
New Records of Tetraselmis sp. Strains with Biotechnological Potential Isolated from Greek Coastal Lagoons

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
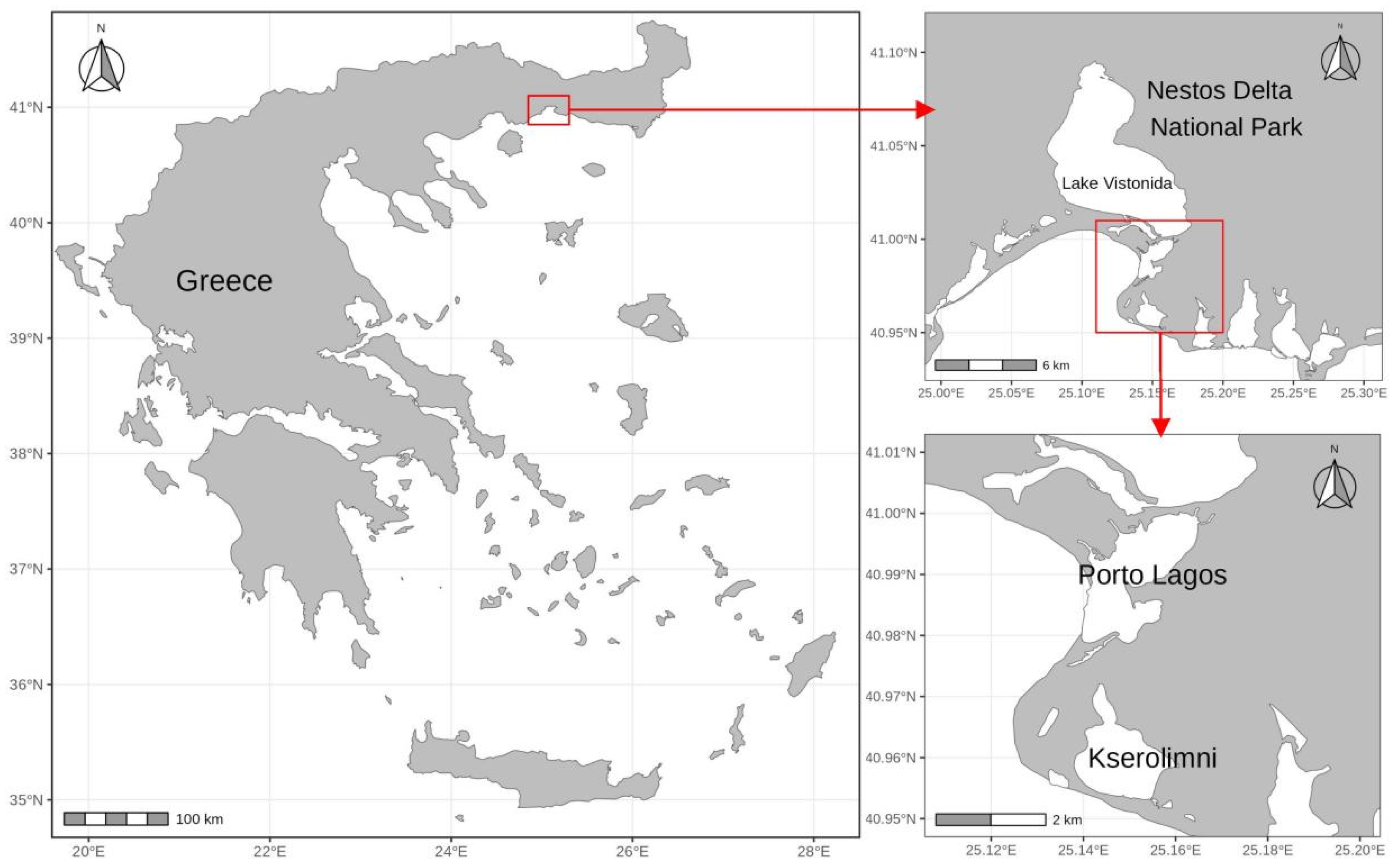
2.1. Sampling
2.2. Strain Isolation and Cultivation
2.3. Microscopic Observation and Morphology
2.4. Molecular Methods
2.5. Culture Kinetics
2.6. Biochemical Analyses
2.6.1. Total Protein Content (TP)
2.6.2. Total Lipid Content (TL)
2.6.3. Total Polysaccharide Content (PS)
2.6.4. Pigment Analysis
2.7. Antioxidant Capacity
2.7.1. TPC
2.7.2. TFC
2.7.3. FRAP
2.7.4. TEAC
2.8. Statistical Analysis
3. Results
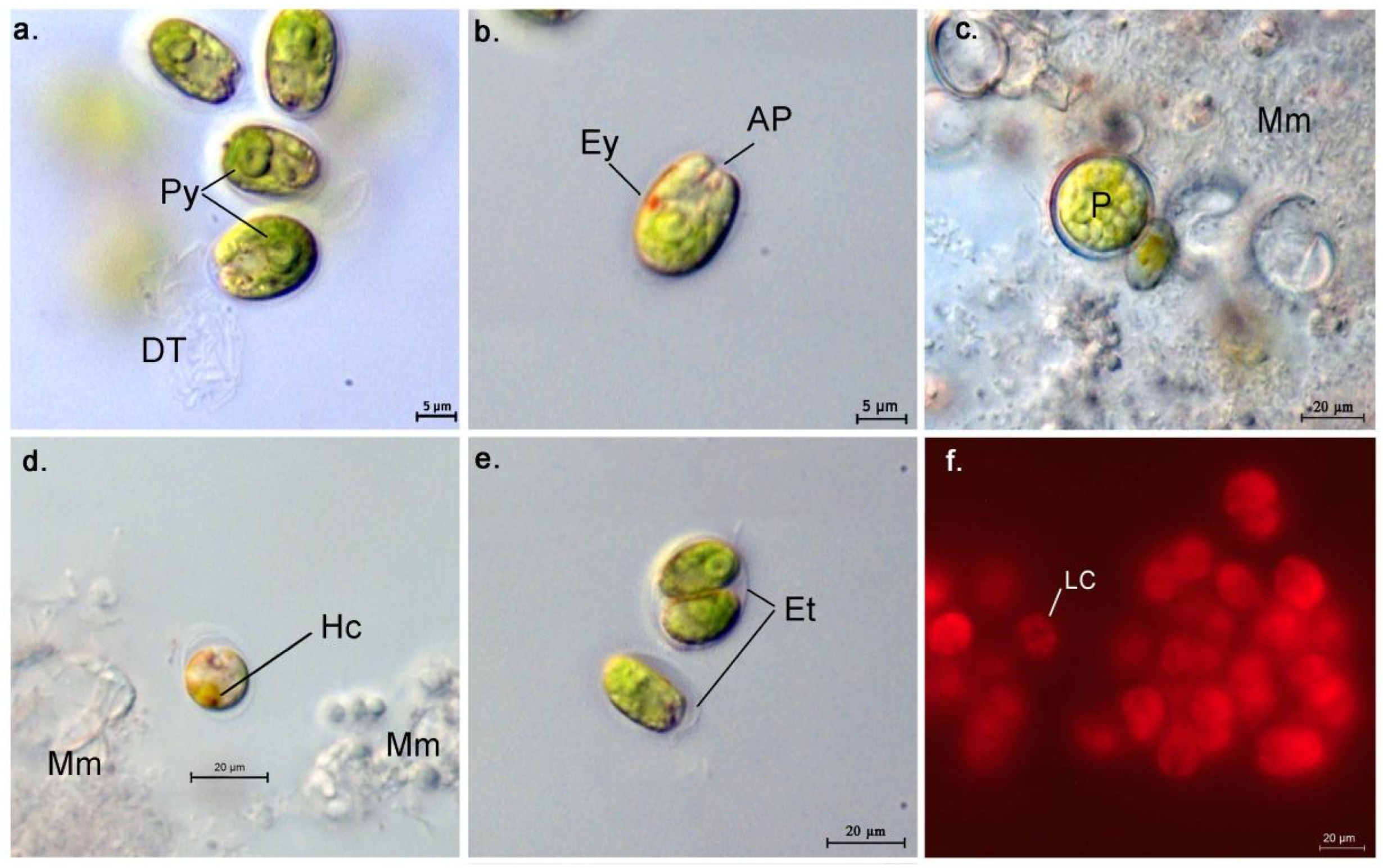
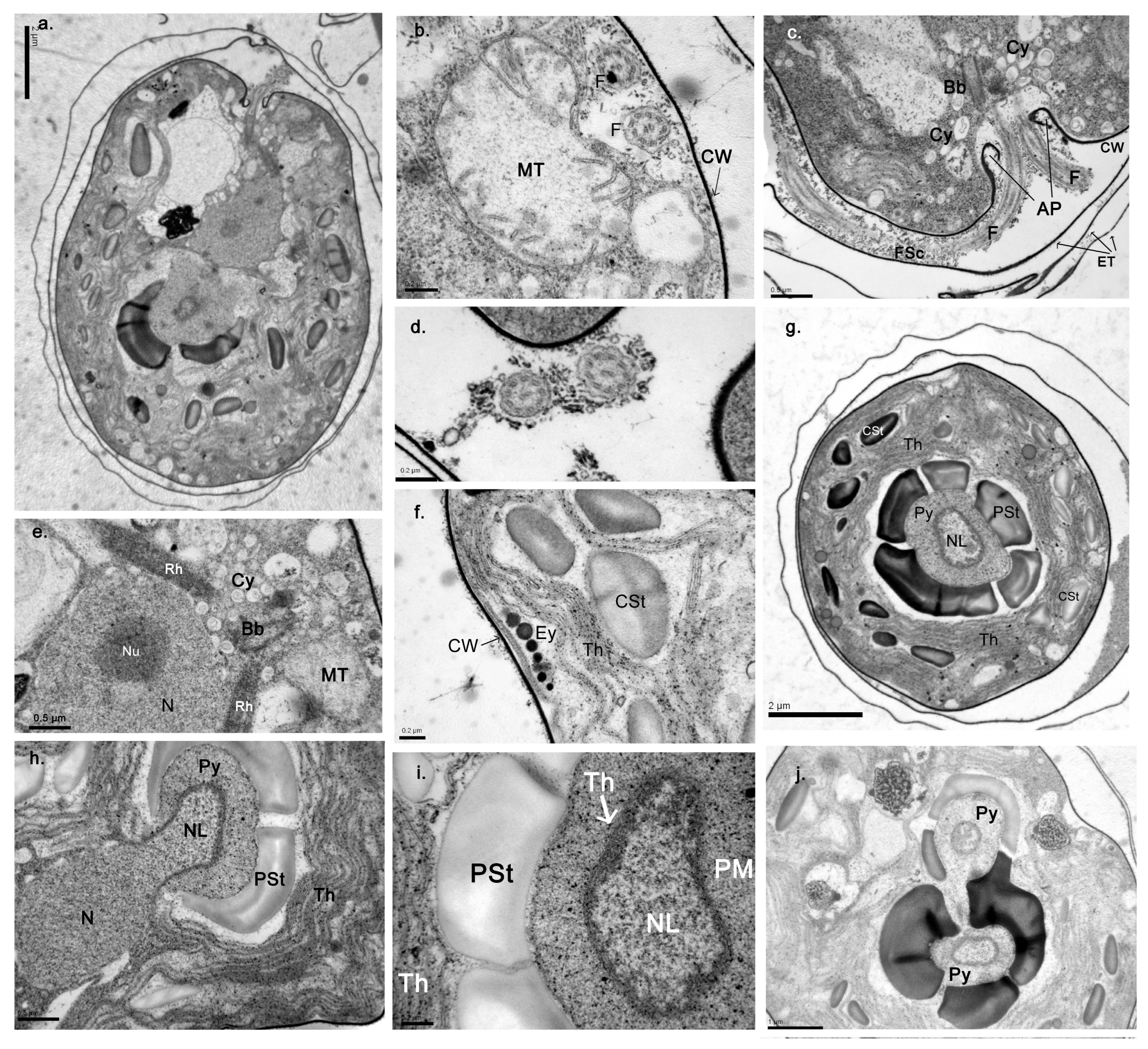
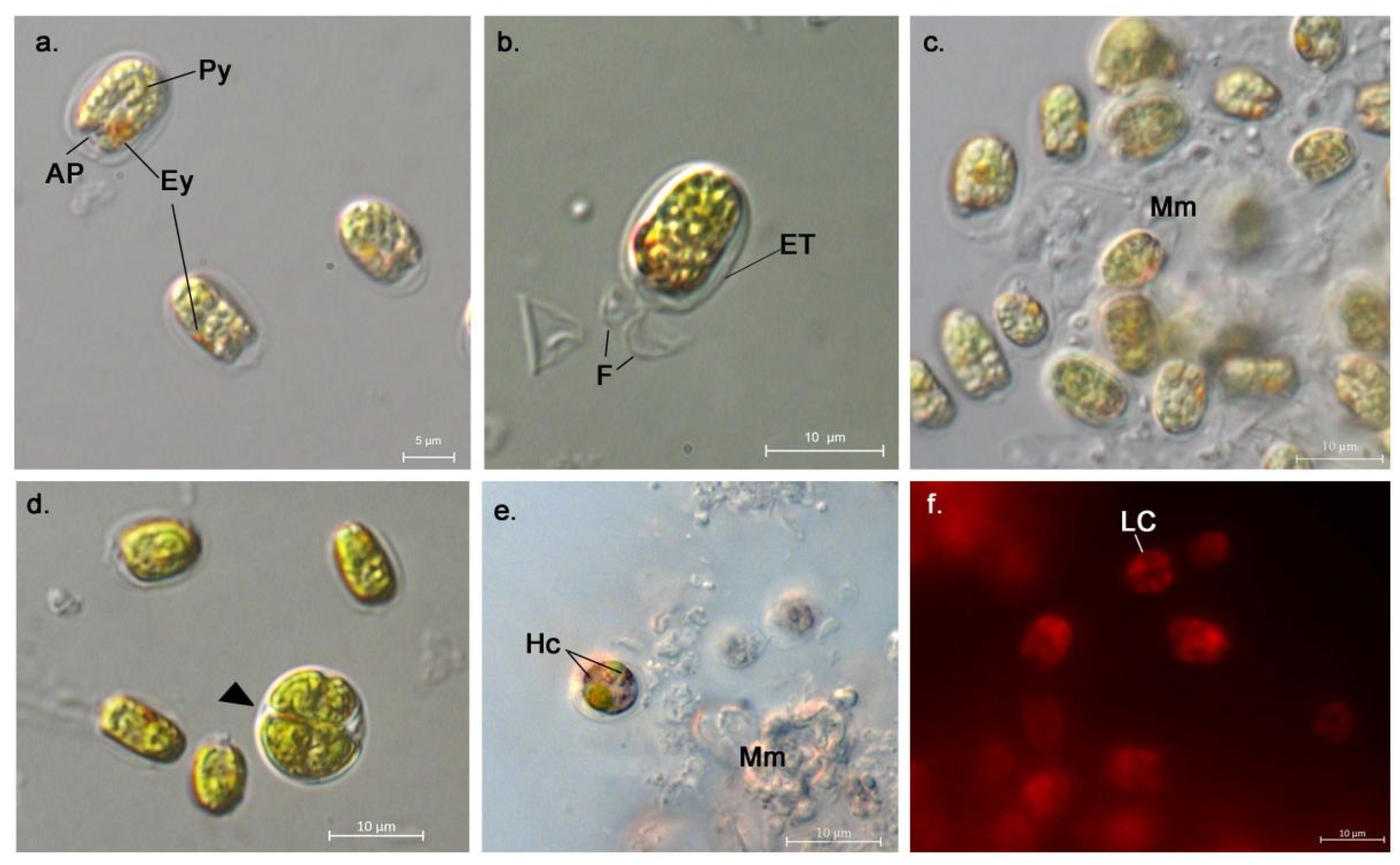
3.1. Morphological Description of the Strains
- AthU-Al PLA 1-2
- AthU-Al KSI 1-3
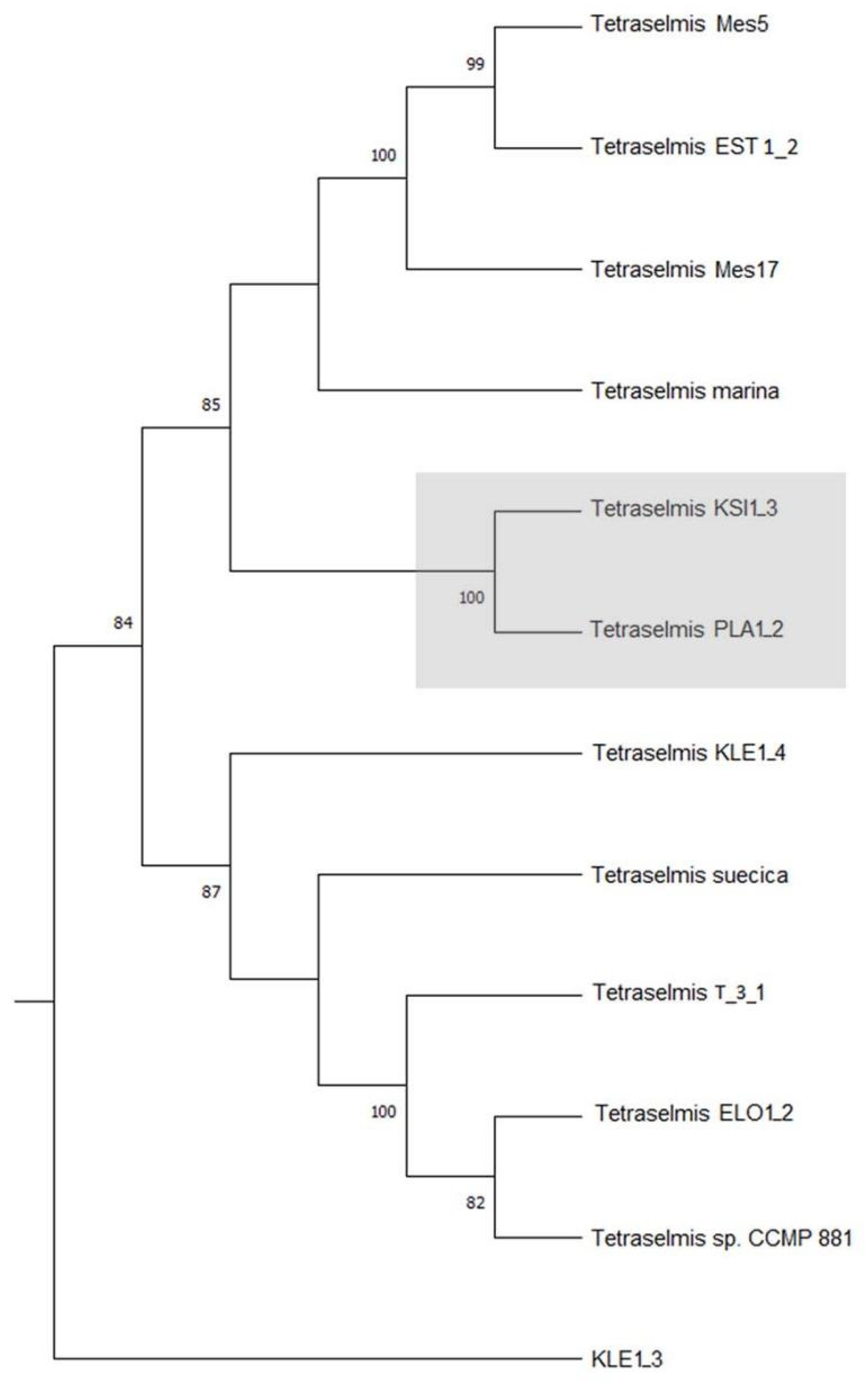
3.2. Molecular Analyses
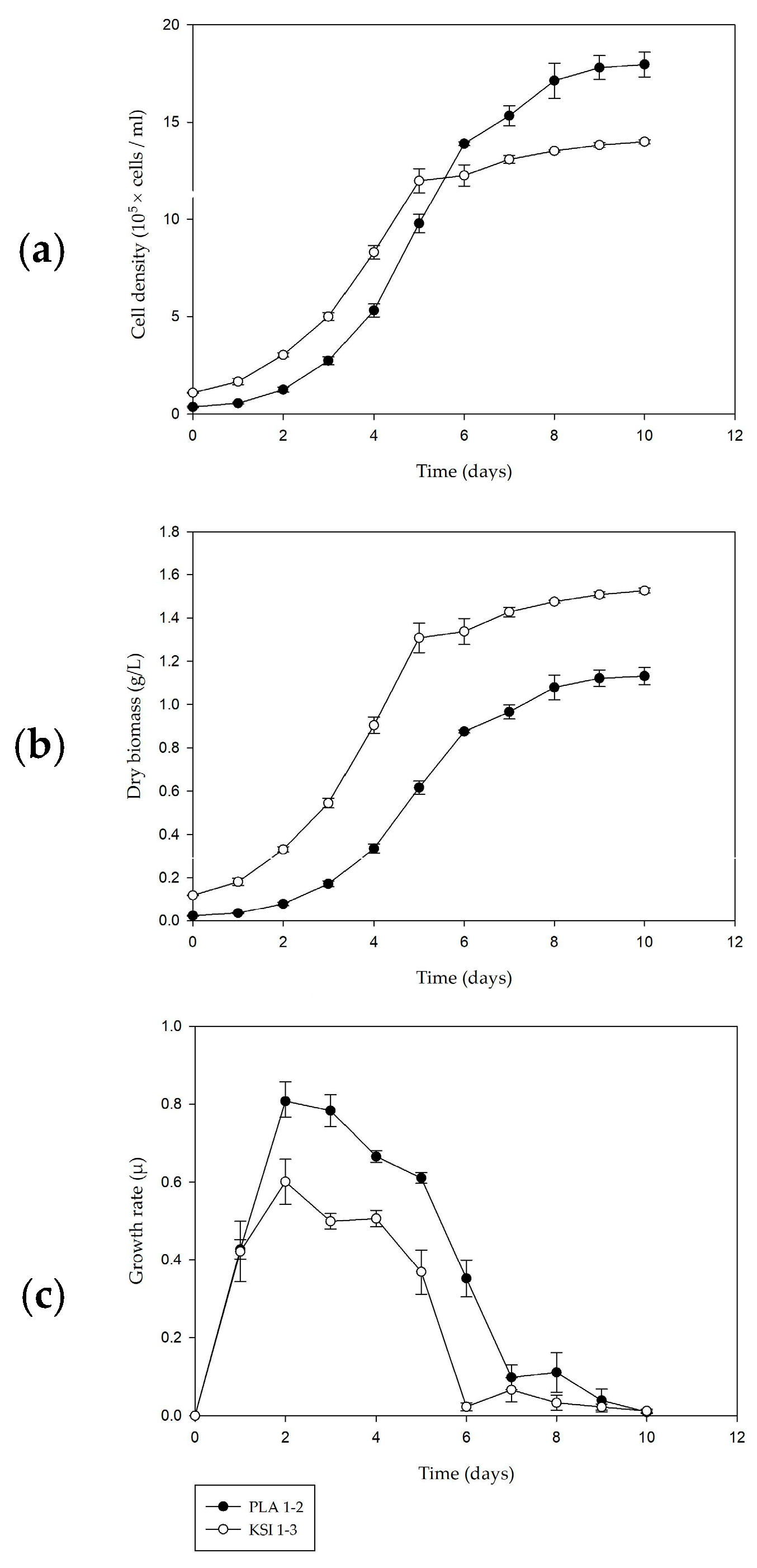
3.3. Culture Kinetics
3.4. Biochemical Analyses
3.5. Antioxidant Capacity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blackburn, S.I.; Lee-Chang, K.J. Microalgae: A renewable resource for food and fuels and more. In Blue Biotechnology: Production and Use of Marine Molecules; La Barre, S., Bates, S.S., Eds.; Wiley-VCH: Weinheim, Germany, 2018; Volume 1, pp. 1–32. [Google Scholar] [CrossRef]
- Newton, A.; Icely, J.; Cristina, S.; Brito, A.; Cardoso, A.C.; Colijn, F.; Dalla Riva, S.; Gertz, F.; Hansen, J.W.; Holmer, M.; et al. An overview of ecological status, vulnerability and future perspectives of European large shallow, semi-enclosed coastal systems, lagoons and transitional waters. Estuar. Coast. Shelf Sci. 2014, 140, 95–122. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Pérez-Ruzafa, I.M.; Newton, A.; Marcos, C. Coastal lagoons: Environmental variability, ecosystem complexity, and goods and services uniformity. In Coasts and Estuaries: The Future; Wolanski, E., Day, J.W., Elliott, M., Ramachandran, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 253–276. [Google Scholar] [CrossRef]
- Tzovenis, I.; Fountoulaki, E.; Dolapsakis, N.; Kotzamanis, I.; Nengas, I.; Bitis, I.; Cladas, Y.; Economou-Amilli, A. Screening for marine nanoplanktic microalgae from Greek coastal lagoons (Ionian Sea) for use in mariculture. J. Appl. Phycol. 2009, 21, 457–469. [Google Scholar] [CrossRef]
- Chantzistrountsiou, X.; Tzovenis, I.; Parmakellis, A.; Economou-Amilli, A. Characterization of Tetraselmis verrucosa f. rubens (Chlorodendrophyceae) strains from coastal lagoons of Western Greece using a multivariate approach. Phytotaxa 2016, 278, 225–240. [Google Scholar] [CrossRef]
- Qazi, W.M.; Ballance, S.; Uhlen, A.K.; Kousoulaki, K.; Haugen, J.E.; Rieder, A. Protein enrichment of wheat bread with the marine green microalgae Tetraselmis chuii–Impact on dough rheology and bread quality. LWT 2021, 143, 111115. [Google Scholar] [CrossRef]
- Schüler, L.M.; Bombo, G.; Duarte, P.; Santos, T.F.; Maia, I.B.; Pinheiro, F.; Marques, J.; Jacinto, R.; Schulze, P.S.; Pereira, H.; et al. Carotenoid biosynthetic gene expression, pigment and n-3 fatty acid contents in carotenoid-rich Tetraselmis striata CTP4 strains under heat stress combined with high light. Bioresour. Technol. 2021, 337, 125385. [Google Scholar] [CrossRef]
- Leliaert, F.; Smith, D.R.; Moreau, H.; Herron, M.D.; Verbruggen, H.; Delwiche, C.F.; De Clerck, O. Phylogeny and molecular evolution of the green algae. Crit. Rev. Plant. Sci. 2012, 31, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Efthimiou, G.; Mertzanis, A.; Emmanouloudis, D. Direct and indirect human-made impact on the natural ecosystems of the River Nestos. In Proceedings of the First International Conference on Environmental Research and Assessment, Ars Docenti, Romania, 23–27 March 2003; pp. 23–27. [Google Scholar]
- Dafis, S.; Papastergiadou, E.; Georgiou, K.; Babalonas, D.; Georgiadis, T.; Papageorgiou, M.; Lazaridou, T.; Tsiaousi, V. (Eds.) Directive 92/43/EC Works Biotopes in Greece: Network Natura 2000; Gouladri Museum of Natural History—Hellenic Center of Biotopes and Wetlands, EKBY: Thessaloniki, Greece, 1997. [Google Scholar]
- Pavlikakis, G.E.; Tsihrintzis, V.A. Integrating humans in ecosystem management using multi-criteria decision making. J. Am. Water Resour. Assoc. 2003, 39, 277–288. [Google Scholar] [CrossRef]
- Municipality of Komotini Official Website. Available online: https://www.komotini.gr/perivalon/fysiko-perivallon (accessed on 3 March 2023). (In Greek).
- NaturaGraeca, Nestos Delta and Lakes Vistonida-Ismarida. Available online: https://www.naturagraeca.com/ws/119,181,385,2,1,Nestos-Delta-and-lakes-Vistonida-Ismarida (accessed on 3 March 2023).
- Guillard, R.R.L.; Hargraves, P.E. Stichochrysis immobilis is a diatom, not a chrysophyte. Phycologia 1993, 32, 234–236. [Google Scholar] [CrossRef]
- Andersen, R.A.; Kawachi, M. Traditional Microalgae Isolation Techniques. In Algal Culturing Techniques; Andersen, R.A., Ed.; Elservier Academic Press: Oxford, UK, 2005; pp. 90–92. [Google Scholar]
- Norris, R.E.; Hori, T.; Chihara, M. Revision of the genus Tetraselmis (class Prasinophyceae). Bot. Mag. 1980, 93, 317–339. [Google Scholar] [CrossRef]
- Hori, T.; Norris, R.E.; Chihara, M. Studies on the ultrastructure and taxonomy of the genus Tetraselmis (Prasinophyceae). II: Subgenus Prasinocladia. Bot. Mag. 1983, 96, 385–392. [Google Scholar] [CrossRef]
- Hillebrand, H.; Dürselen, C.D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Olmos-Soto, J.; Paniagua-Michel, J.; Contreras, R.; Ochoa, L. DNA Fingerprinting Intron-Sizing Method to Accomplish a Specific, Rapid, and Sensitive Identification of Carotenogenic Dunaliella Species. In Microbial Carotenoids from Bacteria and Microalgae; Barredo, J.L., Ed.; Humana Press: Totowa, NJ, USA, 2012; Volume 892, pp. 269–281. [Google Scholar] [CrossRef]
- Nozaki, H.; Itoh, M.; Sano, R.; Uchida, H.; Watanabe, M.M.; Kuroiwa, T. Phylogenetic relationships within the colonial Volvocales (Chlorophyta) inferred from rbcL gene sequence data. J. Phycol. 1995, 31, 970–979. [Google Scholar] [CrossRef]
- Assunção, P.; Jaén-Molina, R.; Caujapé-Castells, J.; de la Jara, A.; Carmona, L.; Freijanes, K.; Mendoza, H. Phylogenetic position of Dunaliella acidophila (Chlorophyceae) based on ITS and rbcL sequences. J. Appl. Phycol. 2012, 24, 635–639. [Google Scholar] [CrossRef]
- Preetha, K.; John, L.; Subin, C.S.; Vijayan, K.K. Phenotypic and genetic characterization of Dunaliella (Chlorophyta) from Indian salinas and their diversity. Aquat. Biosyst. 2012, 8, 27. [Google Scholar] [CrossRef] [Green Version]
- Vieira, H.H.; Bagatini, I.L.; Guinart, C.M.; Vieira, A.A.H. TufA gene as molecular marker for freshwater Chlorophyceae. Algae 2016, 31, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, D67–D72. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; ISBN 0-19-513584-9. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Bremner, J.M. Total nitrogen. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties 9.2; Norman, A.G., Ed.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 1149–1178. [Google Scholar] [CrossRef]
- Olsen, M.F.L.; Pedersen, J.S.; Thomsen, S.T.; Martens, H.J.; Petersen, A.; Jensen, P.E. Outdoor cultivation of a novel isolate of the microalgae Scenedesmus sp. and the evaluation of its potential as a novel protein crop. Physiol. Plant. 2021, 173, 483–494. [Google Scholar] [CrossRef]
- Mishra, S.K.; Suh, W.I.; Farooq, W.; Moon, M.; Shrivastav, A.; Park, M.S.; Yang, J.W. Rapid quantification of microalgal lipids in aqueous medium by a simple colorimetric method. Biores. Tech. 2014, 155, 330–333. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Borovkov, A.B.; Gudvilovich, I.N.; Avsiyan, A.L.; Memetshaeva, N.O.A.; Lelekov, A.S.; Novikova, T.M. Production characteristics of Dunaliella salina at two-phase pilot cultivation (Crimea). Turk. J. Fish. Aquat. Sci. 2019, 20, 401–408. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Jan, S.; Khan, M.R.; Rashid, U.; Bokhari, J. Assessment of antioxidant potential, total phenolics and flavonoids of different solvent fractions of Monotheca buxifolia fruit. Osong Public Health Res. Perspect. 2013, 4, 246–254. [Google Scholar] [CrossRef] [Green Version]
- Safafar, H.; Van Wagenen, J.; Møller, P.; Jacobsen, C. Carotenoids, phenolic compounds and tocopherols contribute to the antioxidative properties of some microalgae species grown on industrial wastewater. Mar. Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benzie, I.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. In Methods in Enzymology; Packer, L., Ed.; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 15–27. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Guiry, M.D. AlgaeBase; World-Wide Electronic Publication, National University of Ireland: Galway, Ireland, 2013; Available online: https://www.algaebase.org (accessed on 9 March 2023).
- Levasseur, W.; Perré, P.; Pozzobon, V. A review of high value-added molecules production by microalgae in light of the classification. Biotechnol. Adv. 2020, 41, 107545. [Google Scholar] [CrossRef]
- Grubišić, M.; Šantek, B.; Zorić, Z.; Čošić, Z.; Vrana, I.; Gašparović, B.; Čož-Rakovac, R.; Ivančić Šantek, M. Bioprospecting of Microalgae Isolated from the Adriatic Sea: Characterization of Biomass, Pigment, Lipid and Fatty Acid Composition, and Antioxidant and Antimicrobial Activity. Molecules 2022, 27, 1248. [Google Scholar] [CrossRef]
- Barten, R.J.; Wijffels, R.H.; Barbosa, M.J. Bioprospecting and characterization of temperature tolerant microalgae from Bonaire. Algal Res. 2020, 50, 102008. [Google Scholar] [CrossRef]
- Ahmed, F.; Fanning, K.; Netzel, M.; Turner, W.; Li, Y.; Schenk, P.M. Profiling of carotenoids and antioxidant capacity of microalgae from subtropical coastal and brackish waters. Food Chem. 2014, 165, 300–306. [Google Scholar] [CrossRef] [Green Version]
- Fon-Sing, S.; Borowitzka, M.A. Isolation and screening of euryhaline Tetraselmis spp. suitable for large-scale outdoor culture in hypersaline media for biofuels. J. Appl. Phycol. 2016, 28, 1–14. [Google Scholar] [CrossRef]
- Lee, H.J.; Hur, S.B. Genetic relationships among multiple strains of the genus Tetraselmis based on partial 18S rDNA sequences. Algae 2009, 24, 205–212. [Google Scholar] [CrossRef]
- González, M.A.; Aguayo, P.A.; Inostroza, I.D.L.; Castro, P.A.; Fuentes, G.A.; Gómez, P.I. Ultrastructural and molecular characterization of Tetraselmis strains (Chlorodendrophyceae, Chlorophyta) isolated from Chile. Gayana Bot. 2015, 72, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Hori, T.; Norris, R.E.; Chihara, M. Studies on the ultrastructure and taxonomy of the genus Tetraselmis (Prasinophyceae). I: Subgenus Tetraselmis. Bot. Mag. 1982, 95, 49–61. [Google Scholar] [CrossRef]
- Hori, T.; Norris, R.E.; Chihara, M. Studies on the ultrastructure and taxonomy of the genus Tetraselmis (Prasinophyceae). III. Subgenus Parviselmis. Bot. Mag. 1986, 99, 123–135. [Google Scholar] [CrossRef]
- Fučíková, K.; Leliaert, F.; Cooper, E.D.; Škaloud, P.; D’hondt, S.; De Clerck, O.; Gurgel, C.F.D.; Lewis, L.A.; Lewis, P.O.; Lopez-Bautista, J.M.; et al. New phylogenetic hypotheses for the core Chlorophyta based on chloroplast sequence data. Front. Ecol. Evol. 2014, 2, 63. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Song, H.; Liu, X.; Liu, B.; Hu, Z.; Liu, G. Morphology and molecular phylogeny of coccoid green algae Coelastrella sensu lato (Scenedesmaceae, Sphaeropeales), including the description of three new species and two new varieties. J. Phycol. 2019, 55, 1290–1305. [Google Scholar] [CrossRef]
- Nieves, M.; Voltolina, D.; Piña, P. Growth and biomass production of Tetraselmis suecica and Dunaliella tertiolecta in a standard medium added with three products of zeolitic nature. Aquac. Eng. 2005, 32, 403–410. [Google Scholar] [CrossRef]
- Renaud, S.M.; Thinh, L.V.; Parry, D.L. The gross chemical composition and fatty acid composition of 18 species of tropical Australian microalgae for possible use in mariculture. Aquaculture 1999, 170, 147–159. [Google Scholar] [CrossRef]
- Meseck, S.L.; Alix, J.H.; Wikfors, G.H. Photoperiod and light intensity effects on growth and utilization of nutrients by the aquaculture feed microalga, Tetraselmis chui (LY429). Aquaculture 2005, 246, 393–404. [Google Scholar] [CrossRef]
- Lim, D.K.Y.; Garg, S.; Timmins, M.; Zhang, E.S.; Thomas-Hall, S.R.; Schuhmann, H.; Li, Y.; Schenk, P.M. Isolation and evaluation of oil-producing microalgae from subtropical coastal and brackish waters. PLoS ONE 2012, 7, e40751. [Google Scholar] [CrossRef] [PubMed]
- Fakhri, M.; Arifin, N.B.; Budianto, B.; Yuniarti, A.; Hariati, A.M. Effect of Salinity and Photoperiod on Growth of Microalgae Nannochloropsis sp. and Tetraselmis sp. Nat. Environ. Pollut. Technol. 2015, 14, 563. [Google Scholar]
- Arkronrat, W.; Deemark, P.; Oniam, V. Growth performance and proximate composition of mixed cultures of marine microalgae (Nannochloropsis sp. & Tetraselmis sp.) with monocultures. Songklanakarin J. Sci. Technol. 2016, 38, 1–5. [Google Scholar]
- Rafay, R.; Uratani, J.M.; Hernandez, H.H.; Rodríguez, J. Growth and Nitrate Uptake in Nannochloropsis gaditana and Tetraselmis chuii Cultures Grown in Sequential Batch Reactors. Front. Mar. Sci. 2020, 7, 77. [Google Scholar] [CrossRef] [Green Version]
- Moussa, I.D.B.; Chtourou, H.; Karray, F.; Sayadi, S.; Dhouib, A. Nitrogen or phosphorus repletion strategies for enhancing lipid or carotenoid production from Tetraselmis marina. Bioresour. Technol. 2017, 238, 325–332. [Google Scholar] [CrossRef]
- Kim, G.; Bae, J.; Lee, K. Nitrate repletion strategy for enhancing lipid production from marine microalga Tetraselmis sp. Bioresour. Technol. 2016, 205, 274–279. [Google Scholar] [CrossRef]
- Adarme-Vega, T.C.; Thomas-Hall, S.R.; Lim, D.K.; Schenk, P.M. Effects of long chain fatty acid synthesis and associated gene expression in microalga Tetraselmis sp. Mar. Drugs 2014, 12, 3381–3398. [Google Scholar] [CrossRef] [Green Version]
- Tsai, H.P.; Chuang, L.T.; Chen, C.N.N. Production of long chain omega-3 fatty acids and carotenoids in tropical areas by a new heat-tolerant microalga Tetraselmis sp. DS3. Food Chem. 2016, 192, 682–690. [Google Scholar] [CrossRef] [Green Version]
- Fabregas, J.; Herrero, C.; Cabezas, B.; Abalde, J. Mass culture and biochemical variability of the marine microalga Tetraselmis suecica Kylin (Butch) with high nutrient concentrations. Aquaculture 1985, 49, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Sigaud, T.C.S.; Aidar, E. Salinity and temperature effects on the growth and chlorophyll-α content of some planktonic algae. Bol. Inst. Oceanográfico 1993, 41, 95–103. [Google Scholar] [CrossRef]
- da Silva Gorgônio, C.M.; Aranda, D.A.G. Morphological and chemical aspects of Chlorella pyrenoidosa, Dunaliella tertiolecta, Is ochrysis galbana and Tetraselmis gracilis microalgae. Nat. Sci. 2013, 5, 783–791. [Google Scholar] [CrossRef] [Green Version]
- da Silva Ferreira, V.; Sant’Anna, C. Impact of culture conditions on the chlorophyll content of microalgae for biotechnological applications. World J. Microbiol. Biotechnol. 2017, 33, 20. [Google Scholar] [CrossRef] [PubMed]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; De Brabanter, J.; De Cooman, L. Antioxidant potential of microalgae in relation to their phenolic and carotenoid content. J. Appl. Phycol. 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Antioxidant activity of carotenoids. Mol. Aspects Med. 2003, 24, 345–351. [Google Scholar] [CrossRef]
- Young, A.J.; Lowe, G.L. Carotenoids—Antioxidant properties. Antioxidants 2018, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ravipati, A.S.; Koyyalamudi, S.R.; Jeong, S.C.; Reddy, N.; Smith, P.T.; Bartlett, J.; Shanmugam, K.; Münch, G.; Wu, M.J. Antioxidant and Anti-inflammatory Activities of Selected Medicinal Plants Containing Phenolic and Flavonoid Compounds. J. Agric. Food Chem. 2011, 59, 12361–12367. [Google Scholar] [CrossRef]
- Huyut, Z.; Beydemir, Ş.; Gülçin, İ. Antioxidant and Antiradical Properties of Selected Flavonoids and Phenolic Compounds. Biochem. Res. Int. 2017, 2017, 7616791. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Cheng, K.; Wong, C.; Fan, K.; Chen, F.; Jiang, Y. Evaluation of antioxidant capacity and total phenolic content of different fractions of selected microalgae. Food Chem. 2007, 102, 771–776. [Google Scholar] [CrossRef]
- Hajimahmoodi, M.; Faramarzi, M.A.; Mohammadi, N.; Soltani, N.; Oveisi, M.R.; Nafissi-Varcheh, N. Evaluation of antioxidant properties and total phenolic contents of some strains of microalgae. J. Appl. Phycol. 2010, 22, 43–50. [Google Scholar] [CrossRef]
- Bulut, O.; Akın, D.; Sönmez, Ç.; Öktem, A.; Yücel, M.; Öktem, H.A. Phenolic compounds, carotenoids, and antioxidant capacities of a thermo-tolerant Scenedesmus sp. (Chlorophyta) extracted with different solvents. J. Appl. Phycol. 2019, 31, 1675–1683. [Google Scholar] [CrossRef]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: An overview. Futur. J. Pharm. Sci. 2021, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Assunção, M.F.G.; Amaral, R.; Martins, C.B.; Ferreira, J.D.; Ressurreição, S.; Santos, S.D.; Varejão, J.M.; Santos, L.M. Screening microalgae as potential sources of antioxidants. J. Appl. Phycol. 2017, 29, 865–877. [Google Scholar] [CrossRef]
- Geetha, B.V.; Navasakthi, R.; Padmini, E. Investigation of antioxidant capacity and phytochemical composition of Sun Chlorella—An in vitro study. J. Aquac. Res. Dev. 2010, 1, 104. [Google Scholar] [CrossRef] [Green Version]
- Custódio, L.; Justo, T.; Silvestre, L.; Barradas, A.; Duarte, C.V.; Pereira, H.; Barreira, L.; Rauter, A.P.; Alberício, F.; Varela, J. Microalgae of different phyla display antioxidant, metal chelating and acetylcholinesterase inhibitory activities. Food Chem. 2012, 131, 134–140. [Google Scholar] [CrossRef]
- Goh, S.-H.; Yusoff, F.M.; Loh, S.-P. A comparison of the antioxidant properties and total phenolic content in a diatom, Chaetoceros sp. and a green microalga, Nannochloropsis sp. J. Agr. Sci. 2010, 2, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, M.; Santos, R.A.; Iglesias, P.; Couto, A.; Serra, C.R.; Gouvinhas, I.; Barros, A.; Oliva-Teles, A.; Enes, P.; Díaz-Rosales, P. Effect of extraction method and solvent system on the phenolic content and antioxidant activity of selected macro-and microalgae extracts. J. Appl. Phycol. 2020, 32, 349–362. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lagoon | SA (km2) | P (km) | MD (m) | T(°C) | CND (mS) | S (‰) | DO (mg·L−1) |
|---|---|---|---|---|---|---|---|
| Porto Lagos (PLA) | 13 | - | - | 23.7 | 8.04 | 64.8 | 49 |
| Ksirolimni (KSI) | 1.90 | 5.8 | 1.0 | 23.9 | 8.22 | 63.0 | 47 |
| Gene | Primer | Sequence | Reference |
|---|---|---|---|
| 18S | MA1 | 5′-CGGGATCCGTAGTCATATGCTTGTCTC-3′ | Olmos-Soto et al., 2012 [19] |
| MA2 | 5′-CGGAATTCCTTCTGCAGGTTCACC-3′ | ||
| DSs | 5′-GCAGGAGAGCTAATAGGA-3′ | ||
| rbcL | rbcL 475–497 | 5′-CGTGACAAACTAAACAAATATGG-3′ | Nozaki et al., 1995 [20] Assunção et al., 2012 [21] |
| rbcL 1181–1160 | 5′-AAGATTTCAACTAAAGCTGGCA-3′ | ||
| ITS region | ITS1 | 5′-TCCGTAGGTGAACCTGCGG-3′ | Preetha et al., 2012 [22] |
| ITS4 | 5′-TCCTCCGCTTAT-TGATATGC-3′ | ||
| tufA | TufA F | 5′-TGAAACAGAAMAWCGTCATTATGC-3′ | Vieira et al., 2016 [23] |
| TufA R | 5′-CCTTCNCGAATMGCRAAWCGC-3′ |
| Genetic Locus | Strain Code | Accession Number | Score (Bits) | Identities (ID) | % ID | Gaps | Strand | Scientific Name |
|---|---|---|---|---|---|---|---|---|
| ITS | PLA 1-2 | KP100529.1 | 1101 | 599/601 | 99.67 | 0 | ++ | Tetraselmis sp. DS3 |
| KSI 1-3 | 1101 | 599/601 | 99.67 | 0 | ++ | |||
| rbcL | PLA 1-2 | KX904699.1 | 1042 | 600/618 | 97.09 | 0 | ++ | Tetraselmis rubens V 2_2 |
| KSI 1-3 | 941 | 541/558 | 96.95 | 0 | ++ | |||
| tufA | PLA 1-2 | ON645926.1 | 1149 | 824/924 | 89.18 | 3/924 | ++ | Tetraselmis marina |
| KSI 1-3 | 1171 | 838/939 | 89.24 | 3/939 | +− | |||
| 18S | KSI 1-3 data | OQ220342.1 | 2974 | 1615/1618 | 99.81 | 1/1618 | ++ | Tetraselmis marina BEA0158B |
| KX904704.1 | 2916 | 1584/1587 | 99.81 | 1/1587 | ++ | Tetraselmis rubens R 9_3 | ||
| KX904703.1 | 2913 | 1582/1585 | 99.81 | 1/1585 | ++ | Tetraselmis rubens R 9_2 | ||
| KT860871.1 | 2876 | 1562/1565 | 99.81 | 1/1565 | ++ | Tetraselmis rubens RCC133 |
| Strain | Chl a (mg/g DW) | Chl b (mg/g DW) | TC (mg/g DW) | TP (% of DW) | TL (% of DW) | PS (% of DW) |
|---|---|---|---|---|---|---|
| PLA 1-2 | 7.58 ** ± 0.54 | 5.73 ** ± 0.48 | 3.25 * ± 0.25 | 15.76 * ± 1.09 | 16.37 * ± 0.98 | 40.94 ** ± 1.71 |
| KSI 1-3 | 5.16 ** ± 0.17 | 3.46 ** ± 0.15 | 3.71 * ± 0.12 | 28.60 * ± 1.35 | 12.77 * ± 1.25 | 27.76 ** ± 0.03 |
| Strain | TPC (mg Gallic Acid/g DW) | TFC (mg Quercetin/g DW) | FRAP (mg Ascorbic Acid/g DW) | TEAC (mg TROLOX/g DW) |
|---|---|---|---|---|
| PLA 1-2 | 5.99 ± 0.09 | 1.34 ** ± 0.11 | 1.42 ** ± 0.02 | 3.73 ** ± 0.37 |
| KSI 1-3 | 5.84 ± 0.05 | 3.77 ** ± 0.32 | 1.70 ** ± 0.03 | 5.49 ** ± 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntzouvaras, A.; Chantzistrountsiou, X.; Papageorgiou, N.; Koletti, A.; Adamakis, I.-D.; Zografaki, M.-E.; Marka, S.; Vasilakis, G.; Tsirigoti, A.; Tzovenis, I.; et al. New Records of Tetraselmis sp. Strains with Biotechnological Potential Isolated from Greek Coastal Lagoons. Water 2023, 15, 1698. https://doi.org/10.3390/w15091698
Ntzouvaras A, Chantzistrountsiou X, Papageorgiou N, Koletti A, Adamakis I-D, Zografaki M-E, Marka S, Vasilakis G, Tsirigoti A, Tzovenis I, et al. New Records of Tetraselmis sp. Strains with Biotechnological Potential Isolated from Greek Coastal Lagoons. Water. 2023; 15(9):1698. https://doi.org/10.3390/w15091698
Chicago/Turabian StyleNtzouvaras, Alexandros, Xanthi Chantzistrountsiou, Niki Papageorgiou, Aikaterini Koletti, Ioannis-Dimosthenis Adamakis, Maria-Eleftheria Zografaki, Sofia Marka, Gabriel Vasilakis, Amerssa Tsirigoti, Ioannis Tzovenis, and et al. 2023. "New Records of Tetraselmis sp. Strains with Biotechnological Potential Isolated from Greek Coastal Lagoons" Water 15, no. 9: 1698. https://doi.org/10.3390/w15091698
APA StyleNtzouvaras, A., Chantzistrountsiou, X., Papageorgiou, N., Koletti, A., Adamakis, I. -D., Zografaki, M. -E., Marka, S., Vasilakis, G., Tsirigoti, A., Tzovenis, I., Flemetakis, E., & Economou-Amilli, A. (2023). New Records of Tetraselmis sp. Strains with Biotechnological Potential Isolated from Greek Coastal Lagoons. Water, 15(9), 1698. https://doi.org/10.3390/w15091698