
Unexplored Potential: Metabolite Screening of Local Lake Algae Isolated from Al-Asfar Lake in Saudi Arabia

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Information
2.2. Instruments and Reagents
2.3. Sample Preparation
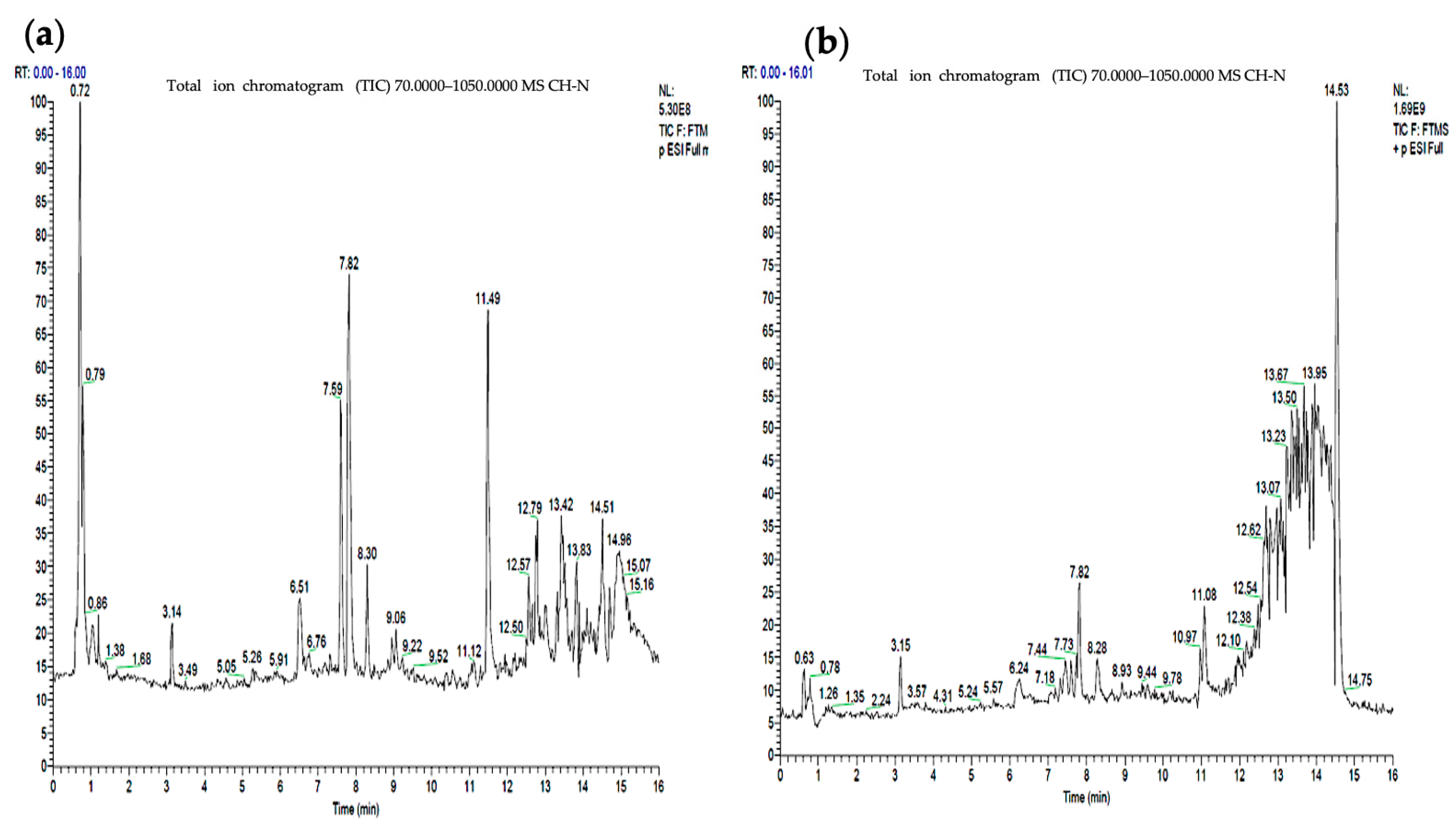
3. LC-MS Analysis
4. Statistical Analysis
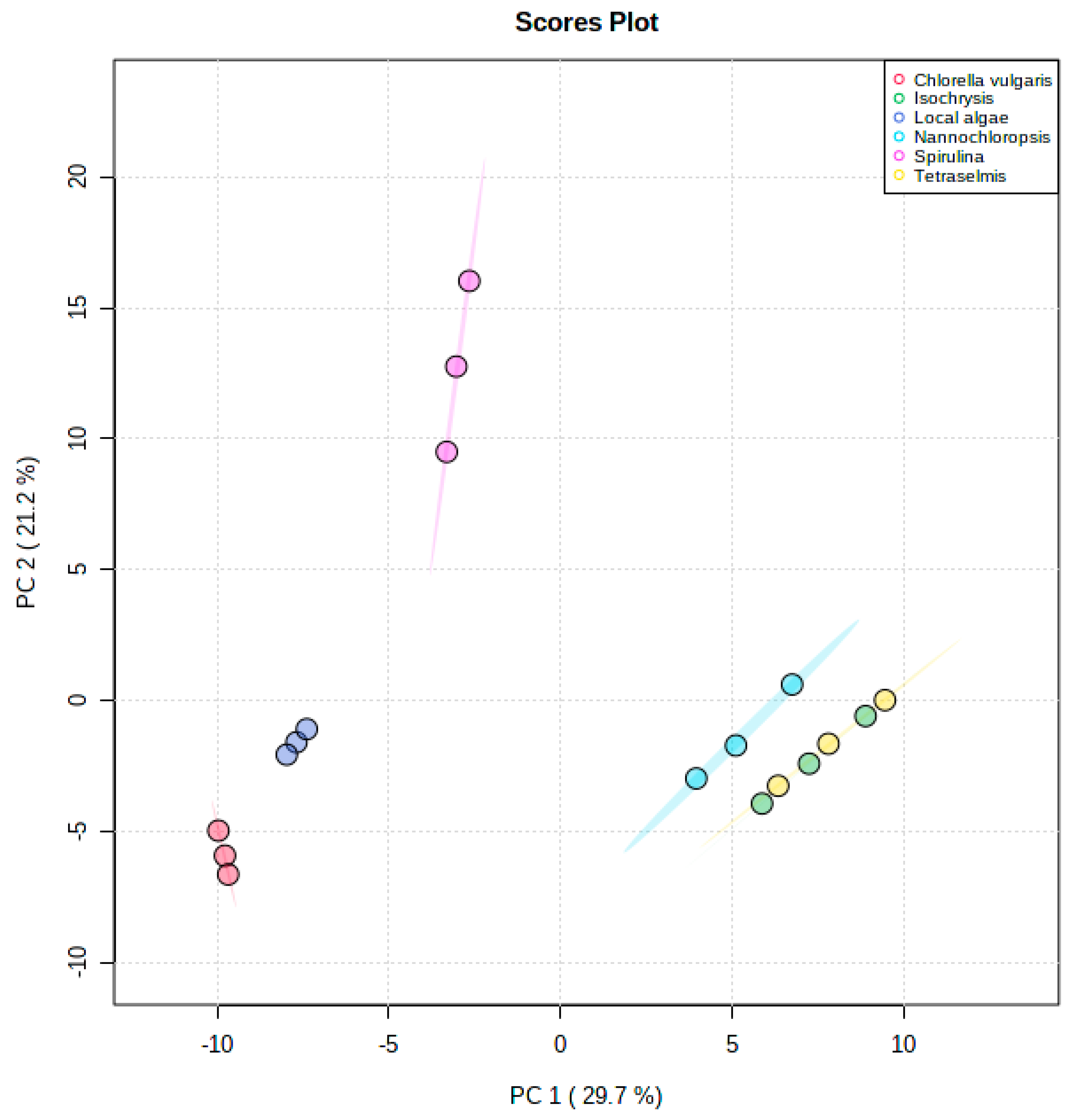
4.1. Principal Component Analysis (PCA)
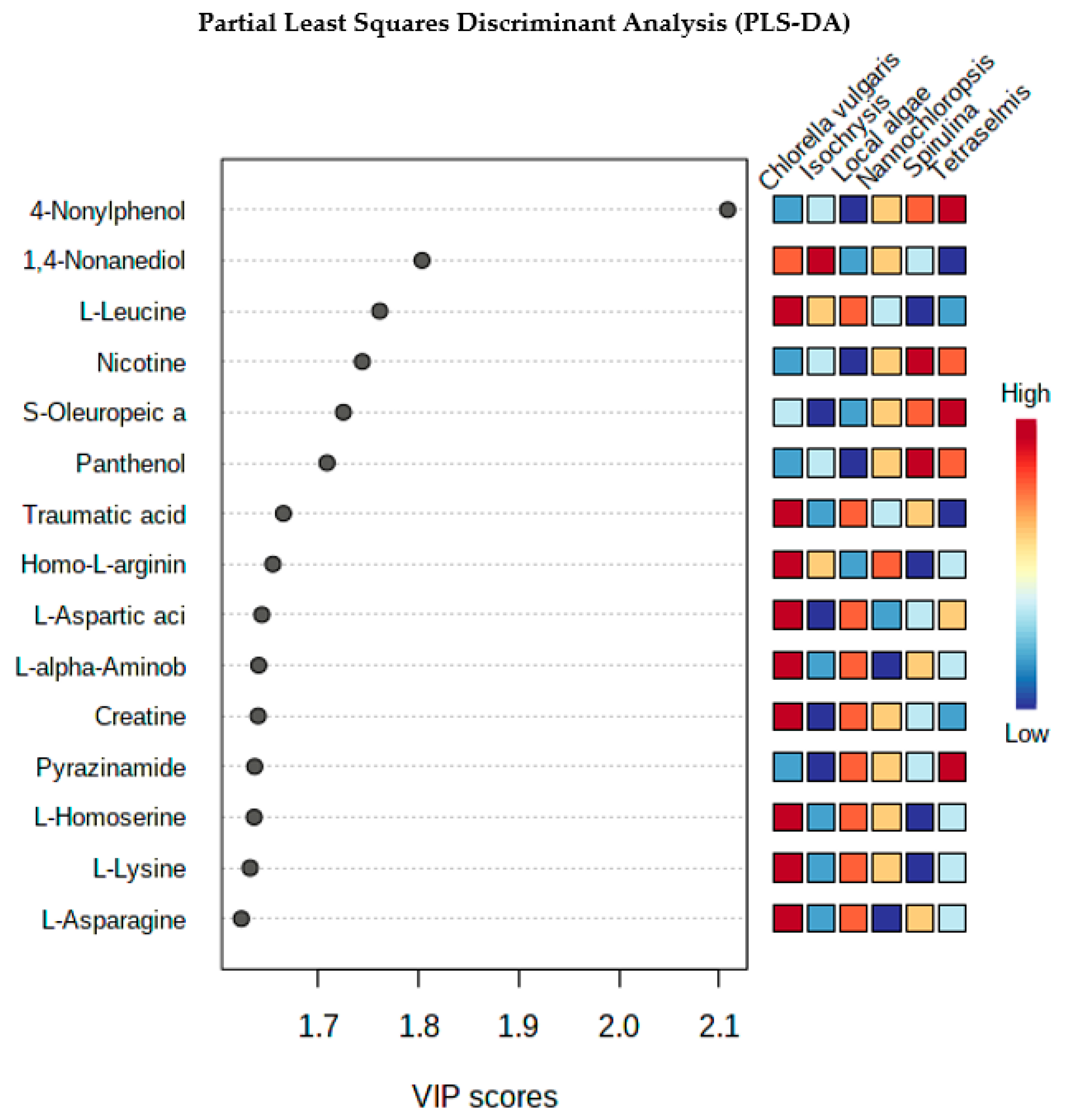
4.2. Partial Least Squares Discriminant Analysis (PLS-DA)
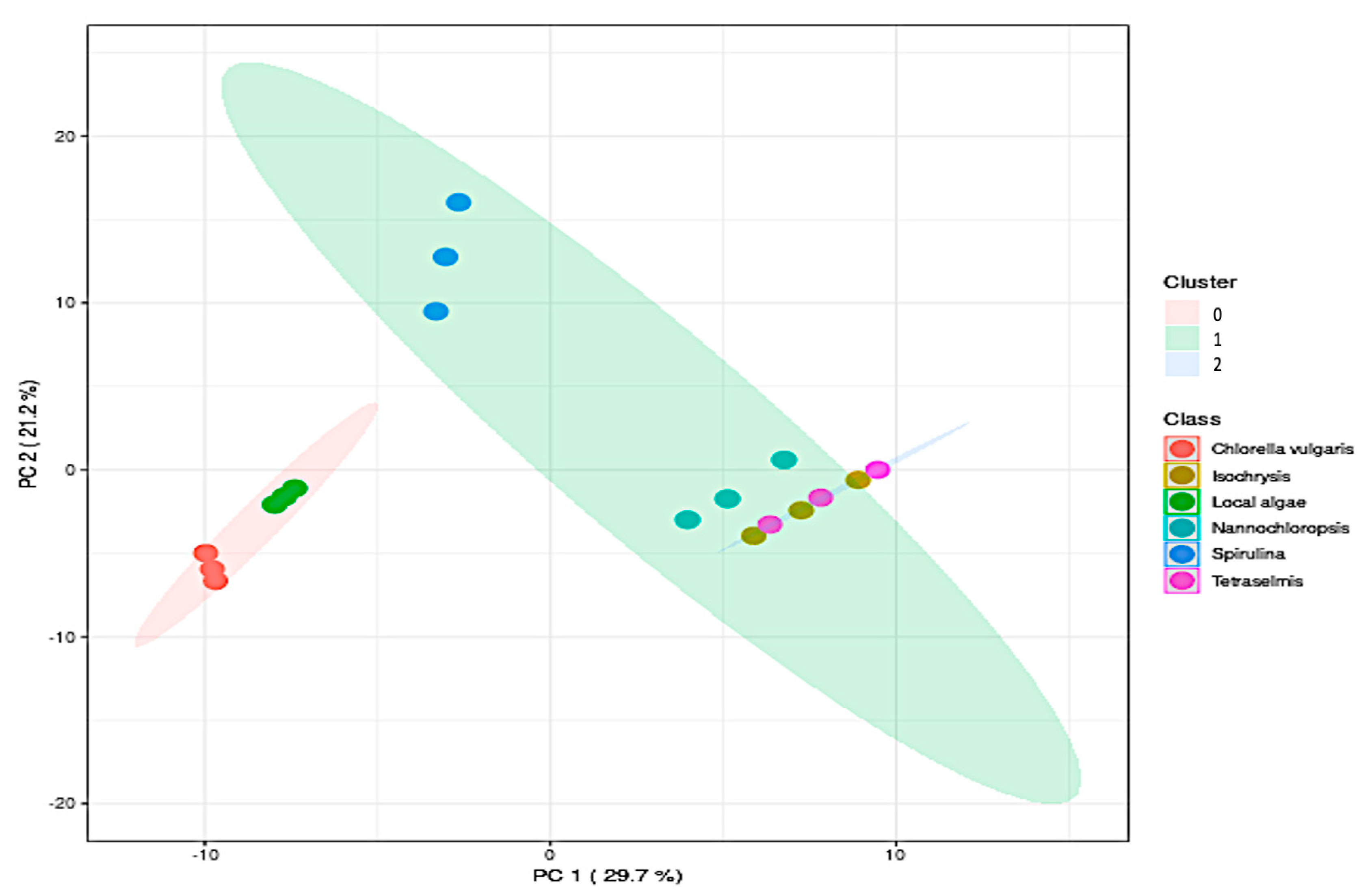
4.3. Hierarchical Cluster Analysis
5. Results
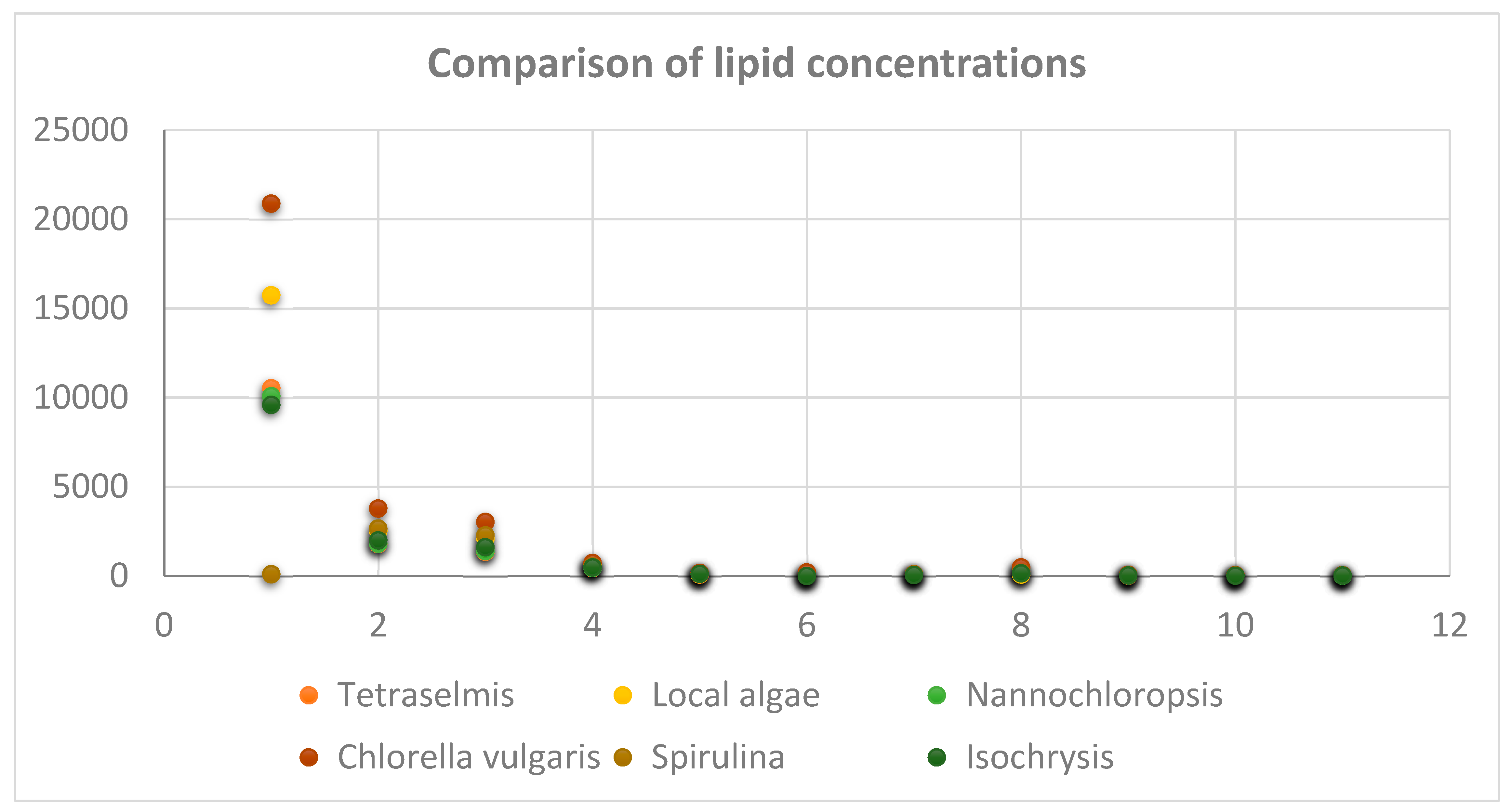
Metabolic Analysis Comparison of Local Algae with Five Known Algae Species
6. Discussion
6.1. The Likelihood of Biodiesel Production
6.2. Disadvantages/Limitations of the Study
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Abderrahman, W.A.; Bader, T.A. Remote sensing application to the management of agricultural drainage water in severely arid region: A case study. Remote Sens. Environ. 1992, 42, 239–246. [Google Scholar] [CrossRef]
- El Mahmoudi, A.S.; Al-Dakheel, Y.Y.; Hussein, A.H.A.; Massoud, M.A. The Environment of Al. Asfar Evaporation Lake in Al. Hassa, Saudi Arabia: An Integrated Study. In Proceedings of the Third International Conference on Water Resources and Arid Environments (2008) and the First Arab Water Forum, Riyadh, Saudi Arabia, 16–19 November 2008; p. 23. [Google Scholar]
- El Mahmoudi, A.S.; Massoud, M.A.; Al-Dakheel, Y.Y.; Hussein, A.H.A.; Center, W.S. Studies of Al Asfar and Al Uyoun Evaporation Lakes Water Quality and the Potential of Its Reuse in Agriculture Activities, Al Hassa Area, KSA. J. King Abdulaziz Univ. Meteorol. Environ. Arid. Land Agric. Sci. 2011, 22, 67–85. [Google Scholar] [CrossRef]
- Sathasivam, R.; Radhakrishnan, R.; Hashem, A.; Abd_Allah, E.F. Microalgae metabolites: A rich source for food and medicine. Saudi J. Biol. Sci. 2019, 26, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Do Nascimento, M.; Ortiz-Marquez, J.C.F.; Sanchez-Rizza, L.; Echarte, M.M.; Curatti, L. Bioprospecting for fast growing and biomass characterization of oleaginous microalgae from South–Eastern Buenos Aires, Argentina. Bioresour. Technol. 2012, 125, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Koller, M.; Salerno, A.; Tuffner, P.; Koinigg, M.; Böchzelt, H.; Schober, S.; Pieber, S.; Schnitzer, H.; Mittelbach, M.; Braunegg, G. Characteristics and potential of micro algal cultivation strategies: A review. J. Clean. Prod. 2012, 37, 377–388. [Google Scholar] [CrossRef]
- Tejano, L.A.; Peralta, J.P.; Yap, E.E.S.; Chang, Y.W. Bioactivities of enzymatic protein hydrolysates derived from Chlorella sorokiniana. Food Sci. Nutr. 2019, 7, 2381–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minhas, A.K.; Hodgson, P.; Barrow, C.J.; Adholeya, A. A review on the assessment of stress conditions for simultaneous production of microalgal lipids and carotenoids. Front. Microbiol. 2016, 7, 546. [Google Scholar] [PubMed] [Green Version]
- Gong, M.; Bassi, A. Carotenoids from microalgae: A review of recent developments. Biotechnol. Adv. 2016, 34, 1396–1412. [Google Scholar] [CrossRef]
- Hallmann, A. Algae biotechnology–green cell-factories on the rise. Curr. Curr. Biotechnol. 2015, 4, 389–415. [Google Scholar] [CrossRef]
- Doan, Y.T.T.; Obbard, J.P. Two-stage cultivation of a Nannochloropsis mutant for biodiesel feedstock. J. Appl. Phycol. 2015, 27, 2203–2208. [Google Scholar]
- Mutanda, T.; Ramesh, D.; Karthikeyan, S.; Kumari, S.; Anandraj, A.; Bux, F. Bioprospecting for hyper-lipid producing microalgal strains for sustainable biofuel production. Bioresour. Technol. 2011, 102, 57–70. [Google Scholar] [CrossRef]
- Duong, V.T.; Ahmed, F.; Thomas-Hall, S.R.; Quigley, S.; Nowak, E.; Schenk, P.M. High protein-and high lipid-producing microalgae from northern Australia as potential feedstock for animal feed and biodiesel. Front. Bioeng. Biotechnol. 2015, 3, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, E.G.; de Carvalho, J.C.; Aulestia, D.T.M.; Gonzalez, O.I.M.; Soccol, C.R. Bioprospection of green microalgae native to Paraná, Brazil using a multi-criteria analysis: Potential for the production of lipids, proteins, and carotenoids. Bioresour. Technol. Rep. 2020, 10, 100398. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Li, S.; Xia, J.M. 3.0: Toward an optimized workflow for global metabolomics. Metabolites 2020, 10, 186. [Google Scholar] [PubMed]
- Bijlsma, S.; Bobeldijk, I.; Verheij, E.R.; Ramaker, R.; Kochhar, S.; Macdonald, I.A.; Van Ommen, B.; Smilde, A.K. Large-scale human metabolomics studies: A strategy for data (pre-) processing and validation. Anal. Chem. 2006, 78, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. Micro-algae as sources of fine chemicals. Microbiol. Sci. 1986, 3, 372–375. [Google Scholar]
- Al-Dakheel, Y.Y.; Hussein AH, A.; El-Mahmoudi, A.S.; Massoud, M.A. Soil, water chemistry and sedimentological studies of Al Asfar Evaporation Lake and its Inland Sabkha, Al Hassa area, Saudi Arabia. Asian J. Earth Sci. 2010, 3(3), 142–162. [Google Scholar] [CrossRef]
- Del Campo, J.A.; García-González, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar] [CrossRef]
- Mendes, R.L.; Nobre, B.P.; Cardoso, M.T.; Pereira, A.P.; Palavra, A.F. Supercritical carbon dioxide extraction of compounds with pharmaceutical importance from microalgae. Inorg. Chim. Acta 2003, 356, 328–334. [Google Scholar] [CrossRef]
- Batista, A.P.; Gouveia, L.; Bandarra, N.M.; Franco, J.M.; Raymundo, A. Comparison of microalgal biomass profiles as novel functional ingredient for food products. Algal Res. 2013, 2, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Ścieszka, S.; Klewicka, E. Algae in food: A general review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3538–3547. [Google Scholar] [CrossRef] [PubMed]
- Coronel, C.D.; Do Nascimento, M.; Curatti, L. Effect of matching microalgal strains origin and regional weather condition on biomass productivity in environmental photobioreactors. Bioresour. Technol. Rep. 2019, 5, 104–112. [Google Scholar] [CrossRef]
- Pandey, A.; Srivastava, S.; Kumar, S. Isolation, screening and comprehensive characterization of candidate microalgae for biofuel feedstock production and dairy effluent treatment: A sustainable approach. Bioresour. Technol. 2019, 293, 121998. [Google Scholar] [PubMed]
- Schuelter, A.R.; Kroumov, A.D.; Hinterholz, C.L.; Fiorini, A.; Trigueros, D.E.G.; Vendruscolo, E.G.; Zaharieva, M.M.; Módenes, A.N. Isolation and identification of new microalgae strains with antibacterial activity on food-borne pathogens. Engineering approach to optimize synthesis of desired metabolites. Biochem. Eng. J. 2019, 144, 28–39. [Google Scholar] [CrossRef]
- Sanchez, M.; Bernal-Castillo, J.; Rozo, C.; Rodríguez, I. Spirulina (Arthrospira): An edible microorganism: A review. Universitas Scientiarum. 2003, 8, 7–24. [Google Scholar]
- Borowitzka, M.A. High-value products from microalgae—Their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Morimura, Y.; Tamiya, N. Preliminary experiments in the use of Chlorella as human food. Food Technol. 1954, 8, 179–182. [Google Scholar]
- Konishi, F.; Tanaka, K.; Himeno, K.; Taniguchi, K.; Nomoto, K. Antitumor effect induced by a hot water extract of Chlorella vulgaris (CE): Resistance to Meth-A tumor growth mediated by CE-induced polymorphonuclear leukocytes. Cancer Immunol. Immunother. 1985, 19, 73–78. [Google Scholar] [CrossRef]
- Chovančíková, M.; Šimek, V. Effects of hight–fat and Chlorella vulgaris feeding on changes in lipid metabolism in mice. Biol. Bratisl. 2001, 56, 661–666. [Google Scholar]
- Jong-Yuh, C.; Mei-Fen, S. Potential hypoglycemic effects of Chlorella in streptozotocin-induced diabetic mice. Life Sci. 2005, 77, 980–990. [Google Scholar] [CrossRef]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, H.R. Chlorella vulgaris as a source of essential fatty acids and micronutrients: A brief commentary. Open Plant Sci. J. 2017, 10, 92–99. [Google Scholar]
- Li, L.; Li, W.; Kim, Y.H.; Lee, Y.W. Chlorella vulgaris extract ameliorates carbon tetrachloride-induced acute hepatic injury in mice. Exp. Toxicol. Pathol. 2013, 65, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Ito, K.; Ueno, S.; Kumamoto, S.; Ando, Y.; Yamada, A.; Nomoto, K.; Yasunobu, Y. Oral administration of hot water extracts of Chlorella vulgaris reduces IgE production against milk casein in mice. Int. J. Immunopharmacol. 1999, 21, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.J.; Shin, H.S.; Chai, O.H.; Han, J.G.; Shon, D.H. Inhibitory effect of unicellular green algae (Chlorella vulgaris) water extract on allergic immune response. J. Sci. Food Agric. 2013, 93, 3133–3136. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, J.S.; Blasco, I.M.; Gagliano, H.; Daviu, N.; Román, A.G.; Belda, X.; Carrasco, J.; Rocha, M.C.; Neto, J.P.; Armario, A. Chlorella vulgaris reduces the impact of stress on hypothalamic–pituitary–adrenal axis and brain c-fos expression. Psychoneuroendocrinology 2016, 65, 1–8. [Google Scholar] [CrossRef]
- Makpol, S.; Yeoh, T.W.; Ruslam, F.A.C.; Arifin, K.T.; Yusof, Y.A.M. Comparative effect of Piper betle, Chlorella vulgaris and tocotrienol-rich fraction on antioxidant enzymes activity in cellular ageing of human diploid fibroblasts. BMC Complement. Altern. Med. 2013, 13, 210. [Google Scholar] [CrossRef]
- Kang, H.; Lee, C.H.; Kim, J.R.; Kwon, J.Y.; Seo, S.G.; Han, J.G.; Kim, B.G.; Kim, J.E.; Lee, K.W. Chlorella vulgaris attenuates dermatophagoides farinae-induced atopic dermatitis-like symptoms in NC/Nga mice. Int. J. Mol. Sci. 2015, 16, 21021–21034. [Google Scholar] [CrossRef] [Green Version]
- Alam, F.; Mobin, S.; Chowdhury, H. Third generation biofuel from algae. Procedia Eng. 2015, 105, 763–768. [Google Scholar] [CrossRef]
- Sistiafi, A.G.; Putri, D.N. Biodiesel synthesis from Nannochloropsis oculata and Chlorella vulgaris through transesterification process using NaOH/zeolite heterogeneous catalyst. IOP Conf. Ser. Earth Environ. Sci. 2018, 105, 012053. [Google Scholar]
- Aguoru, C.U.; Okibe, P.O. Content and Composition of Lipid Produced by Chlorella vulgaris for Biodiesel Production. Adv. Life Sci. 2015, 36, 96–100. [Google Scholar]
- Kaushik, N.; Dhup, S.; Jamsheer, K.M. Addition of methyl jasmonate and rutin hydrate at harvest time elicits lipid production in Scenedesmus. Botany 2021, 99, 167–173. [Google Scholar] [CrossRef]
- Roy, M.; McDonald, L.M. Metal uptake in plants and health risk assessments in metal-contaminated smelter soils. Land Degrad. Dev. 2015, 26, 785–792. [Google Scholar] [CrossRef]
- Alam, M.A.; Wan, C.; Zhao, X.Q.; Chen, L.J.; Chang, J.S.; Bai, F.W. Enhanced removal of Zn2+ or Cd2+ by the flocculating Chlorella vulgaris JSC-7. J. Hazard. Mater. 2015, 289, 38–45. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipids Molecular Formula | Glycolipid % | Phospholipid % | Triacylglycerol % |
|---|---|---|---|
| C14:0 | 43.5 ± 7 | 27.8 ± 5.5 | 8.9 ± 1.4 |
| C16:0 | 30.5 ± 3.7 | 30.7 ± 2.4 | 22.1 ± 1.6 |
| C16:1n7 | 37.3 ± 2.4 | 29.2 ± 1.8 | 8.4 ± 0.6 |
| C16:2n4 | 60.7 ± 1.5 | 3.3 ± 0.3 | 22 ± 1.5 |
| C16:3n4 | 90.5 ± 0.6 | 2.6 ± 0.8 | 0.4 ± 0.2 |
| C18:0 | 23.2 ± 7.7 | 15.2 ± 3.7 | 63.9 ± 10.8 |
| C18:1n9 | 12.4 ± 1.0 | 75.9 ± 0.8 | 7.8 ± 0.9 |
| C18:1n7 | 29.6 ± 2.0 | 61.9 ± 2.5 | 5.8 ± 0.5 |
| C18:2n6 | 19 ± 1.9 | 70.4 ± 1.1 | 7.7 ± 0.7 |
| C18:3n6 | 52.2 ± 0.9 | 45.9 ± 0.9 | 0 ± 0 |
| C18:3n3 | 34.0 ± 2.25 | 62.1 ± 3.2 | 3.8 ± 4.0 |
| C18:4n3 | 62.8 ± 7.0 | 33.5 ± 2.4 | 3.9 ± 5.2 |
| C18:5n3 | 53.2 ± 1.2 | 22.9 ± 2.4 | 25.2± 2.5 |
| C20:5n3 | 63.5 ± 1.2 | 30.9 ± 0.4 | 2.1 ± 1.6 |
| C22:6 | 20.3 ± 2.4 | 72.5 ± 0.2 | 3.6 ± 2.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsanie, S.I.; Dhawi, F. Unexplored Potential: Metabolite Screening of Local Lake Algae Isolated from Al-Asfar Lake in Saudi Arabia. Water 2023, 15, 2757. https://doi.org/10.3390/w15152757
Alsanie SI, Dhawi F. Unexplored Potential: Metabolite Screening of Local Lake Algae Isolated from Al-Asfar Lake in Saudi Arabia. Water. 2023; 15(15):2757. https://doi.org/10.3390/w15152757
Chicago/Turabian StyleAlsanie, Sumayah I., and Faten Dhawi. 2023. "Unexplored Potential: Metabolite Screening of Local Lake Algae Isolated from Al-Asfar Lake in Saudi Arabia" Water 15, no. 15: 2757. https://doi.org/10.3390/w15152757
APA StyleAlsanie, S. I., & Dhawi, F. (2023). Unexplored Potential: Metabolite Screening of Local Lake Algae Isolated from Al-Asfar Lake in Saudi Arabia. Water, 15(15), 2757. https://doi.org/10.3390/w15152757