
Patterns and Predictors of Phytoplankton Assemblage Structure in a Coastal Lagoon: Species-Specific Analysis Needed to Disentangle Anthropogenic Pressures from Ocean Processes



Abstract
:1. Introduction
2. Materials and Methods
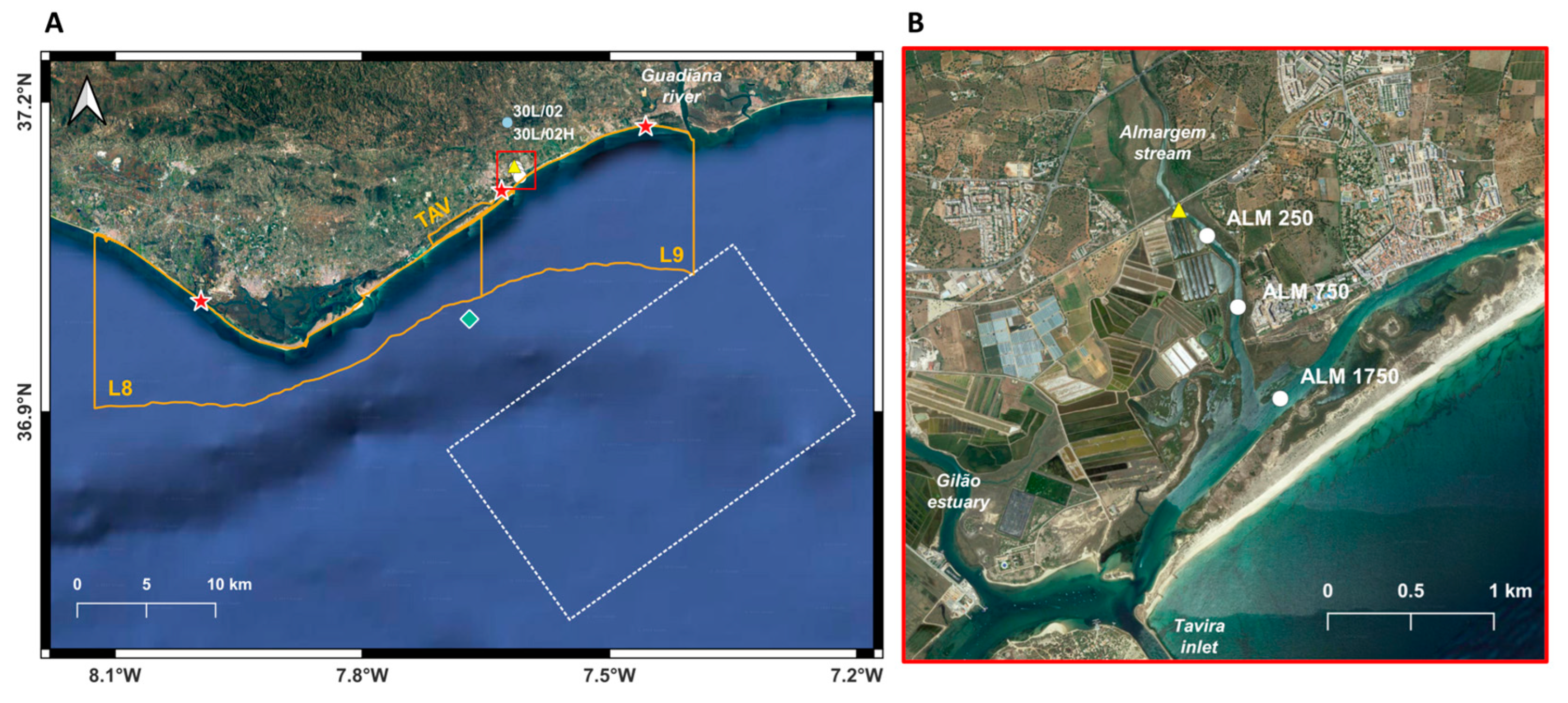
2.1. Study Area
2.2. Sampling Strategy
2.3. Physico-Chemical, Meteorological, and Hydrographic Variables
2.4. Oceanographic Setting at the Adjacent Coastal Area
2.5. Phytoplankton Data
2.5.1. Chlorophyll-a Concentration
2.5.2. Phytoplankton Abundance and Composition
2.6. Data and Statistical Analysis
2.6.1. Univariate Analysis of Phytoplankton and Environmental Data
2.6.2. Multivariate Analysis of Phytoplankton and Environmental Data
3. Results
3.1. Abiotic Environmental Setting
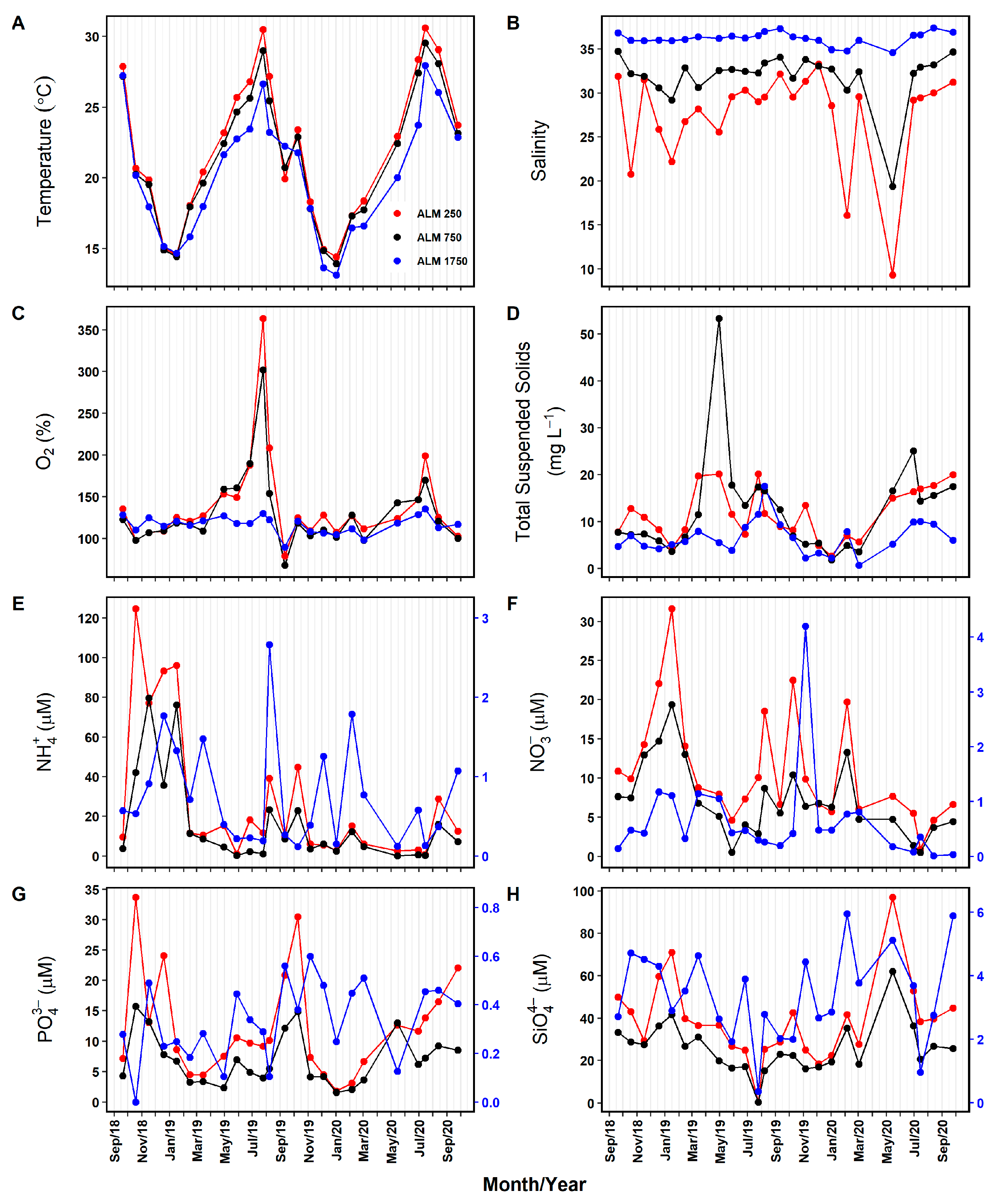
3.1.1. External Forcings: Meteorological, Hydrological, and Oceanographic Variables
3.1.2. Physico-Chemical Conditions in the Ria Formosa Lagoon
3.2. Phytoplankton Assemblage Structure
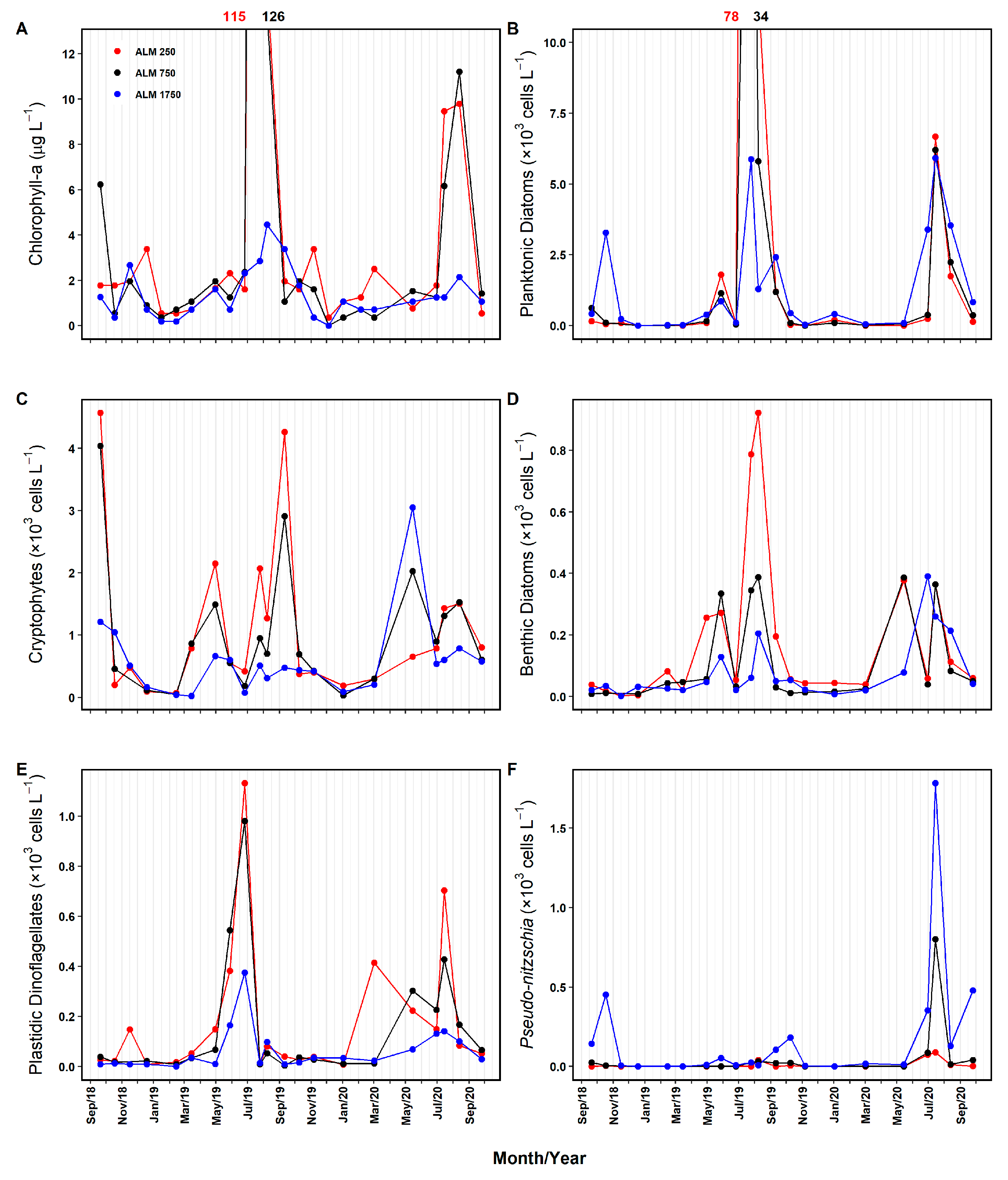
3.2.1. Phytoplankton Biomass, Abundance, and Diversity
3.2.2. Phytoplankton Assemblage Structure and Main Taxa Contributing to Dissimilarity
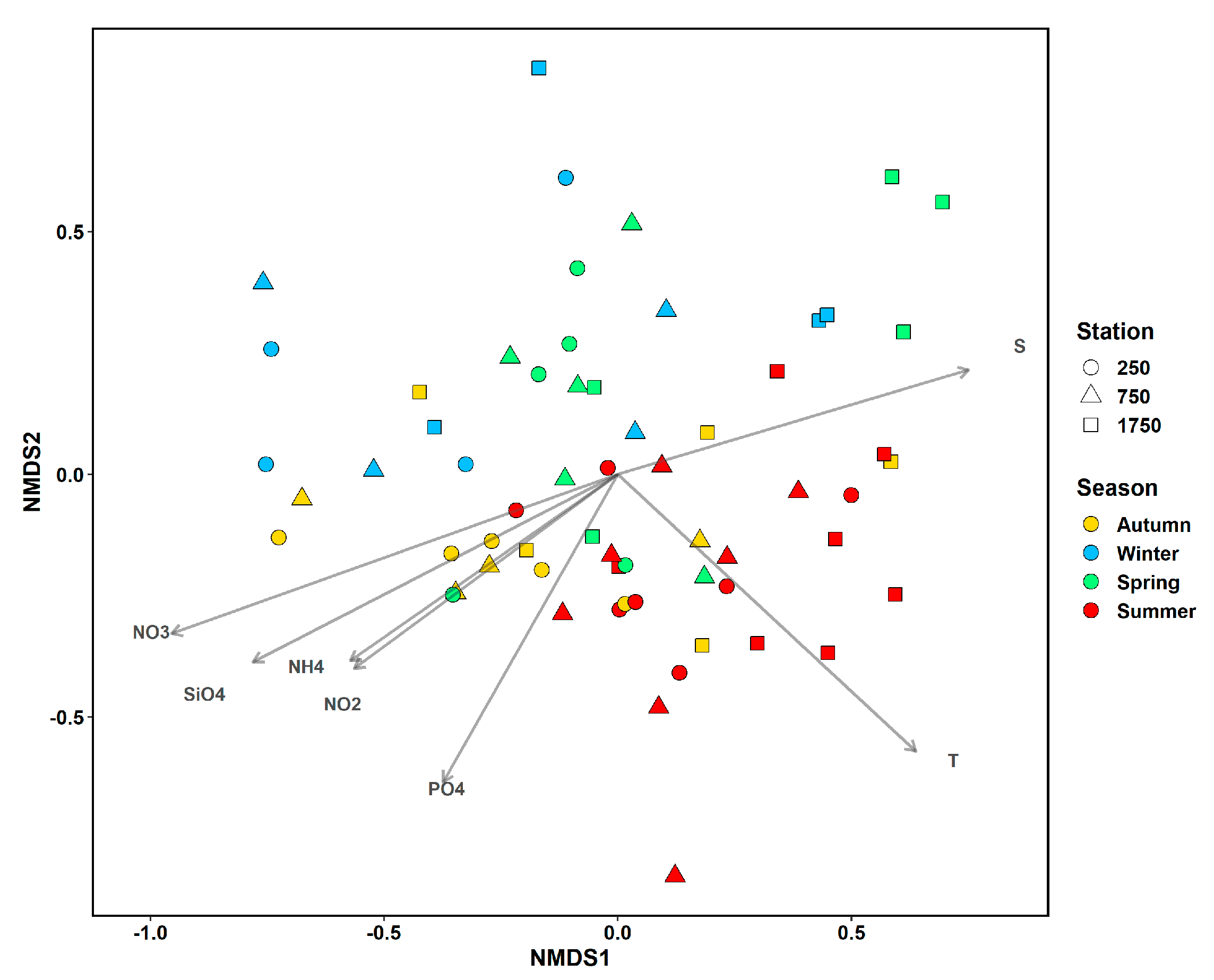
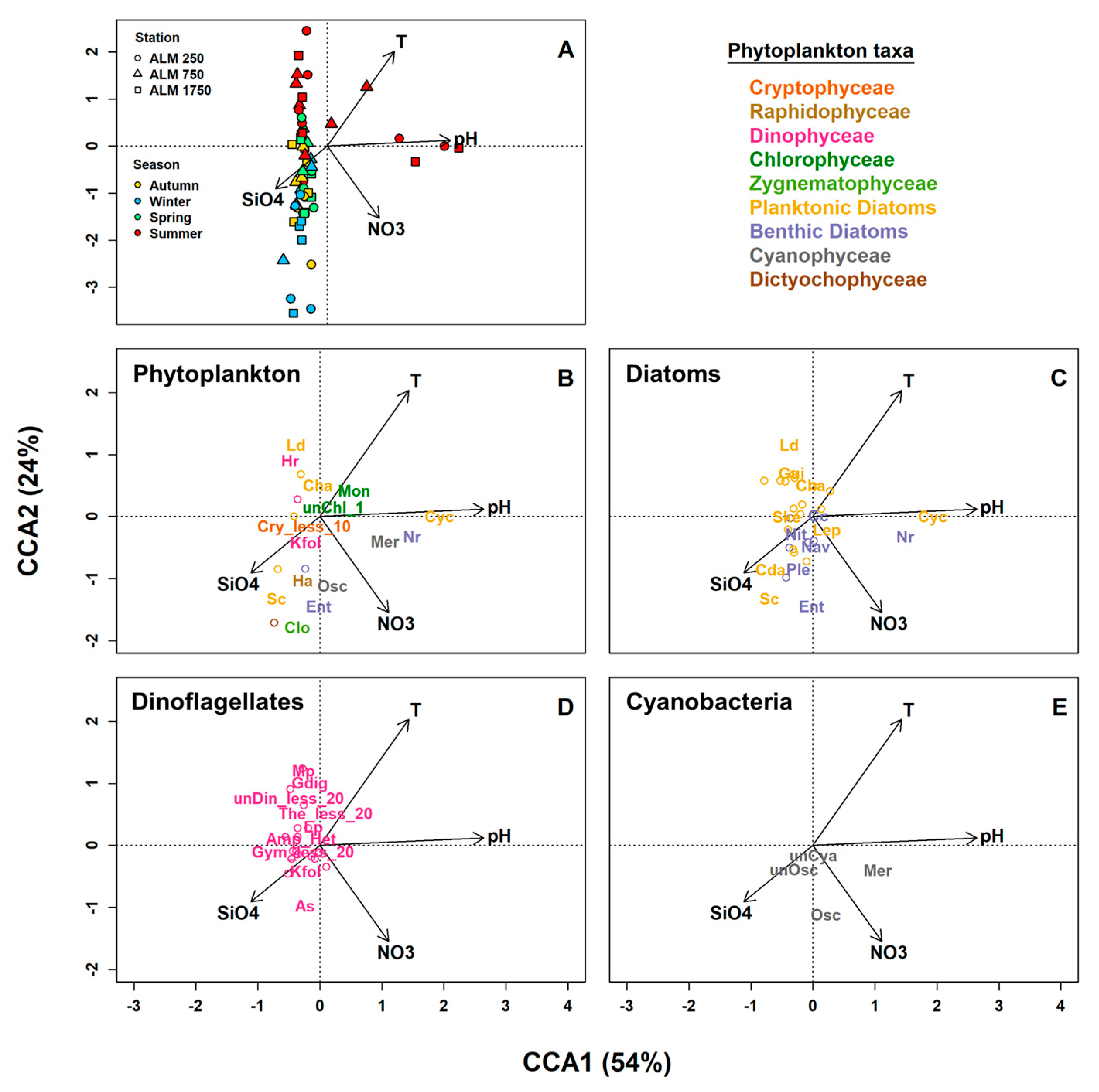
3.3. Linkages between Phytoplankton Assemblage Structure and Abiotic Environmental Variables
4. Discussion
4.1. Abiotic Environmental Setting
4.2. Composition of Phytoplankton Assemblages
4.3. Spatial–Temporal Variability in Phytoplankton Biomass and Abundance
4.4. Spatial–Temporal Variability in the Structure of Phytoplankton Assemblages
4.5. Linkages between the Structure of Phytoplankton Assemblages and Environmental Variables
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Boyd, P.W.; Claustre, H.; Levy, M.; Siegel, D.A.; Weber, T. Multi-Faceted Particle Pumps Drive Carbon Sequestration in the Ocean. Nature 2019, 568, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Glibert, P.M. Margalef Revisited: A New Phytoplankton Mandala Incorporating Twelve Dimensions, Including Nutritional Physiology. Harmful Algae 2016, 55, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Weithoff, G.; Beisner, B.E. Measures and Approaches in Trait-Based Phytoplankton Community Ecology—From Freshwater to Marine Ecosystems. Front. Mar. Sci. 2019, 6, 40. [Google Scholar] [CrossRef]
- Shumway, S.E.; Burkholder, J.M.; Morton, S.L. Harmful Algal Blooms: A Compendium Desk Reference, 1st ed.; Shumway, S.E., Burkholder, J.M., Morton, S.L., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2018; ISBN 9781118994658. [Google Scholar]
- Legendre, L.; Rassoulzadegan, F. Plankton and Nutrient Dynamics in Marine Waters. Ophelia 1995, 41, 153–172. [Google Scholar] [CrossRef]
- Cloern, J.E.; Dufford, R. Phytoplankton Community Ecology: Principles Applied in San Francisco Bay. Mar. Ecol. Prog. Ser. 2005, 285, 11–28. [Google Scholar] [CrossRef]
- Romagnan, J.-B.; Legendre, L.; Guidi, L.; Jamet, J.-L.; Jamet, D.; Mousseau, L.; Pedrotti, M.-L.; Picheral, M.; Gorsky, G.; Sardet, C.; et al. Comprehensive Model of Annual Plankton Succession Based on the Whole-Plankton Time Series Approach. PLoS ONE 2015, 10, e0119219. [Google Scholar] [CrossRef]
- Facca, C.; Aubry, F.B.; Socal, G.; Ponis, E.; Acri, F.; Bianchi, F.; Giovanardi, F.; Sfriso, A. Description of a Multimetric Phytoplankton Index (MPI) for the Assessment of Transitional Waters. Mar. Pollut. Bull. 2014, 79, 145–154. [Google Scholar] [CrossRef]
- Hemraj, D.A.; Hossain, M.A.; Ye, Q.; Qin, J.G.; Leterme, S.C. Plankton Bioindicators of Environmental Conditions in Coastal Lagoons. Estuar. Coast. Shelf Sci. 2017, 184, 102–114. [Google Scholar] [CrossRef]
- McQuatters-Gollop, A.; Johns, D.G.; Bresnan, E.; Skinner, J.; Rombouts, I.; Stern, R.; Aubert, A.; Johansen, M.; Bedford, J.; Knights, A. From Microscope to Management: The Critical Value of Plankton Taxonomy to Marine Policy and Biodiversity Conservation. Mar. Policy 2017, 83, 1–10. [Google Scholar] [CrossRef]
- Tweddle, J.F.; Gubbins, M.; Scott, B.E. Should Phytoplankton Be a Key Consideration for Marine Management? Mar. Policy 2018, 97, 1–9. [Google Scholar] [CrossRef]
- Dix, N.; Phlips, E.; Suscy, P. Factors Controlling Phytoplankton Biomass in a Subtropical Coastal Lagoon: Relative Scales of Influence. Estuaries Coasts 2013, 36, 981–996. [Google Scholar] [CrossRef]
- Kennish, M.J.; Paerl, H.W. Coastal Lagoons: Critical Habitats of Environmental Change, 1st ed.; Kennish, M.J., Paerl, H.W., Eds.; CRC Press: Boca Raton, FL, USA, 2010; ISBN 9780429143595. [Google Scholar]
- Newton, A.; Brito, A.C.; Icely, J.D.; Derolez, V.; Clara, I.; Angus, S.; Schernewski, G.; Inácio, M.; Lillebø, A.I.; Sousa, A.I.; et al. Assessing, Quantifying and Valuing the Ecosystem Services of Coastal Lagoons. J. Nat. Conserv. 2018, 44, 50–65. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Pérez-Ruzafa, I.M.; Newton, A.; Marcos, C. Coastal Lagoons: Environmental Variability, Ecosystem Complexity, and Goods and Services Uniformity. In Coasts and Estuaries: The Future; Wolanski, E., Day, J.W., Elliott, M., Ramachandran, R., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 253–276. ISBN 9780128140048. [Google Scholar]
- Newton, A.; Icely, J.; Cristina, S.; Perillo, G.M.E.; Turner, R.E.; Ashan, D.; Cragg, S.; Luo, Y.; Tu, C.; Li, Y.; et al. Anthropogenic, Direct Pressures on Coastal Wetlands. Front. Ecol. Evol. 2020, 8, 144. [Google Scholar] [CrossRef]
- Ramdani, M.; Elkhiati, N.; Flower, R.J.; Thompson, J.R.; Chouba, L.; Kraiem, M.M.; Ayache, F.; Ahmed, M.H. Environmental Influences on the Qualitative and Quantitative Composition of Phytoplankton and Zooplankton in North African Coastal Lagoons. Hydrobiologia 2009, 622, 113–131. [Google Scholar] [CrossRef]
- Tarafdar, L.; Kim, J.Y.; Srichandan, S.; Mohapatra, M.; Muduli, P.R.; Kumar, A.; Mishra, D.R.; Rastogi, G. Responses of Phytoplankton Community Structure and Association to Variability in Environmental Drivers in a Tropical Coastal Lagoon. Sci. Total Environ. 2021, 783, 146873. [Google Scholar] [CrossRef]
- Law, I.K.; Hii, K.S.; Lau, W.L.S.; Leaw, C.P.; Lim, P.T. Coastal Micro-Phytoplankton Community Changes during the Toxigenic Alexandrium Minutum Blooms in a Semi-Enclosed Tropical Coastal Lagoon (Malaysia, South China Sea). Reg. Stud. Mar. Sci. 2023, 57, 102733. [Google Scholar] [CrossRef]
- Hemraj, D.A.; Hossain, A.; Ye, Q.; Qin, J.G.; Leterme, S.C. Anthropogenic Shift of Planktonic Food Web Structure in a Coastal Lagoon by Freshwater Flow Regulation. Sci. Rep. 2017, 7, 44441. [Google Scholar] [CrossRef]
- Collos, Y.; Bec, B.; Jauzein, C.; Abadie, E.; Laugier, T.; Lautier, J.; Pastoureaud, A.; Souchu, P.; Vaquer, A. Oligotrophication and Emergence of Picocyanobacteria and a Toxic Dinoflagellate in Thau Lagoon, Southern France. J. Sea Res. 2009, 61, 68–75. [Google Scholar] [CrossRef]
- Collos, Y.; Jauzein, C.; Ratmaya, W.; Souchu, P.; Abadie, E.; Vaquer, A. Comparing Diatom and Alexandrium Catenella/Tamarense Blooms in Thau Lagoon: Importance of Dissolved Organic Nitrogen in Seasonally N-Limited Systems. Harmful Algae 2014, 37, 84–91. [Google Scholar] [CrossRef]
- Mercado, J.M.; Cortés, D.; Gómez-Jakobsen, F.; García-Gómez, C.; Ouaissa, S.; Yebra, L.; Ferrera, I.; Valcárcel-Pérez, N.; López, M.; García-Muñoz, R.; et al. Role of Small-Sized Phytoplankton in Triggering an Ecosystem Disruptive Algal Bloom in a Mediterranean Hypersaline Coastal Lagoon. Mar. Pollut. Bull. 2021, 164, 111989. [Google Scholar] [CrossRef] [PubMed]
- Poot-Delgado, C.A.; Okolodkov, Y.B.; Aké-Castillo, J.A.; Rendón-von Osten, J. Annual Cycle of Phytoplankton with Emphasis on Potentially Harmful Species in Oyster Beds of Términos Lagoon, Southeastern Gulf of Mexico. Rev. Biol. Mar. Oceanogr. 2015, 50, 465–477. [Google Scholar] [CrossRef]
- Fantasia, R.L.; Bricelj, V.M.; Ren, L. Phytoplankton Community Structure Based on Photopigment Markers in a Mid-Atlantic U.S. Coastal Lagoon: Significance for Hard-Clam Production. J. Coast. Res. 2017, 78, 106–126. [Google Scholar] [CrossRef]
- Phlips, E.J.; Badylak, S.; Lasi, M.A.; Chamberlain, R.; Green, W.C.; Hall, L.M.; Hart, J.A.; Lockwood, J.C.; Miller, J.D.; Morris, L.J.; et al. From Red Tides to Green and Brown Tides: Bloom Dynamics in a Restricted Subtropical Lagoon Under Shifting Climatic Conditions. Estuaries Coasts 2015, 38, 886–904. [Google Scholar] [CrossRef]
- Phlips, E.J.; Badylak, S.; Nelson, N.G.; Havens, K.E. Hurricanes, El Niño and Harmful Algal Blooms in Two Sub-Tropical Florida Estuaries: Direct and Indirect Impacts. Sci. Rep. 2020, 10, 1910. [Google Scholar] [CrossRef]
- Haraguchi, L.; Carstensen, J.; Abreu, P.C.; Odebrecht, C. Long-Term Changes of the Phytoplankton Community and Biomass in the Subtropical Shallow Patos Lagoon Estuary, Brazil. Estuar. Coast. Shelf Sci. 2015, 162, 76–87. [Google Scholar] [CrossRef]
- Cravo, A.; Barbosa, A.B.; Correia, C.; Matos, A.; Caetano, S.; Lima, M.J.; Jacob, J. Unravelling the Effects of Treated Wastewater Discharges on the Water Quality in a Coastal Lagoon System (Ria Formosa, South Portugal): Relevance of Hydrodynamic Conditions. Mar. Pollut. Bull. 2022, 174, 113296. [Google Scholar] [CrossRef]
- Barbosa, A.B.; Chícharo, M.A. Hydrology and Biota Interactions as Driving Forces for Ecosystem Functioning. In Treatise on Estuarine and Coastal Science; Wolanski, E., McLusky, D.S., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2011; Volume 10, pp. 7–47. ISBN 9780080878850. [Google Scholar]
- Pérez-Ruzafa, A.; Marcos, C.; Pérez-Ruzafa, I.M.; Pérez-Marcos, M. Coastal Lagoons: “Transitional Ecosystems” between Transitional and Coastal Waters. J. Coast. Conserv. 2011, 15, 369–392. [Google Scholar] [CrossRef]
- Cloern, J.E.; Jassby, A.D. Complex Seasonal Patterns of Primary Producers at the Land-Sea Interface. Ecol. Lett. 2008, 11, 1294–1303. [Google Scholar] [CrossRef]
- Cloern, J.E.; Jassby, A.D. Patterns and Scales of Phytoplankton Variability in Estuarine-Coastal Ecosystems. Estuaries Coasts 2010, 33, 230–241. [Google Scholar] [CrossRef]
- Carstensen, J.; Klais, R.; Cloern, J.E. Phytoplankton Blooms in Estuarine and Coastal Waters: Seasonal Patterns and Key Species. Estuar. Coast. Shelf Sci. 2015, 162, 98–109. [Google Scholar] [CrossRef]
- Ligorini, V.; Crayol, E.; Huneau, F.; Garel, E.; Malet, N.; Garrido, M.; Simon, L.; Cecchi, P.; Pasqualini, V. Small Mediterranean Coastal Lagoons Under Threat: Hydro-Ecological Disturbances and Local Anthropogenic Pressures (Size Matters). Estuaries Coasts 2023, 46, 2220–2243. [Google Scholar] [CrossRef] [PubMed]
- Oseji, O.F.; Chigbu, P.; Oghenekaro, E.; Waguespack, Y.; Chen, N. Spatiotemporal Patterns of Phytoplankton Composition and Abundance in the Maryland Coastal Bays: The Influence of Freshwater Discharge and Anthropogenic Activities. Estuar. Coast. Shelf Sci. 2018, 207, 119–131. [Google Scholar] [CrossRef]
- Stefanidou, N.; Katsiapi, M.; Tsianis, D.; Demertzioglou, M.; Michaloudi, E.; Moustaka-Gouni, M. Patterns in Alpha and Beta Phytoplankton Diversity along a Conductivity Gradient in Coastal Mediterranean Lagoons. Diversity 2020, 12, 38. [Google Scholar] [CrossRef]
- Hart, J.A.; Phlips, E.J.; Badylak, S.; Dix, N.; Petrinec, K.; Mathews, A.L.; Green, W.; Srifa, A. Phytoplankton Biomass and Composition in a Well-Flushed, Sub-Tropical Estuary: The Contrasting Effects of Hydrology, Nutrient Loads and Allochthonous Influences. Mar. Environ. Res. 2015, 112, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Abreu, P.C.; Marangoni, J.; Odebrecht, C. So Close, so Far: Differences in Long-Term Chlorophyll a Variability in Three Nearby Estuarine-Coastal Stations. Mar. Biol. Res. 2017, 13, 9–21. [Google Scholar] [CrossRef]
- Aubry, F.B.; Acri, F.; Bastianini, M.; Finotto, S.; Pugnetti, A. Differences and Similarities in the Phytoplankton Communities of Two Coupled Transitional and Marine Ecosystems (the Lagoon of Venice and the Gulf of Venice—Northern Adriatic Sea). Front. Mar. Sci. 2022, 9, 974967. [Google Scholar] [CrossRef]
- Barbosa, A.B. Seasonal and Interannual Variability of Planktonic Microbes in a Mesotidal Coastal Lagoon (Ria Formosa, SE Portugal): Impact of Climatic Changes and Local Human Influences. In Coastal Lagoons: Critical Habitats of Environmental Change; Kennish, M.J., Paerl, H.W., Eds.; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Oxfordshire, UK, 2010; pp. 335–366. ISBN 978-1-4200-8830-4. [Google Scholar]
- Cravo, A.; Cardeira, S.; Pereira, C.; Rosa, M.; Alcântara, P.; Madureira, M.; Rita, F.; Luis, J.; Jacob, J. Exchanges of Nutrients and Chlorophyll a through Two Inlets of Ria Formosa, South of Portugal, during Coastal Upwelling Events. J. Sea Res. 2014, 93, 63–74. [Google Scholar] [CrossRef]
- Cravo, A.; Rosa, A.; Jacob, J.; Correia, C. Dissolved Oxygen Dynamics in Ria Formosa Lagoon (South Portugal)—A Real Time Monitoring Station Observatory. Mar. Chem. 2020, 223, 103806. [Google Scholar] [CrossRef]
- Newton, A.; Icely, J.; Cristina, S.; Brito, A.; Cardoso, A.C.; Colijn, F.; Riva, S.D.; Gertz, F.; Hansen, J.W.; Holmer, M.; et al. An Overview of Ecological Status, Vulnerability and Future Perspectives of European Large Shallow, Semi-Enclosed Coastal Systems, Lagoons and Transitional Waters. Estuar. Coast. Shelf Sci. 2014, 140, 95–122. [Google Scholar] [CrossRef]
- Newton, A.; Cañedo-Argüelles, M.; March, D.; Goela, P.; Cristina, S.; Zacarias, M.; Icely, J. Assessing the Effectiveness of Management Measures in the Ria Formosa Coastal Lagoon, Portugal. Front. Ecol. Evol. 2022, 10, 508218. [Google Scholar] [CrossRef]
- Cravo, A.; Fernandes, D.; Damião, T.; Pereira, C.; Reis, M.P. Determining the Footprint of Sewage Discharges in a Coastal Lagoon in South-Western Europe. Mar. Pollut. Bull. 2015, 96, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Veríssimo, F.; Martins, F.; Janeiro, J. The Role of Ria Formosa as a Waste Water Receiver. In Ria Formosa: Challenges of a Coastal Lagoon in a Changing Environment; Aníbal, J., Gomes, A., Mendes, I., Moura, D., Eds.; Universidade do Algarve Editora: Faro, Portugal, 2019; pp. 47–66. [Google Scholar]
- Arias, P.A.; Bellouin, N.; Coppola, E.; Jones, R.G.; Krinner, G.; Marotzke, J.; Naik, V.; Palmer, M.D.; Plattner, G.-K.; Rogelj, J.; et al. Technical Summary. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; pp. 33–144. ISBN 9781009157896. [Google Scholar]
- Cravo, A.; Cardeira, S.; Pereira, C.; Rosa, M.; Alcântara, P.; Madureira, M.; Rita, F.; Correia, C.; Rosa, A.; Jacob, J. Nutrients and Chlorophyll-a Exchanges through an Inlet of the Ria Formosa Lagoon, SW Iberia during the Productive Season—Unravelling the Role of the Driving Forces. J. Sea Res. 2019, 144, 133–141. [Google Scholar] [CrossRef]
- Rosa, A.; Cravo, A.; Jacob, J.; Correia, C. Water Quality of a Southwest Iberian Coastal Lagoon: Spatial and Temporal Variability. Cont. Shelf Res. 2022, 245, 104804. [Google Scholar] [CrossRef]
- Morais, P.; Chícharo, M.A.; Barbosa, A. Phytoplankton Dynamics in a Coastal Saline Lake (SE-Portugal). Acta Oecologica 2003, 24, 87–96. [Google Scholar] [CrossRef]
- Loureiro, S.; Newton, A.; Icely, J. Boundary Conditions for the European Water Framework Directive in the Ria Formosa Lagoon, Portugal (Physico-Chemical and Phytoplankton Quality Elements). Estuar. Coast. Shelf Sci. 2006, 67, 382–398. [Google Scholar] [CrossRef]
- Barbosa, A.B. Estrutura e Dinâmica Da Teia Alimentar Microbiana Na Ria Formosa; University of Algarve: Faro, Portugal, 2006. [Google Scholar]
- Pereira, M.G.; Icely, J.; Mudge, S.; Newton, A.; Pereira, M.G.; Icely, J.; Mudge, S.; Newton, A.; Icely, J.; Mudge, S.; et al. Temporal and Spatial Variation of Phytoplankton Pigments in the Western Part of Ria Formosa Lagoon, Southern Portugal. Environ. Forensics 2007, 8, 205–220. [Google Scholar] [CrossRef]
- Jacob, J.; Correia, C.; Torres, A.F.; Xufre, G.; Matos, A.; Ferreira, C.; Reis, M.P.; Caetano, S.; Freitas, C.S.; Barbosa, A.B.; et al. Impacts of Decommissioning and Upgrading Urban Wastewater Treatment Plants on the Water Quality in a Shellfish Farming Coastal Lagoon (Ria Formosa, South Portugal). J. Coast. Res. 2020, 95, 45–50. [Google Scholar] [CrossRef]
- Loureiro, S.; Newton, A.; Icely, J. Effects of Nutrient Enrichments on Primary Production in the Ria Formosa Coastal Lagoon (Southern Portugal). Hydrobiologia 2005, 550, 29–45. [Google Scholar] [CrossRef]
- Domingues, R.B. Seasonal and Spatial Variability of Phytoplankton Primary Production in a Shallow Temperate Coastal Lagoon (Ria Formosa, Portugal). Plants 2022, 11, 3511. [Google Scholar] [CrossRef]
- Brito, A.C.; Quental, T.; Coutinho, T.P.; Branco, M.A.C.; Falcão, M.; Newton, A.; Icely, J.; Moita, T. Phytoplankton Dynamics in Southern Portuguese Coastal Lagoons during a Discontinuous Period of 40 Years: An Overview. Estuar. Coast. Shelf Sci. 2012, 110, 147–156. [Google Scholar] [CrossRef]
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in Shellfish from Two Portuguese Transitional Water Bodies Suggests the Marine Dinoflagellate Gymnodinium Catenatum as a Potential BMAA Source. Aquat. Toxicol. 2014, 152, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Domingues, R.B.; Lima, M.J. Unusual Red Tide of the Dinoflagellate Lingulodinium Polyedra during an Upwelling Event off the Algarve Coast (SW Iberia). Reg. Stud. Mar. Sci. 2023, 63, 102998. [Google Scholar] [CrossRef]
- Domingues, R.B.; Guerra, C.C.; Barbosa, A.B.; Brotas, V.; Galvão, H.M. Effects of Ultraviolet Radiation and CO2 Increase on Winter Phytoplankton Assemblages in a Temperate Coastal Lagoon. J. Plankton Res. 2014, 36, 672–684. [Google Scholar] [CrossRef]
- Domingues, R.B.; Guerra, C.C.; Barbosa, A.B.; Galvão, H.M. Are Nutrients and Light Limiting Summer Phytoplankton in a Temperate Coastal Lagoon? Aquat. Ecol. 2015, 49, 127–146. [Google Scholar] [CrossRef]
- Domingues, R.B.; Guerra, C.C.; Galvão, H.M.; Brotas, V.; Barbosa, A.B. Short-Term Interactive Effects of Ultraviolet Radiation, Carbon Dioxide and Nutrient Enrichment on Phytoplankton in a Shallow Coastal Lagoon. Aquat. Ecol. 2017, 51, 91–105. [Google Scholar] [CrossRef]
- Domingues, R.B.; Guerra, C.C.; Barbosa, A.B.; Galvão, H.M. Will Nutrient and Light Limitation Prevent Eutrophication in an Anthropogenically-Impacted Coastal Lagoon? Cont. Shelf Res. 2017, 141, 11–25. [Google Scholar] [CrossRef]
- Domingues, R.B.; Barreto, M.; Brotas, V.; Galvão, H.M.; Barbosa, A.B. Short-Term Effects of Winter Warming and Acidification on Phytoplankton Growth and Mortality: More Losers than Winners in a Temperate Coastal Lagoon. Hydrobiologia 2021, 848, 4763–4785. [Google Scholar] [CrossRef]
- Domingues, R.B.; Nogueira, P.; Barbosa, A.B. Co-Limitation of Phytoplankton by N and P in a Shallow Coastal Lagoon (Ria Formosa): Implications for Eutrophication Evaluation. Estuaries Coasts 2023, 46, 1557–1572. [Google Scholar] [CrossRef]
- Smayda, T.J.; Trainer, V.L. Dinoflagellate Blooms in Upwelling Systems: Seeding, Variability, and Contrasts with Diatom Bloom Behaviour. Prog. Oceanogr. 2010, 85, 92–107. [Google Scholar] [CrossRef]
- Mudge, S.M.; Icely, J.D.; Newton, A. Residence Times in a Hypersaline Lagoon: Using Salinity as a Tracer. Estuar. Coast. Shelf Sci. 2008, 77, 278–284. [Google Scholar] [CrossRef]
- Serpa, D.; Jesus, D.; Falcão, M.; da Fonseca, L.C. Ria Formosa Ecosystem: Socioeconomic Approach; IPIMAR: Matosinhos, Portugal, 2005; Volume 28. [Google Scholar]
- Pólvora, S.; Aníbal, J.; Martins, A. Saline Intrusions at Almargem Waste Water Treatment Plant in Different Tidal Cycles. In INCREaSE 2019; Springer: Berlin/Heidelberg, Germany, 2020; pp. 786–798. [Google Scholar] [CrossRef]
- Grasshoff, K.; Kremling, K.; Ehrhardt, M. Determination of Nutrients. In Methods of Seawater Analysis; Wiley Blackwell: Hoboken, NJ, USA, 1999; pp. 159–228. ISBN 9783527613984. [Google Scholar]
- Direção Regional de Agricultura e Pescas Do Algarve. Available online: https://www.drapalgarve.gov.pt/pt/servicos-e-produtos/servicos/fitossanidade/avisos-agricolas (accessed on 5 November 2022).
- Sistema Nacional de Informação de Recursos Hídricos. Available online: http://snirh.apambiente.pt/ (accessed on 7 November 2022).
- NASA Ocean Color. Available online: http://oceancolor.gsfc.nasa.gov/ (accessed on 20 September 2022).
- Robinson, I.S. Discovering the Ocean from Space: The Unique Applications of Satellite Oceanography; Springer: Berlin/Heidelberg, Germany, 2010; ISBN 978-3-540-24430-1. [Google Scholar]
- Bograd, S.J.; Schroeder, I.; Sarkar, N.; Qiu, X.; Sydeman, W.J.; Schwing, F.B. Phenology of Coastal Upwelling in the California Current. Geophys. Res. Lett. 2009, 36, 1–5. [Google Scholar] [CrossRef]
- Blended Sea Winds. Available online: https://www.ncei.noaa.gov/products/blended-sea-winds (accessed on 20 September 2022).
- Zhang, H.M.; Bates, J.J.; Reynolds, R.W. Assessment of Composite Global Sampling: Sea Surface Wind Speed. Geophys. Res. Lett. 2006, 33, 17714. [Google Scholar] [CrossRef]
- Krug, L.A.; Platt, T.; Sathyendranath, S.; Barbosa, A.B. Unravelling Region-Specific Environmental Drivers of Phytoplankton across a Complex Marine Domain (off SW Iberia). Remote Sens. Environ. 2017, 203, 162–184. [Google Scholar] [CrossRef]
- Lorenzen, C.J. Determination of chlorophyll and pheo-pigments: Spectrophotometric equations. Limnol. Oceanogr. 1967, 12, 343–346. [Google Scholar] [CrossRef]
- OceanColour-CCI. Available online: https://www.oceancolour.org/ (accessed on 20 September 2022).
- Sathyendranath, S.; Brewin, R.J.W.; Brockmann, C.; Brotas, V.; Calton, B.; Chuprin, A.; Cipollini, P.; Couto, A.B.; Dingle, J.; Doerffer, R.; et al. An Ocean-Colour Time Series for Use in Climate Studies: The Experience of the Ocean-Colour Climate Change Initiative (OC-CCI). Sensors 2019, 19, 4285. [Google Scholar] [CrossRef]
- Krug, L.A.; Platt, T.; Sathyendranath, S.; Barbosa, A.B. Patterns and Drivers of Phytoplankton Phenology off SW Iberia: A Phenoregion Based Perspective. Prog. Oceanogr. 2018, 165, 233–256. [Google Scholar] [CrossRef]
- Utermöhl, H. Zur Vervollkommnung Der Quantitativen Phytoplankton-Methodik. SIL Commun. 1958, 9, 1–38. [Google Scholar] [CrossRef]
- Andersen, P.; Thröndsen, J. Estimating Cell Numbers. In Manual on Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; UNESCO Publishing: Paris, France, 2004; pp. 99–130. [Google Scholar]
- Dodge, J.D.; Hart-Jones, B. Marine Dinoflagellates of the British Isles; H.M. Stationery Office: London, UK, 1982. [Google Scholar]
- Tomas, C.R. Identifying Marine Phytoplankton; Academic Press: Miami, FL, USA, 1997. [Google Scholar]
- Kraberg, A.; Baumann, M.E.M.; Dürselen, C.D. Coastal Phytoplankton: Photo Guide for Northern European Seas; Wiltshire, K.H., Boersma, M., Eds.; Pfeil Verlag: Munich, Germany, 2010. [Google Scholar]
- John, D.M.; Whitton, B.A.; Brook, A.J. The Freshwater Algal Flora of the British Isles. An Identification Guide to Freshwater and Terrestrial Algae, 2nd ed.; Cambridge University Press: London, UK, 2011; ISBN 9781108478007. [Google Scholar]
- Wehr, J.; Sheath, R.; Kociolek, J.P. Freshreshwater Algae of North America: Ecology and Classification, 2nd ed.; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. Available online: https://www.algaebase.org (accessed on 15 March 2023).
- Lundholm, N.; Churro, C.; Escalera, L.; Fraga, S.; Hoppenrath, M.; Iwataki, M.; Larsen, J.; Mertens, K.; Moestrup, Ø.; Tillmann, U.; et al. IOC-UNESCO Taxonomic Reference List of Harmful Micro Algae. Available online: https://www.marinespecies.org/hab (accessed on 17 March 2023).
- Lima, M.J.; Relvas, P.; Barbosa, A.B. Variability Patterns and Phenology of Harmful Phytoplankton Blooms off Southern Portugal: Looking for Region-Specific Environmental Drivers and Predictors. Harmful Algae 2022, 116, 102254. [Google Scholar] [CrossRef]
- Instituto Português Do Mar e Da Atmosfera. Available online: http://www.ipma.pt/pt/index.html/ (accessed on 2 February 2023).
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.0.3; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Chao, A. Nonparametric Estimation of the Number of Classes in a Population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004; ISBN 0-632-05633-9. [Google Scholar]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package, Version 2.6-4. 2022. Available online: https://github.com/vegandevs/vegan (accessed on 5 October 2022).
- Kindt, R.; Coe, R. Tree Diversity Analysis. A Manual and Software for Common Statistical Methods for Ecological and Biodiversity Studies; World Agroforestry Centre: Nairobi, Kenya, 2005. [Google Scholar]
- Durbin, J. Incomplete Blocks in Ranking Experiments. Br. J. Psychol. Stat. Sect. 1951, 4, 85–90. [Google Scholar] [CrossRef]
- Cisneros, K.O.; Smit, A.J.; Laudien, J.; Schoeman, D.S. Complex, Dynamic Combination of Physical, Chemical and Nutritional Variables Controls Spatio-Temporal Variation of Sandy Beach Community Structure. PLoS ONE 2011, 6, e23724. [Google Scholar] [CrossRef]
- Rose, V.; Rollwagen-Bollens, G.; Bollens, S.M.; Zimmerman, J. Seasonal and Interannual Variation in Lower Columbia River Phytoplankton (2005–2018): Environmental Variability and a Decline in Large Bloom-Forming Diatoms. Aquat. Microb. Ecol. 2021, 87, 29–46. [Google Scholar] [CrossRef]
- Goral, F.; Schellenberg, J. Goeveg: Functions for Community Data and Ordinations, Version 0.6.5. 2021. Available online: https://github.com/fvlampe/goeveg/ (accessed on 5 October 2022).
- Arbizu, P.M. PairwiseAdonis: Pairwise Multilevel Comparison Using Adonis, version 0.4. 2020. Available online: https://github.com/pmartinezarbizu/pairwiseAdonis (accessed on 5 October 2022).
- De Cáceres, M.; Legendre, P. Associations between Species and Groups of Sites: Indices and Statistical Inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Paliy, O.; Shankar, V. Application of Multivariate Statistical Techniques in Microbial Ecology. Mol. Ecol. 2016, 25, 1032–1057. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Clarke, K.R. Non-Parametric Multivariate Analyses of Changes in Community Structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutation Tests for Univariate or Multivariate Analysis of Variance and Regression. Can. J. Fish. Aquat. Sci. 2001, 58, 626–639. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E, Ltd.: Plymouth, UK, 2006. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Aubry, F.B.; Acri, F.; Finotto, S.; Pugnetti, A. Phytoplankton Dynamics and Water Quality in the Venice Lagoon. Water 2021, 13, 2780. [Google Scholar] [CrossRef]
- Clarke, K.R.; Ainsworth, M. A Method of Linking Multivariate Community Structure to Environmental Variables. Mar. Ecol. Prog. Ser. 1993, 92, 205–219. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Verdonschot, P.F.M. Canonical Correspondence Analysis and Related Multivariate Methods in Aquatic Ecology. Aquat. Sci. 1995, 57, 255–289. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R, 1st ed.; Springer: New York, NY, USA, 2009; ISBN 978-0-387-87457-9. [Google Scholar]
- Roselli, L.; Fabbrocini, A.; Manzo, C.; D’Adamo, R. Hydrological Heterogeneity, Nutrient Dynamics and Water Quality of a Non-Tidal Lentic Ecosystem (Lesina Lagoon, Italy). Estuar. Coast. Shelf Sci. 2009, 84, 539–552. [Google Scholar] [CrossRef]
- Caetano, S.; Correia, C.; Vidal, A.F.T.; Matos, A.; Ferreira, C.; Cravo, A. Fate of Microbial Contamination in a South European Coastal Lagoon (Ria Formosa) under the Influence of Treated Effluents Dispersal. J. Appl. Microbiol. 2023, 134, lxad166. [Google Scholar] [CrossRef]
- CCDRA. Plano Regional de Ordenamento Do Território. Anexo H—Recursos Hídricos, Planeamento e Gestão Do Recurso Água; CCDRA: San Diego, CA, USA, 2004. [Google Scholar]
- Malta, E.; Stigter, T.Y.; Pacheco, A.; Dill, A.C.; Tavares, D.; Santos, R. Effects of External Nutrient Sources and Extreme Weather Events on the Nutrient Budget of a Southern European Coastal Lagoon. Estuaries Coasts 2017, 40, 419–436. [Google Scholar] [CrossRef]
- Phlips, E.J.; Badylak, S.; Nelson, N.G.; Hall, L.M.; Jacoby, C.A.; Lasi, M.A.; Lockwood, J.C.; Miller, J.D. Cyclical Patterns and a Regime Shift in the Character of Phytoplankton Blooms in a Restricted Sub-Tropical Lagoon, Indian River Lagoon, Florida, United States. Front. Mar. Sci. 2021, 8, 730934. [Google Scholar] [CrossRef]
- Trombetta, T.; Vidussi, F.; Mas, S.; Parin, D.; Simier, M.; Mostajir, B. Water Temperature Drives Phytoplankton Blooms in Coastal Waters. PLoS ONE 2019, 14, e0214933. [Google Scholar] [CrossRef]
- Gilabert, J. Seasonal Plankton Dynamics in a Mediterranean Hypersaline Coastal Lagoon: The Mar Menor. J. Plankton Res. 2001, 23, 207–218. [Google Scholar] [CrossRef]
- Klaveness, D. Biology and Ecology of the Cryptophyceae: Status and Challenges. Biol. Oceanogr. 1989, 6, 257–270. [Google Scholar]
- Johnson, M.D.; Beaudoin, D.J.; Frada, M.J.; Brownlee, E.F.; Stoecker, D.K. High Grazing Rates on Cryptophyte Algae in Chesapeake Bay. Front. Mar. Sci. 2018, 5, 89–101. [Google Scholar] [CrossRef]
- Altenburger, A.; Blossom, H.E.; Garcia-Cuetos, L.; Jakobsen, H.H.; Carstensen, J.; Lundholm, N.; Hansen, P.J.; Moestrup; Haraguchi, L. Dimorphism in Cryptophytes—The Case of Teleaulax Amphioxeia/Plagioselmis Prolonga and Its Ecological Implications. Sci. Adv. 2020, 6, eabb1611. [Google Scholar] [CrossRef]
- Reynolds, C.S. Ecology of Phytoplankton: Ecology, Biodiversity and Conservation; Cambridge University Press: Cambridge, UK, 2006; ISBN 9780511542145. [Google Scholar]
- Brito, A.C.; Fernandes, T.F.; Newton, A.; Facca, C.; Tett, P. Does Microphytobenthos Resuspension Influence Phytoplankton in Shallow Systems? A Comparison through a Fourier Series Analysis. Estuar. Coast. Shelf Sci. 2012, 110, 77–84. [Google Scholar] [CrossRef]
- Derolez, V.; Soudant, D.; Malet, N.; Chiantella, C.; Richard, M.; Abadie, E.; Aliaume, C.; Bec, B. Two Decades of Oligotrophication: Evidence for a Phytoplankton Community Shift in the Coastal Lagoon of Thau (Mediterranean Sea, France). Estuar. Coast. Shelf Sci. 2020, 241, 106810. [Google Scholar] [CrossRef]
- Martínez-López, A.; Escobedo-Urías, D.; Reyes-Salinas, A.; Hernández-Real, M.T. Phytoplankton Response to Nutrient Runoff in a Large Lagoon System in the Gulf of California. Hidrobiológica 2007, 17, 101–112. [Google Scholar]
- Su, H.-M.; Lin, H.-J.; Hung, J.-J. Effects of Tidal Flushing on Phytoplankton in a Eutrophic Tropical Lagoon in Taiwan. Estuar. Coast. Shelf Sci. 2004, 61, 739–750. [Google Scholar] [CrossRef]
- Bernardi Aubry, F.; Acri, F.; Bianchi, F.; Pugnetti, A. Looking for Patterns in the Phytoplankton Community of the Mediterranean Microtidal Venice Lagoon: Evidence from Ten Years of Observations. Sci. Mar. 2013, 77, 47–60. [Google Scholar] [CrossRef]
- Leruste, A.; Pasqualini, V.; Garrido, M.; Malet, N.; De Wit, R.; Bec, B. Physiological and Behavioral Responses of Phytoplankton Communities to Nutrient Availability in a Disturbed Mediterranean Coastal Lagoon. Estuar. Coast. Shelf Sci. 2019, 219, 176–188. [Google Scholar] [CrossRef]
- Smayda, T.J.; Reynolds, C.S. Community Assembly in Marine Phytoplankton: Application of Recent Models to Harmful Dinoflagellate Blooms. J. Plankton Res. 2001, 23, 447–461. [Google Scholar] [CrossRef]
- Phlips, E.J.; Badylak, S.; Christman, M.; Wolny, J.; Brame, J.; Garland, J.; Hall, L.; Hart, J.; Landsberg, J.; Lasi, M.; et al. Scales of Temporal and Spatial Variability in the Distribution of Harmful Algae Species in the Indian River Lagoon, Florida, USA. Harmful Algae 2011, 10, 277–290. [Google Scholar] [CrossRef]
- Kemp, A.E.S.; Villareal, T.A. The Case of the Diatoms and the Muddled Mandalas: Time to Recognize Diatom Adaptations to Stratified Waters. Prog. Oceanogr. 2018, 167, 138–149. [Google Scholar] [CrossRef]
- Varona-Cordero, F.; Gutiérrez-Mendieta, F.J.; Meave Del Castillo, M.E. Phytoplankton Assemblages in Two Compartmentalized Coastal Tropical Lagoons (Carretas-Pereyra and Chantuto-Panzacola, Mexico). J. Plankton Res. 2010, 32, 1283–1299. [Google Scholar] [CrossRef]
- Socal, G.; Bianchi, F.; Alberighi, L. Effects of Thermal Pollution and Nutrient Discharges on a Spring Phytoplankton Bloom in the Industrial Area of the Lagoon of Venice. Vie Milieu/Life Environ. 1999, 49, 19–31. [Google Scholar]
- Domingues, R.B.; Barbosa, A.B. Evaluating Underwater Light Availability for Phytoplankton: Mean Light Intensity in the Mixed Layer versus Attenuation Coefficient. Water 2023, 15, 2966. [Google Scholar] [CrossRef]
- Acri, F.; Aubry, F.B.; Berton, A.; Bianchi, F.; Boldrin, A.; Camatti, E.; Comaschi, A.; Rabitti, S.; Socal, G. Plankton Communities and Nutrients in the Venice Lagoon: Comparison between Current and Old Data. J. Mar. Syst. 2004, 51, 321–329. [Google Scholar] [CrossRef]
- Ouaissa, S.; Gómez-Jakobsen, F.; Yebra, L.; Ferrera, I.; Moreno-Ostos, E.; Belando, M.D.; Ruiz, J.M.; Mercado, J.M. Phytoplankton Dynamics in the Mar Menor, a Mediterranean Coastal Lagoon Strongly Impacted by Eutrophication. Mar. Pollut. Bull. 2023, 192, 115074. [Google Scholar] [CrossRef] [PubMed]
- Odebrecht, C.; Abreu, P.C.; Carstensen, J. Retention Time Generates Short-Term Phytoplankton Blooms in a Shallow Microtidal Subtropical Estuary. Estuar. Coast. Shelf Sci. 2015, 162, 35–44. [Google Scholar] [CrossRef]
- Millan-Nuñez, R.; Alvarez-Borrego, S.; Nelson, D.M. Effects of Physical Phenomena on the Distribution of Nutrients and Phytoplankton Productivity in a Coastal Lagoon. Estuar. Coast. Shelf Sci. 1982, 15, 317–335. [Google Scholar] [CrossRef]
- Souchu, P.; Vaquer, A.; Collos, Y.; Landrein, S.; Deslous-Paoli, J.M.; Bibent, B. Influence of Shellfish Farming Activities on the Biogeochemical Composition of the Water Column in Thau Lagoon. Mar. Ecol. Prog. Ser. 2001, 218, 141–152. [Google Scholar] [CrossRef]
- Pérez-Ruzafa, A.; Campillo, S.; Fernández-Palacios, J.M.; García-Lacunza, A.; García-Oliva, M.; Ibañez, H.; Navarro-Martínez, P.C.; Pérez-Marcos, M.; Pérez-Ruzafa, I.M.; Quispe-Becerra, J.I.; et al. Long-Term Dynamic in Nutrients, Chlorophyll a, and Water Quality Parameters in a Coastal Lagoon During a Process of Eutrophication for Decades, a Sudden Break and a Relatively Rapid Recovery. Front. Mar. Sci. 2019, 6, 26. [Google Scholar] [CrossRef]
- Cloern, J.E. Our Evolving Conceptual Model of the Coastal Eutrophication Problem. Mar. Ecol. Prog. Ser. 2001, 210, 223–253. [Google Scholar] [CrossRef]
- Smayda, T.J. Complexity in the Eutrophication–Harmful Algal Bloom Relationship, with Comment on the Importance of Grazing. Harmful Algae 2008, 8, 140–151. [Google Scholar] [CrossRef]
- Cloern, J.E.; Jassby, A.D. Drivers of Change in Estuarine-Coastal Ecosystems: Discoveries from Four Decades of Study in San Francisco Bay. Rev. Geophys. 2012, 50, 1–33. [Google Scholar] [CrossRef]
- Lara-Lara, J.R.; Alvarez-Borrego, S.; Small, L.F. Variability and Tidal Exchange of Ecological Properties in a Coastal Lagoon. Estuar. Coast. Mar. Sci. 1980, 11, 613–637. [Google Scholar] [CrossRef]
- Bernardi Aubry, F.; Acri, F. Phytoplankton Seasonality and Exchange at the Inlets of the Lagoon of Venice (July 2001–June 2002). J. Mar. Syst. 2004, 51, 65–76. [Google Scholar] [CrossRef]
- Cebrián, J.; Valiela, I. Seasonal Patterns in Phytoplankton Biomass in Coastal Ecosystems. J. Plankton Res. 1999, 21, 429–444. [Google Scholar] [CrossRef]
- Trombetta, T.; Vidussi, F.; Roques, C.; Mas, S.; Scotti, M.; Mostajir, B. Co-Occurrence Networks Reveal the Central Role of Temperature in Structuring the Plankton Community of the Thau Lagoon. Sci. Rep. 2021, 11, 17675. [Google Scholar] [CrossRef] [PubMed]
- Rissik, D.; Shon, E.H.; Newell, B.; Baird, M.E.; Suthers, I.M. Plankton Dynamics Due to Rainfall, Eutrophication, Dilution, Grazing and Assimilation in an Urbanized Coastal Lagoon. Estuar. Coast. Shelf Sci. 2009, 84, 99–107. [Google Scholar] [CrossRef]
- Cadée, G.C.; Hegeman, J. Primary Production of Phytoplankton in the Dutch Wadden Sea. Neth. J. Sea Res. 1974, 8, 240–259. [Google Scholar] [CrossRef]
- Bec, B.; Collos, Y.; Souchu, P.; Vaquer, A.; Lautier, J.; Fiandrino, A.; Benau, L.; Orsoni, V.; Laugier, T. Distribution of Picophytoplankton and Nanophytoplankton along an Anthropogenic Eutrophication Gradient in French Mediterranean Coastal Lagoons. Aquat. Microb. Ecol. 2011, 63, 29–45. [Google Scholar] [CrossRef]
- Ligorini, V.; Malet, N.; Garrido, M.; Derolez, V.; Amand, M.; Bec, B.; Cecchi, P.; Pasqualini, V. Phytoplankton Dynamics and Bloom Events in Oligotrophic Mediterranean Lagoons: Seasonal Patterns but Hazardous Trends. Hydrobiologia 2022, 849, 2353–2375. [Google Scholar] [CrossRef]
- Vidal, T.; Calado, A.J.; Moita, M.T.; Cunha, M.R. Phytoplankton Dynamics in Relation to Seasonal Variability and Upwelling and Relaxation Patterns at the Mouth of Ria de Aveiro (West Iberian Margin) over a Four-Year Period. PLoS ONE 2017, 12, e0177237. [Google Scholar] [CrossRef]
- Spatharis, S.; Lamprinou, V.; Meziti, A.; Kormas, K.A.; Danielidis, D.D.; Smeti, E.; Roelke, D.L.; Mancy, R.; Tsirtsis, G. Everything Is Not Everywhere: Can Marine Compartments Shape Phytoplankton Assemblages? Proc. R. Soc. B 2019, 286, 20191890. [Google Scholar] [CrossRef] [PubMed]
- Srichandan, S.; Kim, J.Y.; Bhadury, P.; Barik, S.K.; Muduli, P.R.; Samal, R.N.; Pattnaik, A.K.; Rastogi, G. Spatiotemporal Distribution and Composition of Phytoplankton Assemblages in a Coastal Tropical Lagoon: Chilika, India. Environ. Monit. Assess. 2015, 187, 47. [Google Scholar] [CrossRef] [PubMed]
- del Jiménez-Quiroz, M.C.; Cervantes-Duarte, R.; Funes-Rodríguez, R.; Barón-Campis, S.A.; de García-Romero, F.J.; Hernández-Trujillo, S.; Hernández-Becerril, D.U.; González-Armas, R.; Martell-Dubois, R.; Cerdeira-Estrada, S.; et al. Impact of “The Blob” and “El Niño” in the SW Baja California Peninsula: Plankton and Environmental Variability of Bahia Magdalena. Front. Mar. Sci. 2019, 6, 25. [Google Scholar] [CrossRef]
- Litchman, E.; Klausmeier, C.A.; Schofield, O.M.; Falkowski, P.G. The Role of Functional Traits and Trade-Offs in Structuring Phytoplankton Communities: Scaling from Cellular to Ecosystem Level. Ecol. Lett. 2007, 10, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Leruste, A.; Guilhaumon, F.; De Wit, R.; Malet, N.; Collos, Y.; Bec, B. Phytoplankton Strategies to Exploit Nutrients in Coastal Lagoons with Different Eutrophication Status during Re-Oligotrophication. Aquat. Microb. Ecol. 2019, 83, 131–146. [Google Scholar] [CrossRef]
- Mendes, C.R.B.; Odebrecht, C.; Tavano, V.M.; Abreu, P.C. Pigment-Based Chemotaxonomy of Phytoplankton in the Patos Lagoon Estuary (Brazil) and Adjacent Coast. Mar. Biol. Res. 2016, 13, 22–35. [Google Scholar] [CrossRef]
- Vybernaite-Lubiene, I.; Zilius, M.; Giordani, G.; Petkuviene, J.; Vaiciute, D.; Bukaveckas, P.A.; Bartoli, M. Effect of Algal Blooms on Retention of N, Si and P in Europe’s Largest Coastal Lagoon. Estuar. Coast. Shelf Sci. 2017, 194, 217–228. [Google Scholar] [CrossRef]
- Catherine, Q.; Susanna, W.; Isidora, E.-S.; Mark, H.; Aurélie, V.; Jean-François, H. A Review of Current Knowledge on Toxic Benthic Freshwater Cyanobacteria—Ecology, Toxin Production and Risk Management. Water Res. 2013, 47, 5464–5479. [Google Scholar] [CrossRef]
- Kempton, J.W.; Wolny, J.; Tengs, T.; Rizzo, P.; Morris, R.; Tunnell, J.; Scott, P.; Steidinger, K.; Hymel, S.N.; Lewitus, A.J. Kryptoperidinium Foliaceum Blooms in South Carolina: A Multi-Analytical Approach to Identification. Harmful Algae 2002, 1, 383–392. [Google Scholar] [CrossRef]
- Smayda, T.J. Turbulence, Watermass Stratification and Harmful Algal Blooms: An Alternative View and Frontal Zones as “Pelagic Seed Banks”. Harmful Algae 2002, 1, 95–112. [Google Scholar] [CrossRef]
- Facca, C.; Bilaničovà, D.; Pojana, G.; Sfriso, A.; Marcomini, A. Harmful Algae Records in Venice Lagoon and in Po River Delta (Northern Adriatic Sea, Italy). Sci. World J. 2014, 2014, 806032. [Google Scholar] [CrossRef] [PubMed]
- Stanca, E.; Parsons, M.L. Phytoplankton Diversity along Spatial and Temporal Gradients in the Florida Keys. J. Plankton Res. 2017, 39, 531–549. [Google Scholar] [CrossRef]
- Alves-de-Souza, C.; Benevides, T.S.; Santos, J.B.O.; Von Dassow, P.; Guillou, L.; Menezes, M. Does Environmental Heterogeneity Explain Temporal β Diversity of Small Eukaryotic Phytoplankton? Example from a Tropical Eutrophic Coastal Lagoon. J. Plankton Res. 2017, 39, 698–714. [Google Scholar] [CrossRef]
- Paerl, H.W.; Hall, N.S.; Peierls, B.L.; Rossignol, K.L.; Joyner, A.R. Hydrologic Variability and Its Control of Phytoplankton Community Structure and Function in Two Shallow, Coastal, Lagoonal Ecosystems: The Neuse and New River Estuaries, North Carolina, USA. Estuaries Coasts 2014, 37, 31–45. [Google Scholar] [CrossRef]
- Pulina, S.; Padedda, B.M.; Nicola, S.; Lugliè, A. The Dominance of Cyanobacteria in Mediterranean Hypereutrophic Lagoons: A Case Study of Cabras Lagoon (Sardinia, Italy). Sci. Mar. 2011, 75, 111–120. [Google Scholar] [CrossRef]
- Pulina, S.; Suikkanen, S.; Satta, C.T.; Mariani, M.A.; Padedda, B.M.; Virdis, T.; Caddeo, T.; Sechi, N.; Lugliè, A. Multiannual Phytoplankton Trends in Relation to Environmental Changes across Aquatic Domains: A Case Study from Sardinia (Mediterranean Sea). Off. J. Soc. Bot. Ital. 2016, 150, 660–670. [Google Scholar] [CrossRef]
- Pulina, S.; Satta, C.T.; Padedda, B.M.; Sechi, N.; Lugliè, A. Seasonal Variations of Phytoplankton Size Structure in Relation to Environmental Variables in Three Mediterranean Shallow Coastal Lagoons. Estuar. Coast. Shelf Sci. 2018, 212, 95–104. [Google Scholar] [CrossRef]
- Soria, J.; Caniego, G.; Hernández-Sáez, N.; Dominguez-Gomez, J.A.; Erena, M. Phytoplankton Distribution in Mar Menor Coastal Lagoon (SE Spain) during 2017. J. Mar. Sci. Eng. 2020, 8, 600. [Google Scholar] [CrossRef]
- Güzel, U.; Ongun Sevindik, T.; Uzun, A. The Spatiotemporal Responses of Phytoplankton to Environmental Variables in 7 Coastal Lagoons of Kızılırmak Delta (Samsun, Türkiye). Aquat. Sci. 2023, 85, 73. [Google Scholar] [CrossRef]
- Minicante, S.A.; Piredda, R.; Quero, G.M.; Finotto, S.; Bernardi Aubry, F.; Bastianini, M.; Pugnetti, A.; Zingone, A. Habitat Heterogeneity and Connectivity: Effects on the Planktonic Protist Community Structure at Two Adjacent Coastal Sites (the Lagoon and the Gulf of Venice, Northern Adriatic Sea, Italy) Revealed by Metabarcoding. Front. Microbiol. 2019, 10, 2736. [Google Scholar] [CrossRef]
- Aké-Castillo, J.A.; Vázquez, G. Phytoplankton Variation and Its Relation to Nutrients and Allochthonous Organic Matter in a Coastal Lagoon on the Gulf of Mexico. Estuar. Coast. Shelf Sci. 2008, 78, 705–714. [Google Scholar] [CrossRef]
- Corcoran, A.A.; Wolny, J.; Leone, E.; Ivey, J.; Murasko, S. Drivers of Phytoplankton Dynamics in Old Tampa Bay, FL (USA), a Subestuary Lagging in Ecosystem Recovery. Estuar. Coast. Shelf Sci. 2017, 185, 130–140. [Google Scholar] [CrossRef]
- Philippart, C.J.M.; Cadée, G.C.; van Raaphorst, W.; Riegman, R. Long-Term Phytoplankton-Nutrient Interactions in a Shallow Coastal Sea: Algal Community Structure, Nutrient Budgets, and Denitrification Potential. Limnol. Oceanogr. 2000, 45, 131–144. [Google Scholar] [CrossRef]
- Coesel, P.F.M. Poor Physiological Adaptation to Alkaline Culture Conditions in Closterium Acutum Var. Variabile, a Planktonic Desmid from Eutrophic Waters. Eur. J. Phycol. 1993, 28, 53–57. [Google Scholar] [CrossRef]
- Bernardi Aubry, F.; Berton, A.; Bastianini, M.; Socal, G.; Acri, F. Phytoplankton Succession in a Coastal Area of the NW Adriatic, over a 10-Year Sampling Period (1990–1999). Cont. Shelf Res. 2004, 24, 97–115. [Google Scholar] [CrossRef]
- Glibert, P.M.; Allen, J.I.; Bouwman, A.F.; Brown, C.W.; Flynn, K.J.; Lewitus, A.J.; Madden, C.J. Modeling of HABs and Eutrophication: Status, Advances, Challenges. J. Mar. Syst. 2010, 83, 262–275. [Google Scholar] [CrossRef]
- Rodrigues, M.; Rosa, A.; Cravo, A.; Jacob, J.; Fortunato, A.B. Effects of Climate Change and Anthropogenic Pressures in the Water Quality of a Coastal Lagoon (Ria Formosa, Portugal). Sci. Total Environ. 2021, 780, 146311. [Google Scholar] [CrossRef]
- Trombetta, T.; Vidussi, F.; Roques, C.; Scotti, M.; Mostajir, B. Marine Microbial Food Web Networks During Phytoplankton Bloom and Non-Bloom Periods: Warming Favors Smaller Organism Interactions and Intensifies Trophic Cascade. Front. Microbiol. 2020, 11, 502336. [Google Scholar] [CrossRef]
- Cereja, R.; Brotas, V.; Brito, A.C.; Rodrigues, M. Effects of Droughts, Sea Level Rise, and Increase in Outfall Discharges on Phytoplankton in a Temperate Estuary (Tagus Estuary, Portugal). Reg. Environ. Chang. 2023, 23, 111. [Google Scholar] [CrossRef]
- Lopez, C.B.; Tilney, C.L.; Muhlbach, E.; Bouchard, J.N.; Villac, M.C.; Henschen, K.L.; Markley, L.R.; Abbe, S.K.; Shankar, S.; Shea, C.P.; et al. High-Resolution Spatiotemporal Dynamics of Harmful Algae in the Indian River Lagoon (Florida)—A Case Study of Aureoumbra Lagunensis, Pyrodinium Bahamense, and Pseudo-Nitzschia. Front. Mar. Sci. 2021, 8, 769877. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Station (ALM) | Mean ± SD [% Contr.] | Min–Max | N | DbS |
|---|---|---|---|---|---|
| Chlorophyll-a concentration and phytoplankton abundance | |||||
| Chlorophyll-a (µg L−1) | 250 | 8.5 ± 25.4 | 0.5–115.2 | 20 | 250 > 1750 ** and 750 > 1750 * |
| 750 | 9.0 ± 28.6 | 0.4–126.5 | 19 | ||
| 1750 | 1.5 ± 1.1 | 0.2–4.5 | 19 | ||
| Total Abundance (×103 cells L−1) | 250 | 8.131 ± 21.826 | 0.147–98.733 | 20 | ns |
| 750 | 4.767 ± 8.805 | 0.132–38.151 | 19 | ||
| 1750 | 2.148 ± 2.180 | 0.130–7.600 | 19 | ||
| Planktonic Diatoms (×103 cells L−1) | 250 | 5.052 ± 17.522 [73.7] | bd–78.432 | 20 | ns |
| 750 | 2.626 ± 7.718 [63.6] | bd–33.580 | 19 | ||
| 1750 | 1.19 ± 1.87 [60.4] | bd–5.922 | 19 | ||
| Pseudo-nitzschia spp. (×103 cells L−1) | 250 | 0.011 ± 0.026 [0.2] | bd–0.089 | 20 | 250 < 1750 *** and 750 < 1750 * and 250 < 750 * |
| 750 | 0.056 ± 0.182 [1.3] | bd–0.801 | 19 | ||
| 1750 | 0.173 ± 0.416 [7.7] | bd–1.782 | 19 | ||
| Cryptophyceans (×103 cells L−1) | 250 | 1.092 ± 1.281 [16.8] | 0.069–4.566 | 20 | ns |
| 750 | 0.978 ± 1.033 [24.5] | 0.032–4.04 | 19 | ||
| 1750 | 0.579 ± 0.678 [25.1] | 0.016–3.049 | 19 | ||
| Plastidic Dinoflagellates (×103 cells L−1) | 250 | 0.184 ± 0.286 [2.7] | 0.008–1.132 | 20 | ns |
| 750 | 0.152 ± 0.254 [3.7] | 0.004–0.981 | 19 | ||
| 1750 | 0.057 ± 0.090 [2.7] | 0.0002–0.376 | 19 | ||
| Kryptoperidinium foliaceum (×103 cells L−1) | 250 | 0.075 ± 0.136 [1.1] | bd–0.410 | 20 | 250 > 1750 *** and 750 > 1750 *** |
| 750 | 0.089 ± 0.187 [2.1] | bd–0.711 | 19 | ||
| 1750 | 0.0002 ± 0.001 [0.01] | bd–0.005 | 19 | ||
| Benthic Diatoms (×103 cells L−1) | 250 | 0.18 ± 0.26 [2.7] | 0.003–0.92 | 20 | ns |
| 750 | 0.12 ± 0.15 [2.8] | 0.009–0.39 | 19 | ||
| 1750 | 0.06 ± 0.07 [3.5] | 0.002–0.260 | 19 | ||
| Cyanobacteria (×103 cells L−1) | 250 | 0.197 ± 0.24 [2.8] | bd–0.8190 | 20 | ns |
| 750 | 0.081 ± 0.1384 [1.9] | bd–0.5341 | 19 | ||
| 1750 | 0.0078 ± 0.0167 [0.5] | bd–0.0585 | 19 | ||
| Phytoplankton diversity metrics | |||||
| Estimated species richness (Observed Species richness) | 250 | 136.2 (93) | na | 21 | na |
| 750 | 132.7 (90) | na | 20 | ||
| 1750 | 152.6 (120) | na | 21 | ||
| Average observed species richness | 250 | 19.9 ± 6.4 | 8–34 | 21 | 250 < 1750 ** and 750 < 1750 * |
| 750 | 20.7 ± 6.1 | 10–32 | 20 | ||
| 1750 | 27.1 ± 8.9 | 11–41 | 21 | ||
| Species dominance (Hulburt index) | 250 | 69.3 ± 14.7 | 39.9–95.5 | 21 | ns |
| 750 | 66.2 ± 13.7 | 40.1–92.3 | 20 | ||
| 1750 | 63.6 ± 14.0 | 37.3–90.3 | 21 | ||
| Species diversity (Shannon–Wiener index) | 250 | 1.6 ± 0.4 | 0.6–2.2 | 21 | ns |
| 750 | 1.6 ± 0.4 | 0.6–24 | 20 | ||
| 1750 | 1.7 ± 0.4 | 0.8–2.5 | 21 | ||
| Species evenness (Pielou index) | 250 | 0.5 ± 0.2 | 0.2–0.8 | 21 | ns |
| 750 | 0.5 ± 0.1 | 0.2–0.8 | 20 | ||
| 1750 | 0.5 ± 0.1 | 0.2–0.8 | 21 | ||
| Phytoplankton OTUs | IV (%) | p-Val. | ||
|---|---|---|---|---|
| 250 | 750 | 1750 | ||
| Chaetoceros spp. ∆ | 14.2 | 22.5 | 36.8 | |
| Unidentified Chlorophyte 1 ▼ | 27.9 | 25.7 | 33.6 | |
| Unidentified Coccolithophores ● | 31.6 | 29.1 | 17.8 | |
| Unidentified Cryptophyceae >10 μm + | 28.3 | 33.4 | 31.8 | |
| Unidentified Cryptophyceae <10 μm + | 33.3 | 31.4 | 30.4 | ns |
| Dinophysis acuminata complex ○ | 0 | 0 | 38.1 | ** |
| Eutreptiella spp. ˆ | 25.8 | 34.3 | 15.7 | |
| Unidentified Gymnodiniales <20 μm ○ | 36.6 | 25.6 | 19.4 | |
| Heterocapsa spp. ○ | 5.2 | 2.0 | 40.2 | * |
| Kryptoperidinium foliaceum ○ | 47.4 | 41.1 | 0.5 | ** |
| Unidentified Oscillatoriales □ | 31.0 | 22.4 | 2.4 | ** |
| Unidentified Pennales >10 μm ∆ | 34.9 | 30.4 | 23.6 | |
| Prorocentrum cf. scutellum ○ | 0.1 | 0 | 37.1 | ** |
| Prorocentrum micans ○ | 0.8 | 0.2 | 52.4 | ** |
| Prorocentrum spp. ○ | 0.1 | 0 | 37.7 | ** |
| Prorocentrum triestinum ○ | 0 | 0.02 | 33.2 | ** |
| Pseudo-nitzschia delicatissima group ∆ | 7.8 | 21.1 | 35.3 | |
| Pseudo-nitzschia seriata group ∆ | 0.9 | 6.9 | 48.1 | ** |
| Scrippsiella spp. ○ | 1.7 | 2.2 | 55.8 | ** |
| Tripos fusus ○ | 0.1 | 0 | 37.2 | ** |
| Phytoplankton OTUs | IV (%) | p-Val. | |||
|---|---|---|---|---|---|
| Autumn | Winter | Spring | Summer | ||
| Akashiwo cf. sanguinea ○ | 33.1 | 0 | 0 | 1.0 | ** |
| Chaetoceros spp. ∆ | 6.5 | 3.6 | 30.4 | 39.1 | ** |
| Unidentified Chlorophyte 1 ▼ | 21.9 | 6.2 | 20.8 | 41.8 | |
| Closterium sp. ● | 0 | 41.7 | 0 | 0 | ** |
| Unidentified Cryptophyceae <10 μm + | 21.1 | 13.5 | 29.5 | 31.6 | |
| Cylindrotheca closterium ∆ | 7.8 | 21.4 | 18.4 | 36.1 | |
| Eutreptiella spp. ˆ | 37.1 | 2.9 | 14.4 | 27.8 | ** |
| Unidentified Gymnodiniales <20 μm ○ | 13.9 | 6.7 | 38.4 | 26.3 | |
| Kryptoperidinium foliaceum ○ | 7.9 | 13.9 | 45.7 | 8.7 | |
| Navicula spp. ∆ | 1.1 | 9.2 | 30.3 | 5.6 | ns |
| Unidentified Pennales >10 μm ∆ | 11.2 | 16.8 | 35.2 | 27.6 | |
| Unidentified Pennales <10 μm ∆ | 2.9 | 9.3 | 15.7 | 40.8 | ** |
| Pseudo-nitzschia delicatissima group ∆ | 14.1 | 1.4 | 7.0 | 48.7 | * |
| Sundstroemia setigera/S. pungens ∆ | 1.0 | 0.2 | 0 | 65.8 | ** |
| Thalassiosira sp. ∆ | 0.2 | 1.7 | 1.1 | 50.2 | ** |
| Unidentified thecate dinoflagellates >20 μm ○ | 0.4 | 2.3 | 30.5 | 11.2 | ns |
| Lagoonal System (Country) | Environmental Variables | Methods | Reference |
|---|---|---|---|
| I—Mediterranean Sea | |||
| Cabras (Italy) | DIN, DIN/P, P, S | CCA | [172] |
| P, S | RDA | [173] | |
| Calich, Santa Giusta and Corru S’Ittiri (Italy) | DIN, P, S, Secchi depth, Si, T | RDA | [174] |
| Diana and Urbino (France) | Rainfall, S, T, turbidity | CCA | [156] |
| Mar Menor (Spain) | Chla, Secchi depth, turbidity | PCA | [175] |
| NO3, S, Si, T | CCA | [24] | |
| Chla, DIN, DIN/P, Kd, P, Si, Si/P, T | NMDS-envfit | [141] | |
| Thau (France) | NO3, P, wind | PCA | [122] |
| Depth, light, pressure, S, Si, T, turbidity, wind | NMDS-envfit | [152] | |
| Ulu, Uzun, Tatlı, Gıcı, Liman, Cernek, and Karaboğaz (Turkey) | Light, NO3, pH, P, S, Si, T | RDA | [176] |
| Venice (Italy) | T | NMDS, CCA | [177] |
| II—Atlantic Ocean | |||
| Florida Bay (USA) | Light, T, waves | NMDS, CCA | [169] |
| Patos lagoon estuary (Brazil) | Freshwater discharge, NH4, P, S, Secchi depth, T | CCA | [29] |
| Chla, DIN, P, S, Si, T | CCA | [163] | |
| Rodrigo de Freitas (Brazil) | NH4, NO3, P, pH, precipitation, S, Secchi depth, Si, T, water stability | CCA | [170] |
| Sontecomapan (Mexico) | pH, S, Si, T | CCA | [178] |
| Tampa Bay (USA) | N, DIN/DIP, P, pH, S, Si, Si/N, T, visibility | PCA, NMDS, CCA | [179] |
| Wadden Sea (Denmark, Germany and The Netherlands) | DIN, DIN/P, P | PCA | [180] |
| Ria Formosa (Portugal) | T, pH, NO3, Si | NMDS-envfit, BIOENV, CCA | This study |
| III—Pacific Ocean | |||
| Bahia Magdalena (Mexico) | Chla, DO, NH3, NO2, NO3, P, S, Si, T, UI | CCoA, GAMs | [160] |
| Carretas-Pereyra and Chantuto-Panzacola (Mexico) | NH4, NO2, S, Si, T | PCA, CCA | [137] |
| IV—Indian Ocean | |||
| Chilika (India) | DO, light, N, P, pH, S, Si, T, transparency, turbidity | NMDS, CCA | [159] |
| DIN, DO, pH, S, T, transparency | GAMs, HMSC | [19] | |
| Coorong (South Australia) | DIN/P, DO, freshwater discharge, N, NH3, S, T, turbidity | RDA, BIOENV | [21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, M.J.; Barbosa, A.B.; Correia, C.; Matos, A.; Cravo, A. Patterns and Predictors of Phytoplankton Assemblage Structure in a Coastal Lagoon: Species-Specific Analysis Needed to Disentangle Anthropogenic Pressures from Ocean Processes. Water 2023, 15, 4238. https://doi.org/10.3390/w15244238
Lima MJ, Barbosa AB, Correia C, Matos A, Cravo A. Patterns and Predictors of Phytoplankton Assemblage Structure in a Coastal Lagoon: Species-Specific Analysis Needed to Disentangle Anthropogenic Pressures from Ocean Processes. Water. 2023; 15(24):4238. https://doi.org/10.3390/w15244238
Chicago/Turabian StyleLima, Maria João, Ana B. Barbosa, Cátia Correia, André Matos, and Alexandra Cravo. 2023. "Patterns and Predictors of Phytoplankton Assemblage Structure in a Coastal Lagoon: Species-Specific Analysis Needed to Disentangle Anthropogenic Pressures from Ocean Processes" Water 15, no. 24: 4238. https://doi.org/10.3390/w15244238
APA StyleLima, M. J., Barbosa, A. B., Correia, C., Matos, A., & Cravo, A. (2023). Patterns and Predictors of Phytoplankton Assemblage Structure in a Coastal Lagoon: Species-Specific Analysis Needed to Disentangle Anthropogenic Pressures from Ocean Processes. Water, 15(24), 4238. https://doi.org/10.3390/w15244238