging to this family have been identified as cocci or cocco bacilli in shape, either anaerobic or microaerophilic, and non-motile.
Megasphaera and
Veillonella are the only species belonging to this family that have lactate fermentation capacity [
38] (consistent with the experimental conditions of the current research, given that we used lactate as the organic source). This family has been found in numerous human clinical samples and samples of other mammals such as sheep and pigs. Some species of this family have been identified as opportunistic pathogens for animals, including humans.
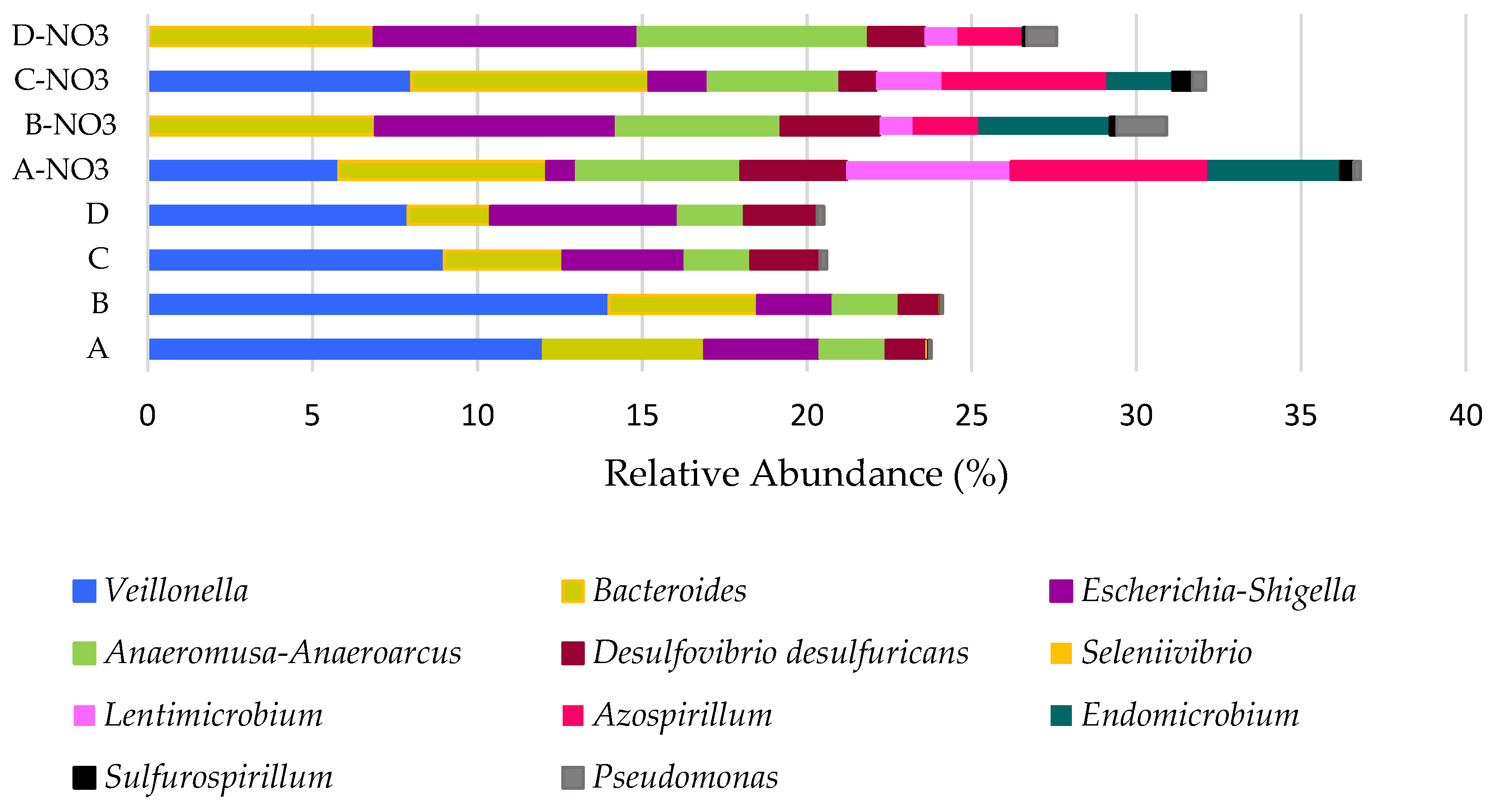
In the absence of nitrate,
Veillonella, Bacteroides, and
Escherichia-Shigella were identified as the most abundant previously known selenium-reducing bacteria.
Anaeromusa-Anaeroarcus (
Veillonellales-Selenomonadales),
Lentimicrobium (
Sphingobacteriales),
Endomicrobium (
Endomicrobiales), and
Azospirillum (
Azospirillales) were barely detectable or not detectable when nitrate was absent, but they became among the dominant genera when nitrate was present in the feed (
Figure 3). These genera were previously known as denitrifying bacteria [
21,
22,
25]. Further,
Bacteroides,
Escherichia-Shigella, and
Azospirillum could be considered the functional bacteria for selenium respiration when nitrate is present.
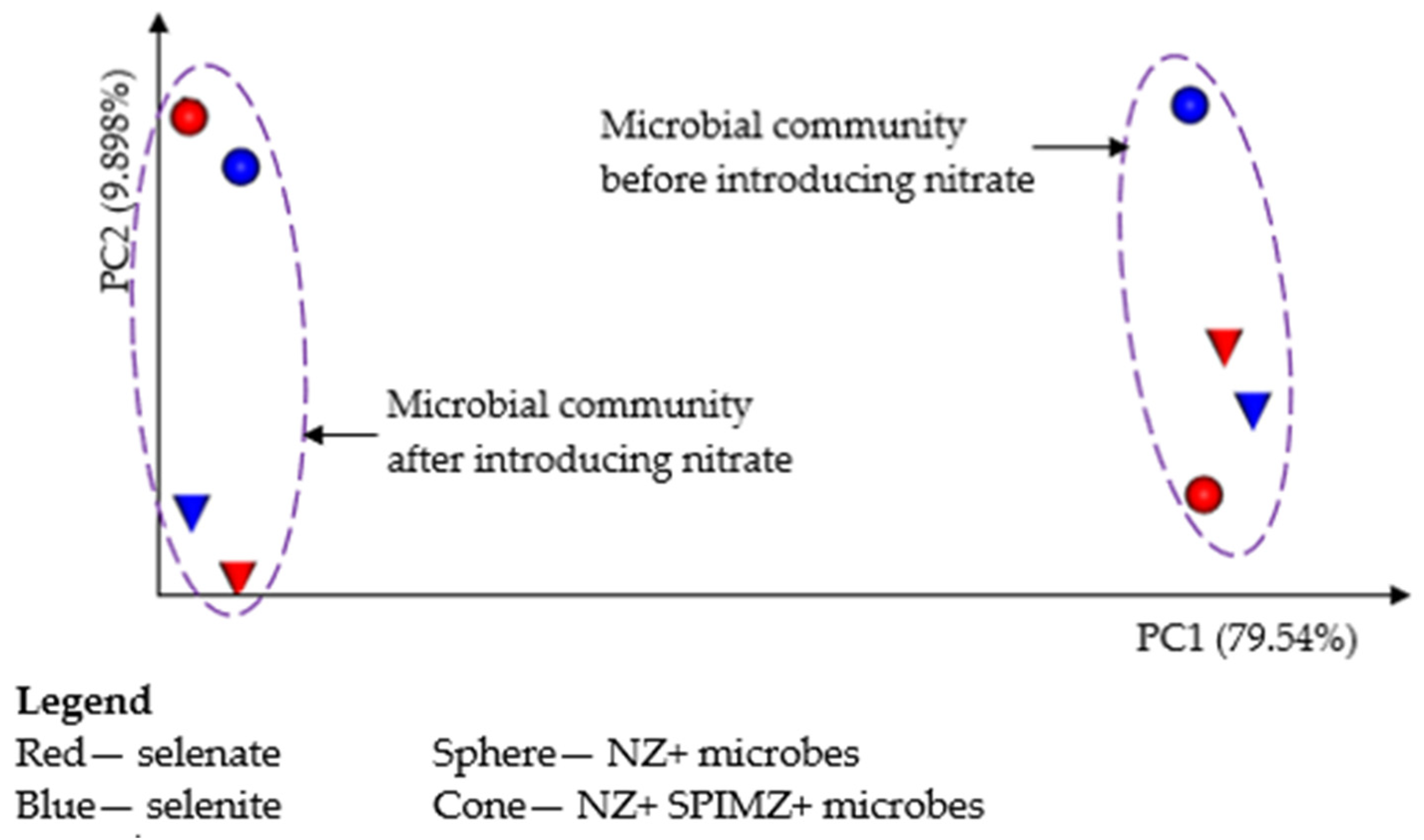
A principal coordinate analysis (PCoA) bi-axis plot based on weighted uniFrac for zeolite samples after treatment of the influent containing selenium with the absence and presence of nitrate is shown in
Figure 4. The most important trend is that the samples before introducing nitrate show higher values for axis 1 than those after introducing nitrate, which tends to cause the microbial compositions to cluster into two different groups. This is additional evidence to support that nitrate significantly shaped the makeup of selenium-reducing microorganisms.
No significant difference in the diversity of microbial communities was found between selenium speciation in the feed source. Before adding nitrate, all columns had a similar microbial composition, whereas after adding nitrate, there was a completely different community structure, while a similar composition exists in all columns. This finding confirms that nitrate anions exert a greater influence on the microbial community. This is reflected by the Shannon index (evenness and richness), Faith PD (richness and diversity), and Pielou’s evenness index (evenness), as given in
Table 3. No notable difference among the indices with respect to the adsorbent or selenium species exists. However, all the indices increased after introducing the nitrate anions, indicating that the evenness, richness, and diversity of microbial community increased in the presence of nitrates, which in turn indicates that the number of different species present in the bioreactors increased, and that the size of the population in each species was more evenly distributed than that in the absence the nitrate [
40].
4. Conclusions
Modification of zeolite positively impacted selenate removal. During the first 14 days, selenate removal in the modified zeolite column was twice as high compared to the column containing natural zeolite only. The growth period for selenium reducers is 14 days. Following the 14-day growth period, maximum removal reached 99% and remained stable in all four columns.
Based on this study, the presence of nitrate had a negative impact on selenium bioremediation using chemically modified zeolite columns. A more pronounced decrease in selenium removal efficiency was observed for selenite (reduced to 60%) than for selenate (reduced to 93%). Hence, pre-treatment might be required when nitrate is present along with selenite. Results suggest that gut microorganisms can endure nitrate after 9 days of acclimation. The selenium removal efficiency is independent of the bed composition of zeolite, as all four columns show 99% removal efficiency in the absence of nitrates. The alteration of the microbial community structure observed with the introduction of nitrate also had a negative impact on selenium removal, with a decrease in the abundance of selenium-reducing bacteria observed.
In the absence of nitrate, biofilm communities mainly comprise selenium-reducing reducing bacteria (Veillonella, Bacteroides and Escherichia). In contrast, the presence of nitrate led to mostly denitrifying bacteria (Anaeromusa-Anaeroarcus, Lentimicrobium, Azospirillum and Endomicrobium). Bacteroides, Escherichia-Shigella and Azospirillum are likely to be the functional bacteria for selenium respiration when nitrate is present.
Overall, this study provides valuable insights into the factors that influence selenium bioremediation using chemically modified zeolite columns, and highlights the influence of the presence of nitrate in selenium removal and the reshaping of microbial communities in bioremediation systems. Based on the findings of this study, biofiltration with modified zeolite media could be used to treat selenium-contaminated wastewater in situ. The modified zeolite, in conjunction with selenium reducing bacteria, could also be used in permeable reactive barrier applications to remediate selenium-contaminated groundwater.







{kind=link}
{kind=link}
{kind=link}
{kind=link}



