


Trophic Positions of Sympatric Copepods across the Subpolar Front of the East Sea during Spring: A Stable Isotope Approach

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samplings
2.2. Zooplankton Community Structure and Stable Isotope Analysis
2.3. Statistical Analysis
3. Results
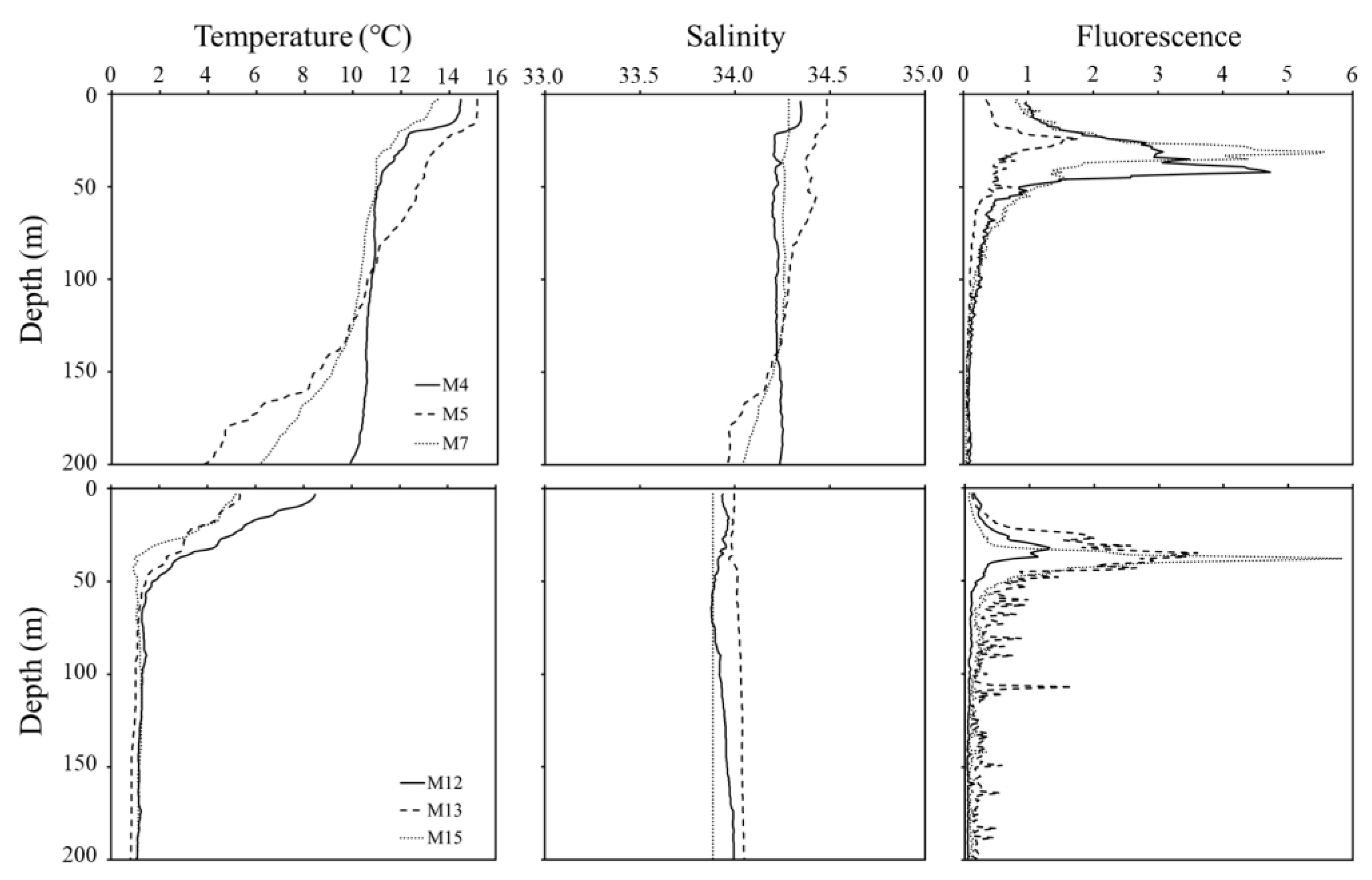
3.1. Temperature, Salinity, and Chlorophyll A Concentration
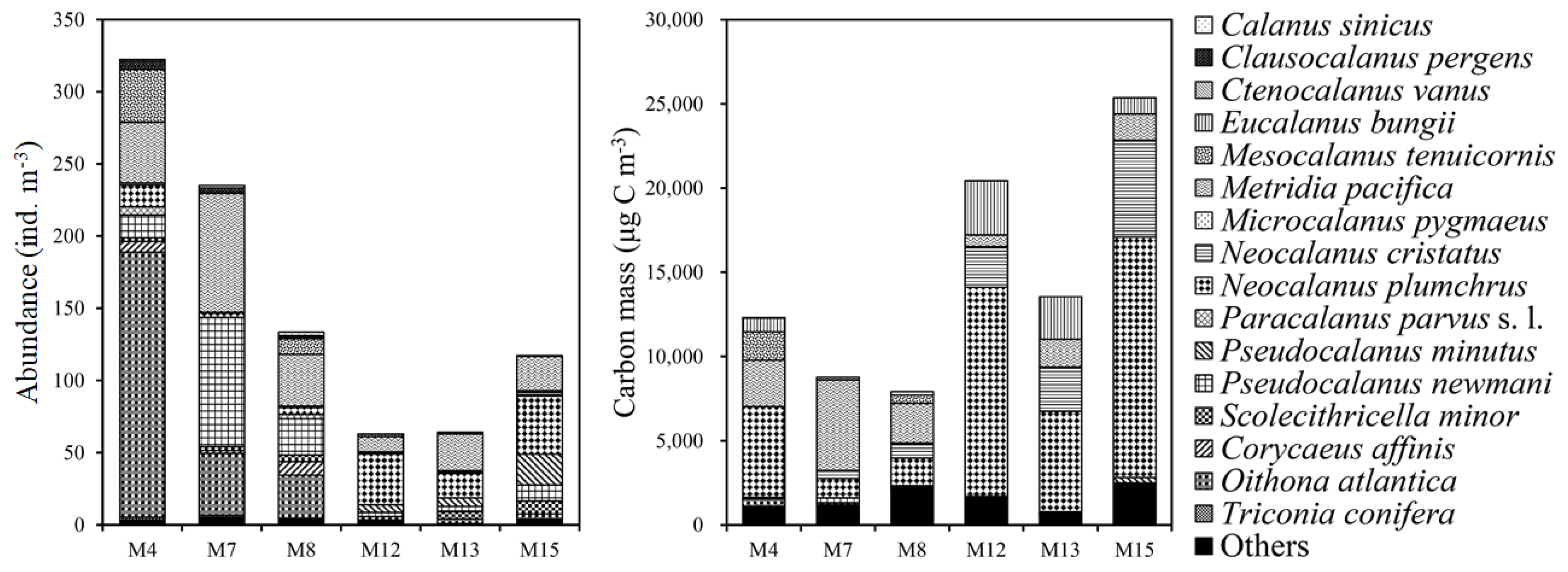
3.2. Copepod Abundance and Carbon Masses across the SPF
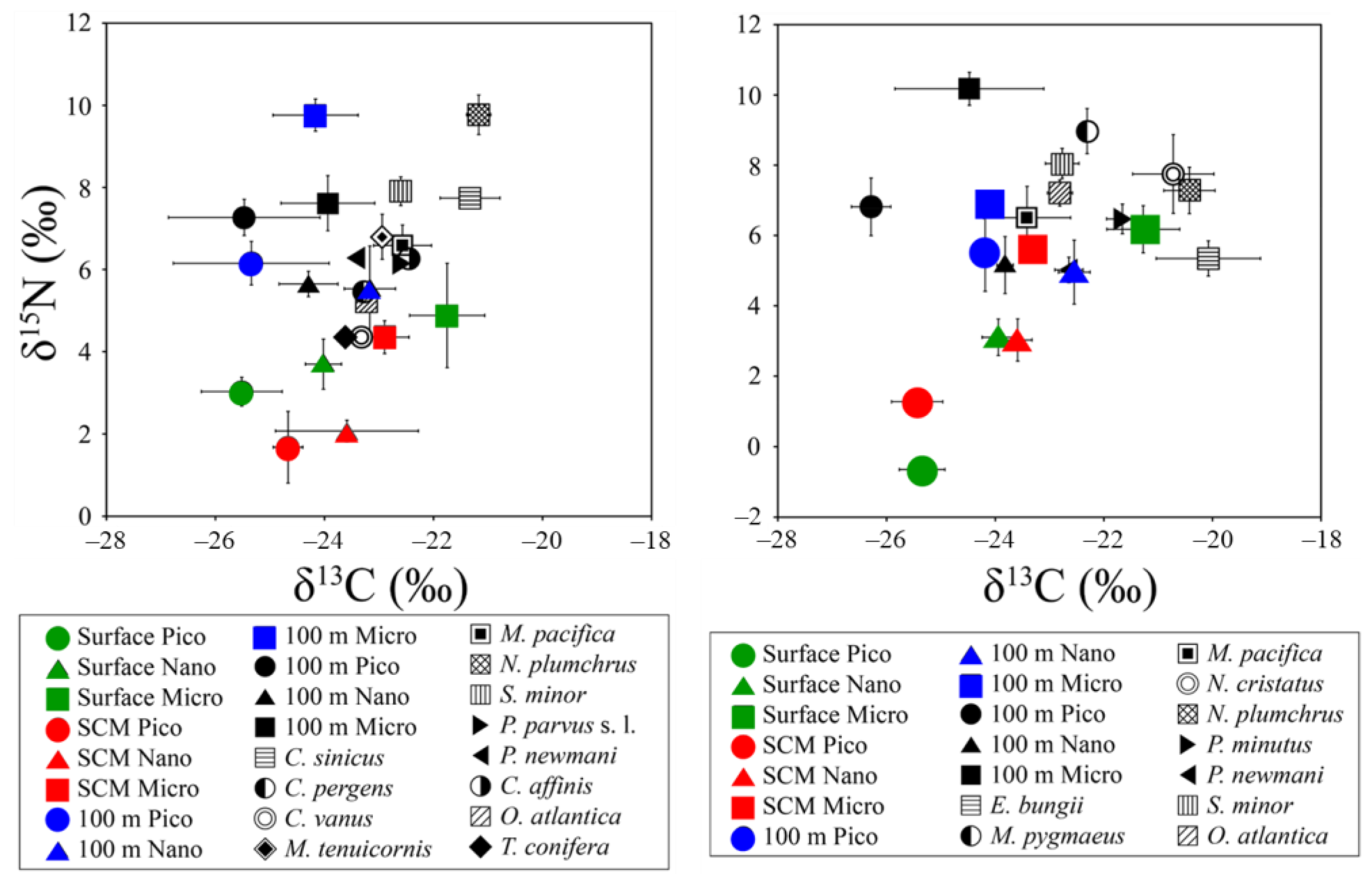
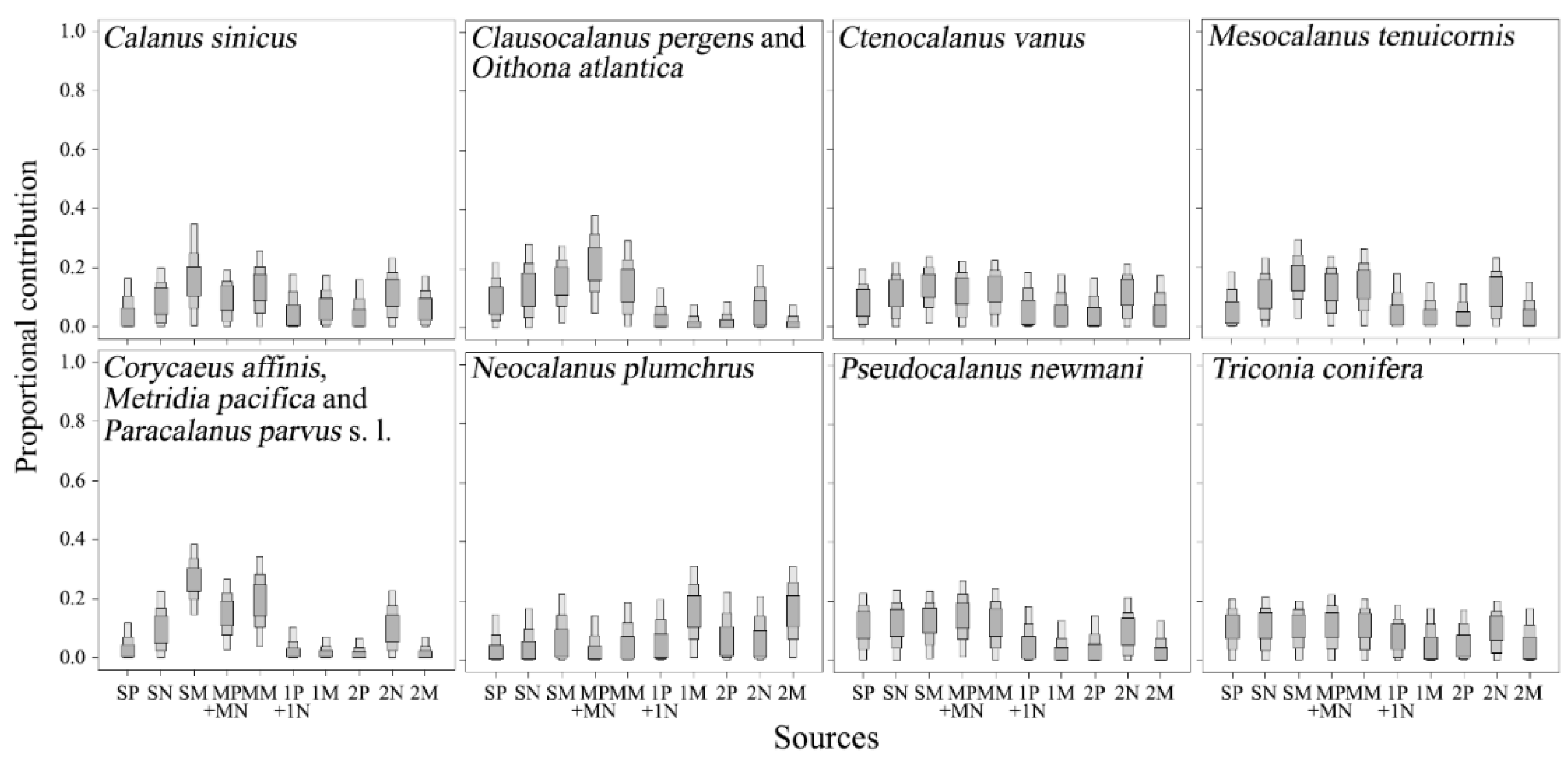
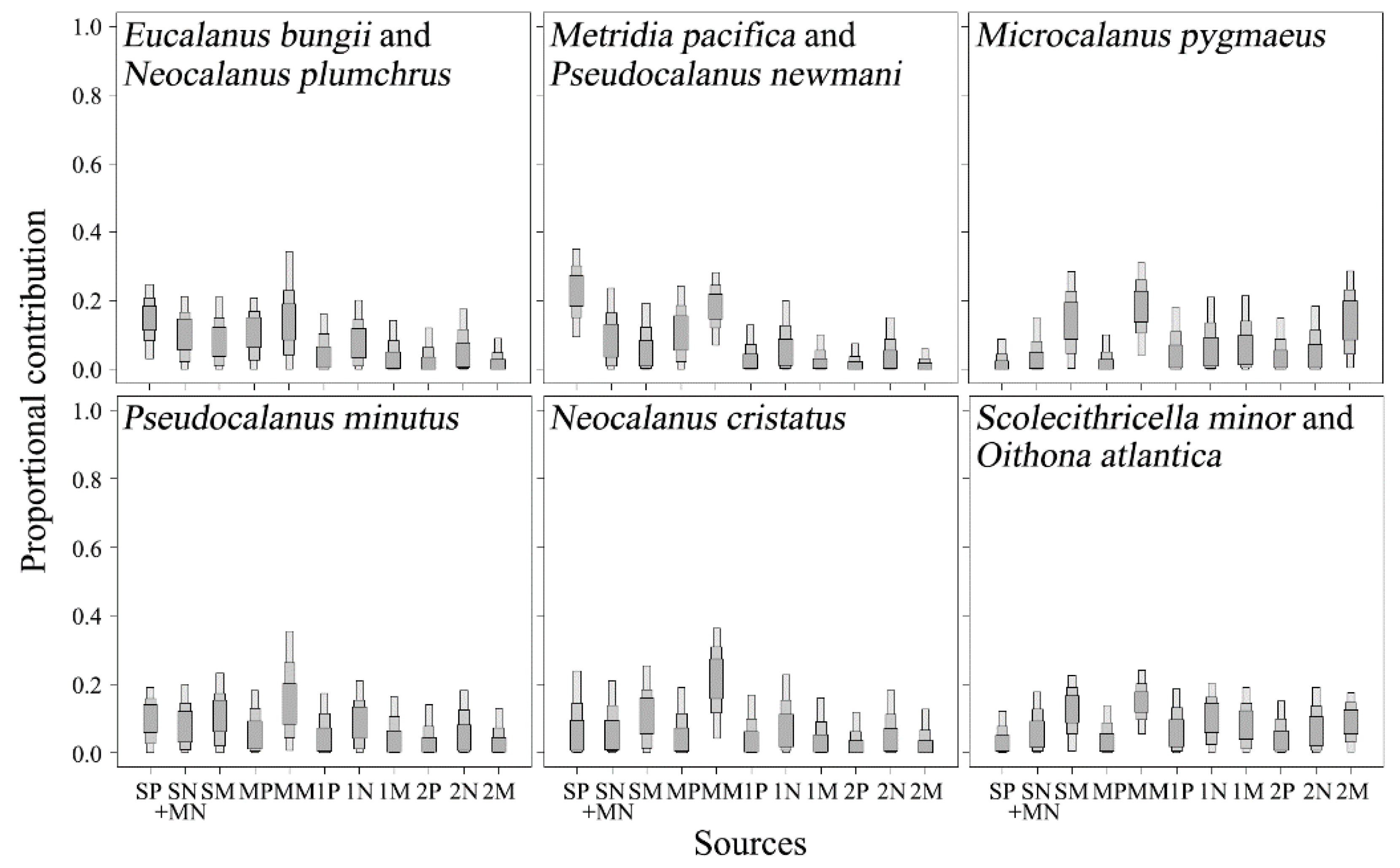
3.3. Vertical Profiles of δ13C and δ15N Values of Pico- to Micro-POMs
3.4. Carbon and Nitrogen Stable Isotope Ratios of Copepods
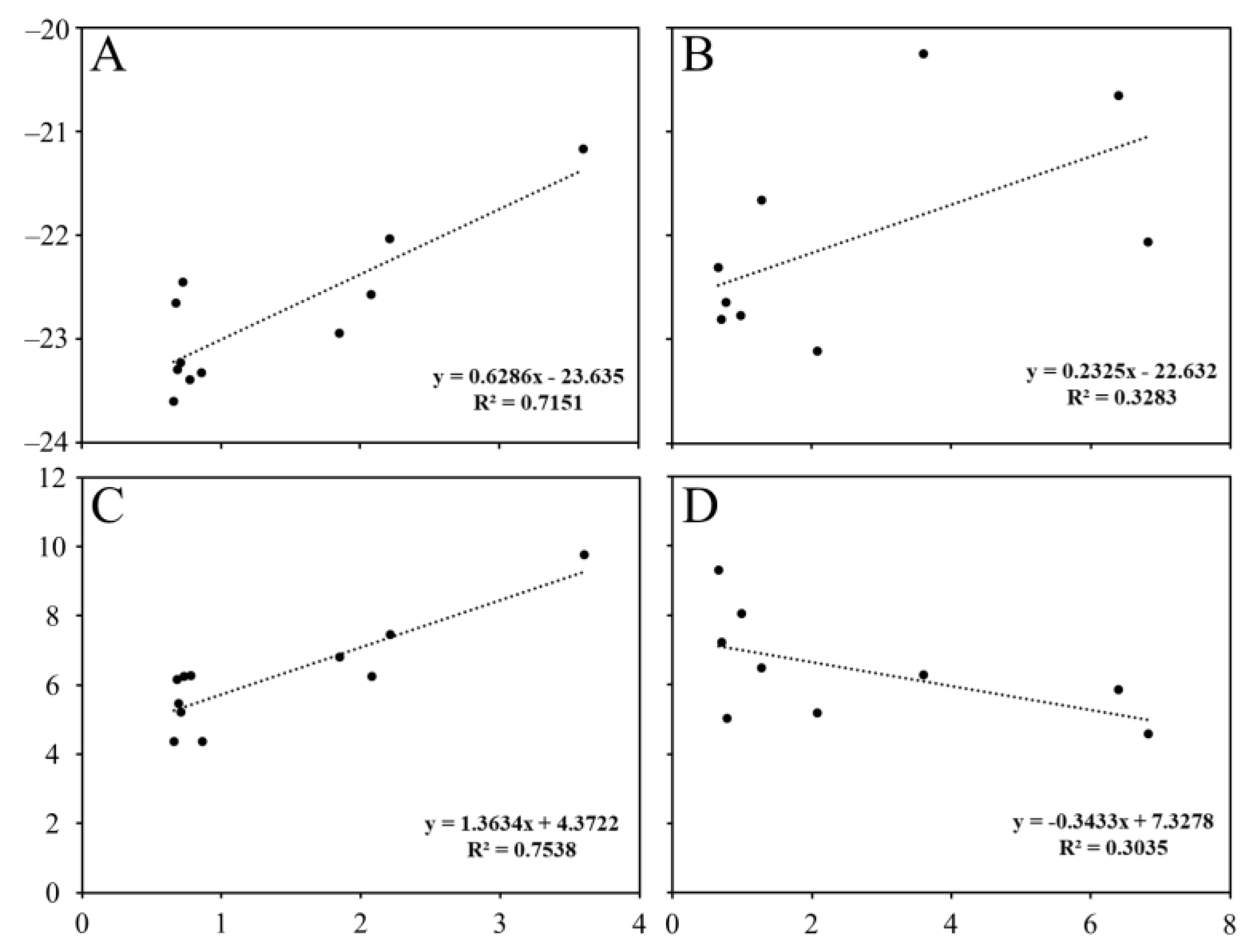
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, K.R.; Kim, K. What is happening in the East Sea (Japan Sea)? Recent chemical observations from CREAMS 93296. J. Korean Soc. Oceanogr. 1996, 31, 164–172. [Google Scholar]
- Talley, L.D.; Min, D.H.; Lobanov, V.B.; Luchin, V.A.; Ponomarev, V.I.; Salyuk, A.N.; Shcherbina, A.Y.; Tishchenko, P.Y.; Zhabin, I. Japan/East Sea water masses and their relation to the sea’s circulation. Oceanography 2006, 19, 32–49. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Kim, K.R.; Min, D.H.; Volkov, Y.; Yoon, J.H.; Takematsu, M. Warming and structure changes in the East (Japan) Sea: A clue to future changes in global oceans? Geophys. Res. Lett. 2001, 28, 3293–3296. [Google Scholar] [CrossRef]
- Kang, D.J.; Park, S.; Kim, Y.G.; Kim, K.; Kim, K.R. A moving-boundary box model (MBBM) for oceans in change: An application to the East/Japan Sea. Geophys. Res. Lett. 2003, 30, 1299. [Google Scholar] [CrossRef]
- Longhurst, A.R.; Harrison, W.G. The biological pump: Profiles of plankton production and consumption in the upper ocean. Prog. Oceanogr. 1989, 22, 47–123. [Google Scholar] [CrossRef]
- Fragoulis, C.; Christou, E.D.; Hecq, J.H. Comparison of marine copepod outfluxes: Nature, rate, fate and role in the carbon and nitrogen cycles. Adv. Mar. Biol. 2005, 47, 253–309. [Google Scholar]
- Kleppel, G.S. On the diets of calanoid copepods. Mar. Ecol. Prog. Ser. 1993, 99, 183–195. [Google Scholar] [CrossRef]
- Escribano, R.; Pérez, C.S. Variability in fatty acids of two marine copepods upon changing food supply in the coastal upwelling zone off Chile: Importance of the picoplankton and nanoplankton fractions. J. Mar. Biol. Assoc. UK 2010, 90, 301–313. [Google Scholar] [CrossRef]
- Ward, P.; Whitehouse, M.; Brandon, M.; Shreeve, R.; Woodd-Walker, R. Mesozooplankton community structure across the Antarctic Circumpolar Current to the north of South Georgia: Southern Ocean. Mar. Biol. 2003, 143, 121–130. [Google Scholar] [CrossRef]
- Elliott, D.T.; Pierson, J.J.; Roman, M.R. Relationship between environmental conditions and zooplankton community structure during summer hypoxia in the northern Gulf of Mexico. J. Plankton Res. 2012, 34, 602–613. [Google Scholar] [CrossRef] [Green Version]
- Beaugrand, G.; Ibanez, F.; Lindley, J.A. Diversity of calanoid copepods in the North Atlantic and adjacent seas: Species associations and biogeography. Mar. Ecol. Prog. Ser. 2002, 232, 179–195. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, G.E. The paradox of the plankton. Am. Nat. 1961, 92, 137–145. [Google Scholar] [CrossRef]
- Litchman, E.; Ohman, M.D.; Kiørboe, T. Trait-based approaches to zooplankton communities. J. Plankton Res. 2013, 35, 473–484. [Google Scholar] [CrossRef] [Green Version]
- Benedetti, F.; Gasparini, S.; Ayata, S.D. Identifying copepod functional groups from species functional traits. J. Plankton Res. 2016, 38, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Ashjian, C.J.; Davis, C.S.; Gallager, S.M.; Alatalo, P. Characterization of the zooplankton community, size composition, and distribution in relation to hydrography in the Japan/East Sea. Deep-Sea Res. II 2005, 52, 1363–1392. [Google Scholar] [CrossRef]
- Rho, T.K.; Kim, Y.B.; Park, J.I.; Lee, Y.W.; Im, D.H.; Kang, D.J.; Lee, T.S.; Yoon, S.T.; Kim, T.H.; Kwak, J.H.; et al. Plankton community response to physico-chemical forcing in the Ulleung Basin, East Sea during summer. Ocean Polar Res. 2010, 32, 269–289. [Google Scholar] [CrossRef] [Green Version]
- Doney, S.C.; Ruckelshaus, M.; Duffy, J.E.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N.; et al. Climate change impacts on marine ecosystems. Annu. Rev. Mar. Sci. 2012, 4, 11–37. [Google Scholar] [CrossRef] [Green Version]
- Choi, A.R.; Park, Y.G.; Choi, H.J. Changes in the Tsushima Warm Current and the impact under a global warming scenario in coupled climate models. Ocean Polar Res. 2013, 35, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Shokri, M.; Cozzoli, F.; Vignes, F.; Bertoli, M.; Pizzul, E.; Basset, A. Metabolic rate and climate change across latitudes: Evidence of mass-dependent responses in aquatic amphipods. J. Exp. Biol. 2022, 225, jeb244842. [Google Scholar] [CrossRef]
- Rubalcaba, J.G. Oceanic vertical migrators in a warming world. Nat. Clim. Chang. 2022, 12, 973–974. [Google Scholar] [CrossRef]
- Jung, S. Asynchronous responses of fish assemblages to climate-driven ocean regime shifts between the upper and deep layer in the Ulleung Basin of the East Sea from 1986 to 2010. Ocean Sci. J. 2014, 49, 1–10. [Google Scholar] [CrossRef]
- Joo, H.T.; Son, S.H.; Park, J.W.; Kang, J.J.; Jeong, J.Y.; Lee, C.I.; Kang, C.K.; Lee, S.H. Long-term pattern of primary productivity in the East/Japan Sea based on ocean color data derived from MODIS-aqua. Remote Sens. 2016, 8, 25. [Google Scholar] [CrossRef]
- Fry, B.; Sherr, E.B. δ13C measurements as indicators of carbon flow in marine and freshwater ecosystems. Contrib. Mar. Sci. 1984, 27, 13–47. [Google Scholar]
- Michener, R.H.; Schell, D.M. Stable isotope ratios as tracers in marine aquatic food webs. In Stable Isotopes in Ecology and Environmental Science; Lajtha, K., Michener, R.H., Eds.; Blackwell Scientific: Oxford, UK, 1994; pp. 138–158. [Google Scholar]
- Hobson, K.A.; Alisauskas, R.T.; Clark, R.G. Stable-nitrogen isotope enrichment in avian tissue due to fasting and nutritional stress: Implications for isotopic analysis of diet. Condor 1993, 95, 388–394. [Google Scholar] [CrossRef]
- Williams, R.L.; Wakeham, S.; McKinney, R.; Wishner, K.F. Trophic ecology and vertical patterns of carbon and nitrogen stable isotopes in zooplankton from oxygen minimum zone regions. Deep-Sea Res. I 2014, 90, 36–47. [Google Scholar] [CrossRef]
- Bode, M.; Hagen, W.; Schukat, A.; Teuber, L.; Fonseca-Batista, D.; Dehairs, F.; Auel, H. Feeding strategies of tropical and subtropical calanoid copepods throughout the eastern Atlantic Ocean—Latitudinal and bathymetric aspects. Prog. Oceanogr. 2015, 138, 268–282. [Google Scholar] [CrossRef]
- Im, D.H.; Suh, H.L. Ontogenetic feeding migration of the euphausiid Euphausia pacifica in the East Sea (Japan Sea) in autumn: A stable isotope approach. J. Plankton Res. 2016, 38, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Im, D.H.; Suh, H.L. Evidence for resource partitioning by ontogeny and species in calanoid copepods. Prog. Oceanogr. 2019, 176, 102111. [Google Scholar] [CrossRef]
- Uye, S. Length-weight relationships of important zooplankton from the Inland Sea of Japan. J. Oceanogr. Soc. Jpn. 1982, 38, 149–158. [Google Scholar] [CrossRef]
- Syväranta, J.; Vesala, S.; Rask, M.; Ruuhijärvi, J.; Jones, R.I. Evaluating the utility of stable isotope analyses of archived freshwater sample materials. Hydrobiologia 2008, 600, 121–130. [Google Scholar] [CrossRef]
- Bodin, N.; Le Loc, F.; Hily, C. Effect of lipid removal on carbon and nitrogen stable isotope ratios in crustacean tissues. J. Exp. Mar. Biol. Ecol. 2007, 341, 168–175. [Google Scholar] [CrossRef]
- Mintenbeck, K.; Brey, T.; Jacob, U.; Knust, R.; Struck, U. How to account for the lipid effect on carbon stable-isotope ratio (δ13C): Sample treatment effects and model bias. J. Fish Biol. 2008, 72, 815–830. [Google Scholar] [CrossRef] [Green Version]
- Smyntek, P.M.; Teece, M.A.; Schulz, K.L.; Thackeray, S.J. A standard protocol for stable isotope analysis of zooplankton in aquatic food web research using mass balance correction models. Limnol. Oceanogr. 2007, 52, 2135–2146. [Google Scholar] [CrossRef]
- Conover, W.J. Practical Nonparametric Statistics, 3rd ed.; John Wiley and Sons: New York, NY, USA, 1999. [Google Scholar]
- Stricker, D. BrightStat.com: Free statistics online. Comput. Methods Programs Biomed. 2008, 92, 135–143. [Google Scholar] [CrossRef]
- Parnell, A.C.; Inger, R.; Bearhop, S.; Jackson, A.L. Source partitioning using stable isotopes: Coping with too much variation. PLoS ONE 2010, 5, e9672. [Google Scholar] [CrossRef]
- McCutchan, J.H.; Lewis, W.M.; Kendall, C.; McGrath, C.C. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos 2003, 102, 378–390. [Google Scholar] [CrossRef]
- Im, D.H.; Wi, J.H.; Suh, H.L. Evidence for ontogenetic feeding strategies in four calanoid copepods in the East Sea (Japan Sea) in summer, revealed by stable isotope analysis. Ocean Sci. J. 2015, 50, 481–490. [Google Scholar] [CrossRef]
- Longhurst, A.R.; Sameoto, D.; Herman, A. Vertical distribution of Arctic zooplankton in summer: Eastern Canadian archipelago. J. Plankton Res. 1984, 6, 137–168. [Google Scholar] [CrossRef]
- Darnis, G.; Fortier, L. Temperature, food and the seasonal vertical migration of key arctic copepods in the thermally stratified Amundsen Gulf (Beaufort Sea, Arctic Ocean). J. Plankton Res. 2014, 36, 1092–1108. [Google Scholar] [CrossRef]
- Park, C.; Suh, H.L.; Kang, Y.S.; Ju, S.J.; Yang, E.J. Zooplankton. In Oceanography of the East Sea (Japan Sea); Chang, K.I., Zhang, C.I., Park, C., Kang, D.J., Ju, S.J., Lee, S.H., Wimbush, M., Eds.; Springer: Cham, Switzerland, 2016; pp. 297–326. [Google Scholar]
- Kwak, J.H.; Lee, S.H.; Park, H.J.; Choy, E.J.; Jeong, H.D.; Kim, K.R.; Kang, C.K. Monthly measured primary and new productivities in the Ulleung Basin as a biological “hot spot” in the East/Japan Sea. Biogeosciences 2013, 10, 4405–4417. [Google Scholar] [CrossRef] [Green Version]
- Volkman, J.K.; Tanoue, E. Chemical and biological studies of particulate organic matter in the ocean. J. Oceanogr. 2002, 58, 265–279. [Google Scholar] [CrossRef]
- Michels, J.; Schnack-Schiel, S.B. Feeding in dominant Antarctic copepods-does morphology of the mandibular gnathobases relate to diets? Mar. Biol. 2005, 146, 483–495. [Google Scholar] [CrossRef]
- Schnack-Schiel, S.B.; Mizdalski, E. Seasonal variations in distribution and population structure of Microcalanus pygmaeus and Ctenocalanus citer (Copepoda: Calanoida) in the eastern Weddell Sea, Antarctica. Mar. Biol. 1994, 119, 357–366. [Google Scholar] [CrossRef]
- Arashkevich, Y.G. The food and feeding of copepods in the north-western Pacific. Oceanology 1969, 9, 695–709. [Google Scholar]
- Nishida, S.; Ohtsuka, S. Ultrastructure of the mouthpart sensory setae in mesopelagic copepods of the family Scolecitrichidae. Plankton Biol. Ecol. 1997, 44, 81–90. [Google Scholar]
- Marlowe, C.J.; Miller, C.B. Patterns of vertical distribution and migration of zooplankton at Ocean Station “P”. Limnol. Oceanogr. 1975, 20, 824–844. [Google Scholar] [CrossRef]
- Yamaguchi, A.; Ikeda, T.; Hirakawa, K. Diel vertical migration, population structure and life cycle of the copepod Scolecithricella minor (Calanoida: Scolecitrichidae) in Toyama Bay, southern Japan Sea. Plankton Biol. Ecol. 1999, 46, 54–61. [Google Scholar]
- Turner, J.T. The importance of small planktonic copepods and their roles in pelagic marine food webs. Zool. Stud. 2004, 43, 255–266. [Google Scholar]
- Pasternak, A.F. Feeding of copepods of the genus Oncaea (Cyclopoida) in the southeastern Pacific Ocean. Oceanology 1984, 24, 609–612. [Google Scholar]
- Hopkins, T.L. Midwater food web in McMurdo Sound, Ross Sea. Mar. Biol. 1987, 96, 93–106. [Google Scholar] [CrossRef]
- González, H.E.; Smetacek, V. The possible role of the cyclopoid copepod Oithona in the retarding vertical flux of zooplankton fecal material. Mar. Ecol. Prog. Ser. 1994, 113, 233–246. [Google Scholar] [CrossRef]
- Alldredge, A.I. Abandoned larvacean houses: A unique food source in the pelagic environment. Science 1972, 177, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Metz, C. Feeding of Oncaea curvata (Poecilostomatoida, Copepoda). Mar. Ecol. Prog. Ser. 1998, 169, 229–235. [Google Scholar] [CrossRef]
- Wi, J.H.; Böttger-Schnack, R.; Soh, H.Y. Species of Triconia of the conifera-subgroup (Copepoda, Oncaeidae) from Korean waters, including a new species. J. Crustacean Biol. 2010, 30, 673–691. [Google Scholar] [CrossRef] [Green Version]
- Greene, C.H.; Landry, M.R. Carnivorous suspension feeding by the subarctic calanoid copepod Neocalanus cristatus. Can. J. Fish. Aquat. Sci. 1988, 45, 1069–1074. [Google Scholar] [CrossRef]
- Mackas, D.L.; Sefton, H.; Miller, C.B.; Raich, A. Vertical habitat partitioning by large calaniod copepods in the oceanic subarctic Pacific during spring. Prog. Oceanogr. 1993, 32, 259–294. [Google Scholar] [CrossRef]
- Falkenhaug, T.; Tande, K.S.; Semenova, T. Diel, seasonal and ontogenetic variations in the vertical distributions of four marine copepods. Mar. Ecol. Prog. Ser. 1997, 149, 105–119. [Google Scholar] [CrossRef] [Green Version]
- Hairston, N.G.; Hairston, N.G. Cause-effect relationships in energy flow, trophic structure and interspecific interactions. Am. Nat. 1993, 142, 379–411. [Google Scholar] [CrossRef]
- Cozzoli, F.; Shokri, M.; Boulamail, S.; Marrocco, V.; Vignes, F.; Basset, B. The size dependency of foraging behaviour: An empirical test performed on aquatic amphipods. Oecologia 2022, 199, 377–386. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Brose, U.; Jonsson, T.; Berlow, E.L.; Warren, P.; Banasek-Richter, C.; Bersier, L.F.; Blanchard, J.L.; Brey, T.; Capenter, S.R.; Blandenier, M.F.C.; et al. Consumer-resource body-size relationships in natural food webs. Ecology 2006, 87, 2411–2417. [Google Scholar] [CrossRef] [PubMed]
- Rolff, C. Seasonal variation in δ13C and δ15N of size-fractionated plankton at a coastal station in the northern Baltic proper. Mar. Ecol. Prog. Ser. 2000, 203, 47–65. [Google Scholar] [CrossRef] [Green Version]
- Basedow, S.L.; de Silva, N.A.L.; Bode, A.; van Beusekorn, J. Trophic positions of mesozooplankton across the North Atlantic: Estimates derived from biovolume spectrum theories and stable isotope analyses. J. Plankton Res. 2016, 38, 1364–1378. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Chang, C.Y.; García-Comas, C.; Gong, G.C.; Hsieh, C. Increasing zooplankton size diversity enhances the strength of top-down control on phytoplankton through diet niche partitioning. J. Anim. Ecol. 2013, 82, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- García-Comas, C.; Lee, Y.C.; Chang, C.Y.; Gong, G.C.; Hsieh, C. Comparison of copepod species-based and individual-size-based community structuring. J. Plankton Res. 2016, 38, 1006–1020. [Google Scholar] [CrossRef] [Green Version]
- Hérbert, M.P.; Beisner, B.E.; Maranger, R. Linking zooplankton communities to ecosystem functioning: Toward an effect trait framework. J. Plankton Res. 2017, 39, 3–12. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Im, D.-H.; Suh, H.-L. Trophic Positions of Sympatric Copepods across the Subpolar Front of the East Sea during Spring: A Stable Isotope Approach. Water 2023, 15, 416. https://doi.org/10.3390/w15030416
Im D-H, Suh H-L. Trophic Positions of Sympatric Copepods across the Subpolar Front of the East Sea during Spring: A Stable Isotope Approach. Water. 2023; 15(3):416. https://doi.org/10.3390/w15030416
Chicago/Turabian StyleIm, Dong-Hoon, and Hae-Lip Suh. 2023. "Trophic Positions of Sympatric Copepods across the Subpolar Front of the East Sea during Spring: A Stable Isotope Approach" Water 15, no. 3: 416. https://doi.org/10.3390/w15030416
APA StyleIm, D. -H., & Suh, H. -L. (2023). Trophic Positions of Sympatric Copepods across the Subpolar Front of the East Sea during Spring: A Stable Isotope Approach. Water, 15(3), 416. https://doi.org/10.3390/w15030416