
Trophic Assessment of an Artificial Kelp Eisenia bicyclis Bed Off the Eastern Coast of Korea Based on Stable Isotope Analyses

 ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Sites
2.2. Sample Collection and Treatment
2.3. Stable Isotope Analysis
2.4. Data Analyses
3. Results
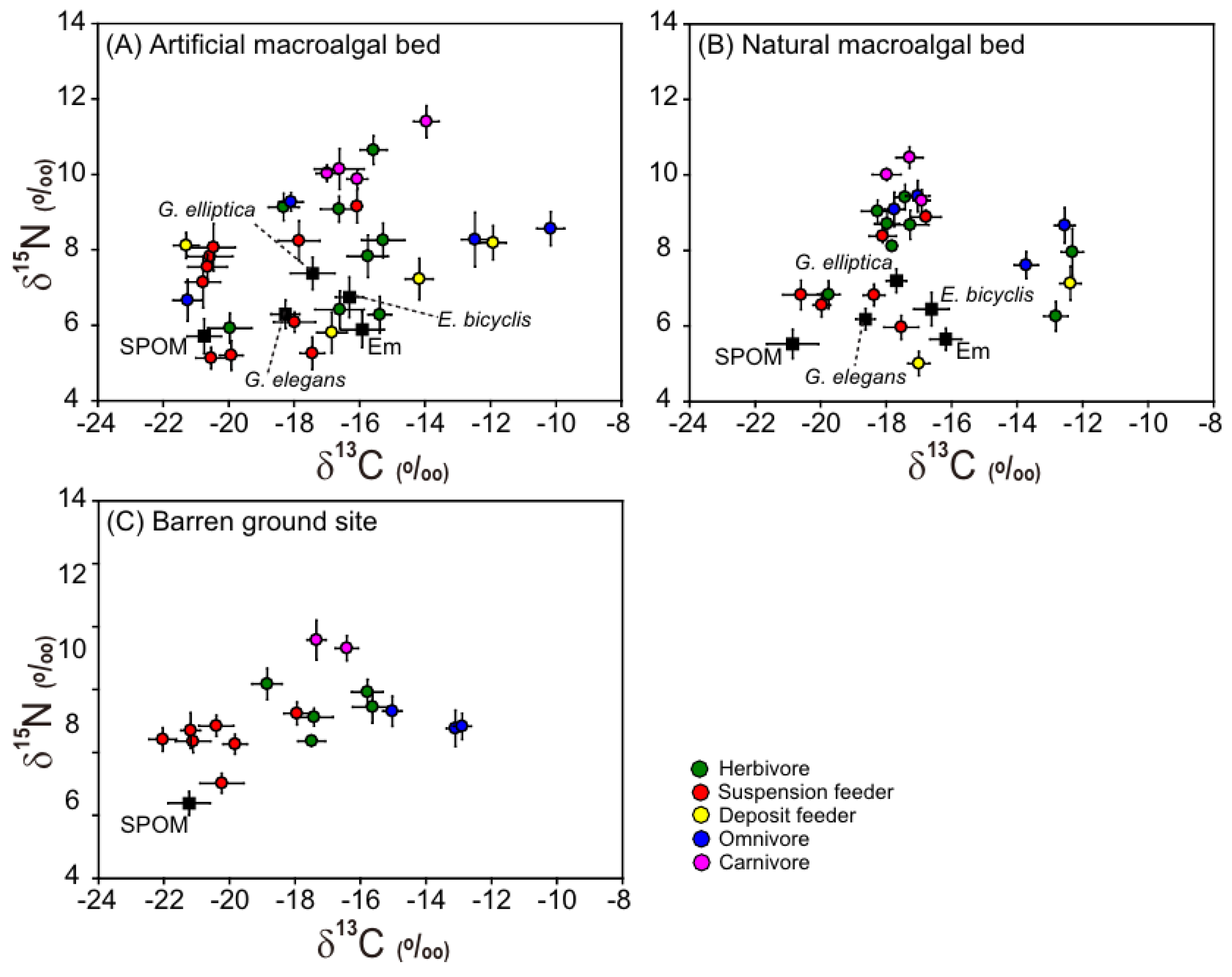
3.1. Carbon and Nitrogen Isotope Ratios of Organic Matter Sources
3.2. Carbon and Nitrogen Isotope Ratios of Macrobenthic Consumers
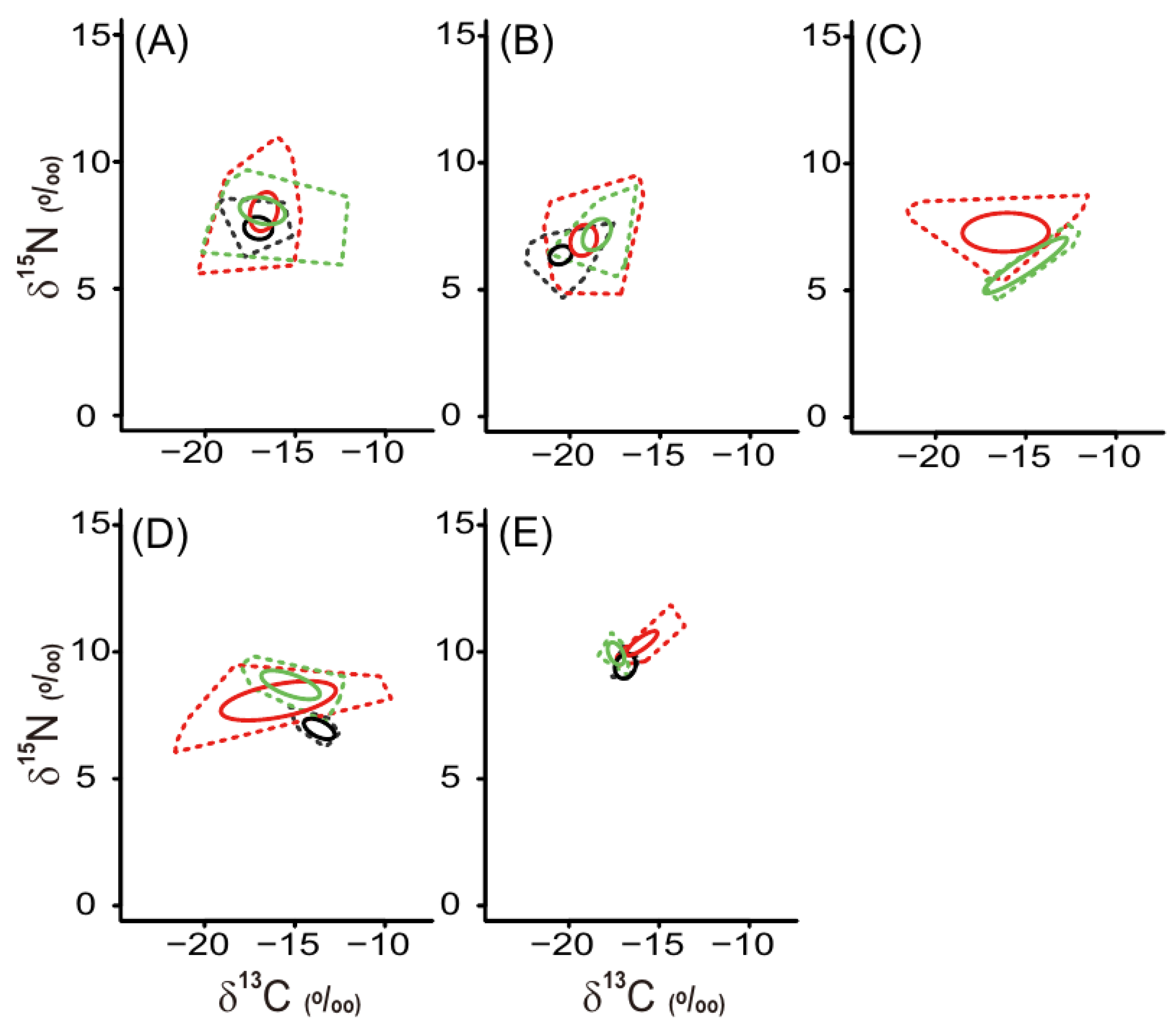
3.3. Isotopic Niche Indices of Macrobenthic Consumers
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bender, D.J.; Contreras, T.A.; Fahrig, L. Habitat loss and population decline: A meta-analysis of the patch size effect. Ecology 1998, 79, 517–533. [Google Scholar] [CrossRef]
- Airoldi, L.; Beck, W.M. Loss, status and trends for coastal marine habitats of Europe. Oceanogr. Mar. Biol. Annu. Rev. 2007, 45, 345–405. [Google Scholar]
- Gedan, K.B.; Silliman, B.R.; Bertness, M.D. Centuries of human-driven change in salt marsh ecosystems. Annu. Rev. Mar. Sci. 2009, 1, 117–141. [Google Scholar] [CrossRef] [Green Version]
- Steneck, R.S.; Graham, M.H.; Bourque, B.J.; Corbett, D.; Erlandson, J.M.; Estes, J.A.; Tegner, M.J. Kelp forest ecosystems: Biodiversity, stability, resilience and future. Environ. Conserv. 2002, 29, 436–459. [Google Scholar] [CrossRef] [Green Version]
- Bode, A.; Alvarez-Ossorio, M.T.; Varela, M. Phytoplankton and macrophyte contributions to littoral food webs in the Galician upwelling estimated from stable isotopes. Mar. Ecol. Prog. Ser. 2006, 318, 89–102. [Google Scholar] [CrossRef] [Green Version]
- Fulton, C.J.; Berkström, C.; Wilson, S.K.; Abesamis, R.A.; Bradley, M.; Åkerlund, C.; Barrett, L.T.; Bucol, A.A.; Chacin, D.H.; Chong-Seng, K.M. Macroalgal meadow habitats support fish and fisheries in diverse tropical seascapes. Fish Fish. 2020, 21, 700–717. [Google Scholar] [CrossRef]
- Seaman, W. Artificial habitats and the restoration of degraded marine ecosystems and fisheries. Hydrobiologia 2007, 580, 143–155. [Google Scholar] [CrossRef]
- Caselle, J.E.; Rassweiler, A.; Hamilton, S.L.; Warner, R.R. Recovery trajectories of kelp forest animals are rapid yet spatially variable across a network of temperate marine protected areas. Sci. Rep. 2015, 5, 14102. [Google Scholar] [CrossRef] [Green Version]
- Reed, D.C.; Schroeter, S.C.; Huang, D.; Anderson, T.W.; Ambrose, R.F. Quantitative assessment of different artificial reef designs in mitigating losses to kelp forest fishes. Bull. Mar. Sci. 2006, 78, 133–150. [Google Scholar]
- Campos, L.; Ortiz, M.; Rodríguez-Zaragoza, F.A.; Oses, R. Macrobenthic community establishment on artificial reefs with Macrocystis pyrifera over barren-ground and soft-bottom habitats. Glob. Ecol. Conserv. 2020, 23, e01184. [Google Scholar] [CrossRef]
- Tsiamis, K.; Salomidi, M.; Gerakaris, V.; Mogg, A.; Porter, E.; Sayer, M.; Küpper, F. Macroalgal vegetation on a north European artificial reef (Loch Linnhe, Scotland): Biodiversity, community types and role of abiotic factors. J. Appl. Phycol. 2020, 32, 1353–1363. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.; Chau, T.V.; Kim, M.; Na, W.-B. Artificial seaweed reefs that support the establishment of submerged aquatic vegetation beds and facilitate ocean macroalgal afforestation: A review. J. Mar. Sci. Eng. 2022, 10, 1184. [Google Scholar] [CrossRef]
- Choi, C.G.; Takeuchi, Y.; Terawaki, T.; Serisawa, Y.; Ohno, M.; Sohn, C.H. Ecology of seaweed beds on two types of artificial reef. J. Appl. Phycol. 2002, 14, 343–349. [Google Scholar] [CrossRef]
- Endo, H.; Nishigaki, T.; Yamamoto, K.; Takeno, K. Subtidal macroalgal succession and competition between the annual, Sargassum horneri, and the perennials, Sargassum patens and Sargassum piluliferum, on an artificial reef in Wakasa Bay, Japan. Fish. Sci. 2019, 85, 61–69. [Google Scholar] [CrossRef]
- Graham, M.H. Effects of local deforestation on the diversity and structure of southern California giant kelp forest food webs. Ecosystems 2004, 7, 341–357. [Google Scholar] [CrossRef]
- Boaventura, D.; Moura, A.; Leitao, F.; Carvalho, S.; Curdia, J.; Pereira, P.; Fonseca, L.C.D.; Santos, M.N.D.; Monteiro, C.C. Macrobenthic colonisation of artificial reefs on the southern coast of Portugal (Ancão, Algarve). Mar. Biodivers. 2006, 335–343. [Google Scholar]
- Santos, M.N.; Monteiro, C.C. A fourteen-year overview of the fish assemblages and yield of the two oldest Algarve artificial reefs (southern Portugal). Hydrobiologia 2007, 580, 225–231. [Google Scholar] [CrossRef]
- Kang, H.Y.; Lee, B.-G.; Park, H.J.; Yun, S.-G.; Kang, C.-K. Trophic structures of artificial reef communities off the southern coast of the Korean peninsula as determined using stable isotope analyses. Mar. Pollut. Bull. 2021, 169, 112474. [Google Scholar] [CrossRef]
- Park, H.J.; Park, T.H.; Kang, H.Y.; Lee, K.-S.; Kim, Y.K.; Kang, C.-K. Assessment of restoration success in a transplanted seagrass bed based on isotopic niche metrics. Ecol. Eng. 2021, 166, 106239. [Google Scholar] [CrossRef]
- Dunton, K.; Schell, D. Dependence of consumers on macroalgal (Laminaria solidungula) carbon in an arctic kelp community: δ13C evidence. Mar. Biol. 1987, 93, 615–625. [Google Scholar] [CrossRef]
- Jennings, S.; Reñones, O.; Morales-Nin, B.; Polunin, N.V.; Moranta, J.; Coll, J. Spatial variation in the 15N and 13C stable isotope composition of plants, invertebrates and fishes on Mediterranean reefs: Implications for the study of trophic pathways. Mar. Ecol. Prog. Ser. 1997, 146, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Layton, C.; Shelamoff, V.; Cameron, M.J.; Tatsumi, M.; Wright, J.T.; Johnson, C.R. Resilience and stability of kelp forests: The importance of patch dynamics and environment-engineer feedbacks. PLoS ONE 2019, 14, e0210220. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.K.; Choy, E.J.; Son, Y.; Lee, J.Y.; Kim, J.K.; Kim, Y.; Lee, K.S. Food web structure of a restored macroalgal bed in the eastern Korean peninsula determined by C and N stable isotope analyses. Mar. Biol. 2008, 153, 1181–1198. [Google Scholar] [CrossRef]
- Kim, M.J.; Yun, H.Y.; Shin, K.-H.; Kim, J.H. Evaluation of food web structure and complexity in the process of kelp bed recovery using stable isotope analysis. Front. Mar. Sci. 2022, 9, 885676. [Google Scholar] [CrossRef]
- Fry, B.; Sherr, E.B. δ13C measurements as indicators of carbon flow in marine and freshwater ecosystems. Contrib. Mar. Sci. 1984, 27, 13–47. [Google Scholar]
- Bearhop, S.; Adams, C.E.; Waldron, S.; Fuller, R.A.; Macleod, H. Determining trophic niche width: A novel approach using stable isotope analysis. J. Anim. Ecol. 2004, 73, 1007–1012. [Google Scholar] [CrossRef] [Green Version]
- Boecklen, W.J.; Yarnes, C.T.; Cook, B.A.; James, A.C. On the use of stable isotopes in trophic ecology. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 411–440. [Google Scholar] [CrossRef] [Green Version]
- Layman, C.A.; Araujo, M.S.; Boucek, R.; Hammerschlag-Peyer, C.M.; Harrison, E.; Jud, Z.R.; Post, D.M. Applying stable isotopes to examine food-web structure: An overview of analytical tools. Biol. Rev. 2012, 87, 545–562. [Google Scholar] [CrossRef]
- Peterson, B.J.; Fry, B. Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst. 1987, 18, 293–320. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Wozniak, A.S.; Roman, C.T.; Wainright, S.C.; McKinney, R.A.; James-Pirri, M.J. Monitoring food web changes in tide-restored salt marshes: A carbon stable isotope approach. Estuar. Coast. 2006, 29, 568–578. [Google Scholar] [CrossRef]
- Eger, A.M.; Marzinelli, E.M.; Christie, H.; Fagerli, C.W.; Fujita, D.; Gonzalez, A.P.; Hong, S.W.; Kim, J.H.; Lee, L.C.; McHugh, T.A. Global kelp forest restoration: Past lessons, present status, and future directions. Biol. Rev. 2022, 97, 1449–1475. [Google Scholar] [CrossRef]
- Anderson, M.J.; Clarke, K.R.; Gorley, R.N. PERMANOVA+ for Primer: Guide to Software and Statistical Methods; University of Auckland and PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER–Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Layman, C.A.; Arrington, D.A.; Montaña, C.G.; Post, D.M. Can stable isotope ratios provide for community-wide measures of trophic structure? Ecology 2007, 88, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Catry, T.; Lourenco, P.M.; Lopes, R.J.; Carneiro, C.; Alves, J.A.; Costa, J.; Rguibi-Idrissi, H.; Bearhop, S.; Piersma, T.; Granadeiro, J.P. Structure and functioning of intertidal food webs along an avian flyway: A comparative approach using stable isotopes. Funct. Ecol. 2016, 30, 468–478. [Google Scholar] [CrossRef]
- Rooney, N.; McCann, K.S.; Moore, J.C. A landscape theory for food web architecture. Ecol. Lett. 2008, 11, 867–881. [Google Scholar] [CrossRef]
- Taormina, B.; Claquin, P.; Vivier, B.; Navon, M.; Pezy, J.-P.; Raoux, A.; Dauvin, J.-C. A review of methods and indicators used to evaluate the ecological modifications generated by artificial structures on marine ecosystems. J. Environ. Manag. 2022, 310, 114646. [Google Scholar] [CrossRef] [PubMed]
- van Katwijk, M.M.; Thorhaug, A.; Marbà, N.; Orth, R.J.; Duarte, C.M.; Kendrick, G.A.; Althuizen, I.H.J.; Balestri, E.; Bernard, G.; Cambridge, M.L.; et al. Global analysis of seagrass restoration: The importance of large-scale planting. J. Appl. Ecol. 2016, 53, 567–578. [Google Scholar] [CrossRef] [Green Version]
- Wortley, L.; Hero, J.M.; Howes, M. Evaluating ecological restoration success: A review of the literature. Restor. Ecol. 2013, 21, 537–543. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Olden, J.D.; Gratton, C. Food-web approaches in restoration ecology. In Foundations of Restoration Ecology; Falk, D.A., Palmer, M., Zedler, J., Eds.; Island Press: Washington, DC, USA, 2006; pp. 165–189. [Google Scholar]
- Park, H.J.; Kwak, J.H.; Kang, H.Y.; Kwon, K.-Y.; Lim, W.; Kang, C.-K. Incorporation of Cochlodinium bloom-derived organic matter into a temperate subtidal macrobenthic food web as traced by stable isotopes. Mar. Pollut. Bull. 2020, 154, 111053. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.Y.; Lee, Y.-J.; Kim, C.; Kim, D.; Kim, D.-H.; Kim, J.-H.; Choi, D.-L.; Kang, C.-K. Food web trophic structure at marine ranch sites off the East coast of Korea. Front. Mar. Sci. 2021, 8, 653281. [Google Scholar] [CrossRef]
- Shin, D.; Park, T.H.; Lee, C.I.; Jeong, J.M.; Lee, S.-J.; Kang, S.; Park, H.J. Trophic ecology of largehead hairtail Trichiurus japonicus in the South Sea of Korea revealed by stable isotope and stomach content analyses. Front. Mar. Sci. 2022, 9, 910436. [Google Scholar] [CrossRef]
- James, W.R.; Lesser, J.S.; Litvin, S.Y.; Nelson, J.A. Assessment of food web recovery following restoration using resource niche metrics. Sci. Total Environ. 2020, 711, 134801. [Google Scholar] [CrossRef] [PubMed]
- Grall, J.; Le Loc’h, F.; Guyonnet, B.; Riera, P. Community structure and food web based on stable isotopes (δ15N and δ13C) analysis of a North Eastern Atlantic maerl bed. J. Exp. Mar. Biol. Ecol. 2006, 338, 1–15. [Google Scholar] [CrossRef]
- Park, H.J.; Han, E.; Lee, W.C.; Kwak, J.H.; Kim, H.C.; Park, M.S.; Kang, C.-K. Trophic structure in a pilot system for the integrated multi-trophic aquaculture off the east coast of Korean peninsula as determined by stable isotopes. Mar. Pollut. Bull. 2015, 95, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.K. Ascidian suspension feeding. J. Exp. Mar. Biol. Ecol. 2007, 342, 127–137. [Google Scholar] [CrossRef]
- Rezek, R.J.; Lebreton, B.; Roark, E.B.; Palmer, T.A.; Pollack, J.B. How does a restored oyster reef develop? An assessment based on stable isotopes and community metrics. Mar. Biol. 2017, 164, 54. [Google Scholar] [CrossRef]
- Ruppert, E.; Fox, R.; Barnes, R. Invertebrate Zoology, 7th ed.; Brooks/Cole Thompson Learning: Belmont, CA, USA, 2004. [Google Scholar]
- Kaehler, S.; Pakhomov, E.; McQuaid, C. Trophic structure of the marine food web at the Prince Edward Islands (Southern Ocean) determined by δ13C and δ15N analysis. Mar. Ecol. Prog. Ser. 2000, 208, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Fredriksen, S. Food web studies in a Norwegian kelp forest based on stable isotope (δ13C and δ15N) analysis. Mar. Ecol. Prog. Ser. 2003, 260, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Newsome, S.D.; Martinez del Rio, C.; Bearhop, S.; Phillips, D.L. A niche for isotopic ecology. Front. Ecol. Environ. 2007, 5, 429–436. [Google Scholar] [CrossRef]
- Rigolet, C.; Thiébaut, E.; Brind’Amour, A.; Dubois, S.F. Investigating isotopic functional indices to reveal changes in the structure and functioning of benthic communities. Funct. Ecol. 2015, 29, 1350–1360. [Google Scholar] [CrossRef] [Green Version]
- Quillien, N.; Nordström, M.C.; Schaal, G.; Bonsdorff, E.; Grall, J. Opportunistic basal resource simplifies food web structure and functioning of a highly dynamic marine environment. J. Exp. Mar. Biol. Ecol. 2016, 477, 92–102. [Google Scholar] [CrossRef]
- Guzzo, M.M.; Haffner, G.D.; Legler, N.D.; Rush, S.A.; Fisk, A.T. Fifty years later: Trophic ecology and niche overlap of a native and non-indigenous fish species in the western basin of Lake Erie. Biol. Invasions 2013, 15, 1695–1711. [Google Scholar] [CrossRef]
- Bustamante, R.H.; Branch, G.M. The dependence of intertidal consumers on kelp-derived organic matter on the west coast of South Africa. J. Exp. Mar. Biol. Ecol. 1996, 196, 1–28. [Google Scholar] [CrossRef]
- Moseman, S.M.; Levin, L.A.; Currin, C.; Forder, C. Colonization, succession, and nutrition of macrobenthic assemblages in a restored wetland at Tijuana Estuary, California. Estuar. Coast. Shelf Sci. 2004, 60, 755–770. [Google Scholar] [CrossRef]
- Nordström, M.C.; Demopoulos, A.W.J.; Whitcraft, C.R.; Rismondo, A.; McMillan, P.; Gonzalez, J.P.; Levin, L.A. Food web heterogeneity and succession in created saltmarshes. J. Appl. Ecol. 2015, 52, 1343–1354. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Organic Matter Source | Artificial Macroalgal Bed | Natural Macroalgal Bed | Barren Ground Site | PERMANOVA | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| δ13C | δ15N | δ13C | δ15N | δ13C | δ15N | ||||||||||||
| n | Mean | SD | Mean | SD | n | Mean | SD | Mean | SD | n | Mean | SD | Mean | SD | Pseudo-F | p-Value | |
| Eisenia bicyclis | 5 | −15.9 | 0.7 | 5.9 | 0.5 | 5 | −16.2 | 0.5 | 5.6 | 0.3 | 0.16 | 0.770 | |||||
| Gelidium elegans | 4 | −18.3 | 0.4 | 6.3 | 0.4 | 4 | −18.6 | 0.3 | 6.2 | 0.3 | 0.49 | 0.654 | |||||
| Grateloupia elliptica | 4 | −17.4 | 0.7 | 7.4 | 0.4 | 4 | −17.7 | 0.3 | 7.2 | 0.3 | 0.73 | 0.510 | |||||
| Epilithic microalgae | 4 | −16.3 | 0.4 | 6.7 | 0.5 | 4 | −16.6 | 0.5 | 6.4 | 0.4 | 0.47 | 0.642 | |||||
| SPOM | 5 | −20.7 | 0.5 | 5.7 | 0.5 | 5 | −20.9 | 0.8 | 5.5 | 0.4 | 5 | −21.2 | 0.6 | 4.4 | 0.4 | 7.83 | 0.003 |
| No. | Species Name | Taxon | Artificial Macroalgal Bed | Natural Macroalgal Bed | Barren Ground | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | δ13C | δ15N | n | δ13C | δ15N | n | δ13C | δ15N | |||||||||
| Herbivore | |||||||||||||||||
| 1 | Acmaea pallida | Gas | 4 | −16.6 | 0.6 | 9.1 | 0.3 | 3 | −17.4 | 0.3 | 9.4 | 0.3 | 3 | −15.6 | 0.6 | 7.4 | 0.5 |
| 2 | Aplysia kurodai | Gas | 3 | −15.4 | 0.3 | 6.3 | 0.5 | ||||||||||
| 3 | Cantharidus jessoensis | Gas | 3 | −15.7 | 0.6 | 7.8 | 0.6 | 3 | −17.8 | 0.1 | 8.1 | 0.1 | |||||
| 4 | Chlorostoma turbinata | Gas | 3 | −15.3 | 0.7 | 8.3 | 0.5 | 3 | −17.3 | 0.6 | 8.7 | 0.4 | 3 | −15.8 | 0.5 | 7.9 | 0.4 |
| 5 | Kelletia lischkei | Gas | 3 | −15.6 | 0.4 | 10.6 | 0.4 | ||||||||||
| 6 | Nipponacmea radula | Gas | −12.3 | 0.3 | 8.0 | 0.6 | |||||||||||
| 7 | Strongylocentrotus nudus | Ech | 3 | −16.6 | 0.7 | 6.4 | 0.5 | 3 | −12.8 | 0.4 | 6.3 | 0.4 | 3 | −17.5 | 0.4 | 6.4 | 0.1 |
| 8 | Tristichotrochus crossleyae | Gas | 3 | −18.0 | 0.4 | 8.7 | 0.3 | ||||||||||
| 9 | Tristichotrochus unicus | Gas | 4 | −18.3 | 0.5 | 9.1 | 0.4 | 3 | −18.3 | 0.5 | 9.0 | 0.3 | 3 | −18.8 | 0.5 | 8.2 | 0.5 |
| 10 | Turbo cornutus | Gas | 3 | −20.0 | 0.7 | 5.9 | 0.4 | 4 | −19.8 | 0.3 | 6.8 | 0.4 | 4 | −17.4 | 0.6 | 7.1 | 0.3 |
| Suspension feeder | |||||||||||||||||
| 11 | Anthopleura sp. | Cni | 3 | −17.8 | 0.6 | 8.2 | 0.5 | ||||||||||
| 12 | Boltenia echinata | Cho | 4 | −22.0 | 0.4 | 6.4 | 0.4 | ||||||||||
| 13 | Bostrycapulus gravispinosus | Gas | 3 | −17.4 | 0.4 | 5.3 | 0.4 | ||||||||||
| 14 | Cardita leana | Biv | −18.1 | 0.4 | 8.4 | 0.2 | |||||||||||
| 15 | Chelyosoma dofleini | Cho | 4 | −20.8 | 0.5 | 7.1 | 0.7 | ||||||||||
| 16 | Crassostrea nipponica | Biv | 3 | −19.8 | 0.4 | 6.3 | 0.3 | ||||||||||
| 17 | Crepidula onyx | Gas | 4 | −18.0 | 0.6 | 6.1 | 0.3 | 4 | −17.5 | 0.5 | 6.0 | 0.3 | 4 | −20.2 | 0.7 | 5.0 | 0.3 |
| 18 | Flustrellidra akkeshiensis | Bry | 3 | −20.5 | 0.5 | 5.1 | 0.3 | ||||||||||
| 19 | Halocynthia roretzi | Cho | 3 | −20.6 | 0.7 | 7.8 | 0.3 | 4 | −21.1 | 0.5 | 6.4 | 0.4 | |||||
| 20 | Hydroides ezoensis | Pol | 3 | −20.5 | 0.7 | 8.1 | 0.6 | 3 | −20.0 | 0.3 | 6.6 | 0.3 | 3 | −20.4 | 0.5 | 6.8 | 0.3 |
| 21 | Leiosolenus lischkei | Biv | −18.4 | 0.3 | 6.8 | 0.3 | |||||||||||
| 22 | Modiolus agripetus lredale | Biv | 3 | −19.9 | 0.4 | 5.2 | 0.4 | ||||||||||
| 23 | Phascolosoma scolops | Sip | 3 | −16.1 | 0.2 | 9.2 | 0.4 | 3 | −16.8 | 0.5 | 8.9 | 0.2 | 3 | −17.9 | 0.4 | 7.2 | 0.4 |
| 24 | Styela clava | Cho | 3 | −20.6 | 0.6 | 7.5 | 0.3 | 3 | −20.6 | 0.6 | 6.8 | 0.4 | 4 | −21.2 | 0.3 | 6.7 | 0.6 |
| Deposit feeder | |||||||||||||||||
| 25 | Ampithoe lacertosa | Cru | 4 | −16.8 | 0.5 | 5.8 | 0.5 | ||||||||||
| 26 | Holotelson tuberculatus | Cru | 3 | −21.3 | 0.4 | 8.1 | 0.3 | ||||||||||
| 27 | Pagurus proximus | Cru | 3 | −14.2 | 0.4 | 7.2 | 0.5 | ||||||||||
| 28 | Pagurus sp. | Cru | 3 | −12.4 | 0.3 | 7.1 | 0.4 | ||||||||||
| 29 | Perampithoe sp. | Cru | 4 | −17.0 | 0.3 | 5.0 | 0.3 | ||||||||||
| 30 | Pilumnus minutus | Cru | 3 | −11.9 | 0.4 | 8.2 | 0.5 | ||||||||||
| Omnivore | |||||||||||||||||
| 31 | Acanthochitona achates | Ppl | 4 | −21.3 | 0.4 | 6.7 | 0.5 | ||||||||||
| 32 | Asterias amurens | Ech | 3 | −13.7 | 0.4 | 7.6 | 0.4 | 3 | −15.0 | 0.3 | 7.3 | 0.5 | |||||
| 33 | Asterina pectinifera | Ech | 3 | −12.5 | 0.4 | 8.3 | 0.7 | 3 | −12.5 | 0.3 | 8.7 | 0.5 | 3 | −13.1 | 0.3 | 6.8 | 0.6 |
| 34 | Halosydna brevisetosa | Pol | 3 | −17.7 | 0.3 | 9.1 | 0.4 | ||||||||||
| 35 | Henricia leviuscula | Ech | 3 | −12.9 | 0.3 | 6.8 | 0.4 | ||||||||||
| 36 | Ophiarachnella gorgonia | Ech | 3 | −10.2 | 0.4 | 8.6 | 0.4 | ||||||||||
| 37 | Platynereis bicanaliculata | Pol | 3 | −18.1 | 0.4 | 9.3 | 0.2 | 3 | −17.0 | 0.4 | 9.4 | 0.4 | |||||
| Carnivore | |||||||||||||||||
| 38 | Leodice antennata | Pol | 3 | −17.0 | 0.3 | 10.0 | 0.2 | ||||||||||
| 39 | Lysidice collaris | Pol | 3 | −16.9 | 0.2 | 9.3 | 0.3 | ||||||||||
| 40 | Mitrella bicincta | Gas | 3 | −16.1 | 0.3 | 9.9 | 0.2 | 3 | −18.0 | 0.4 | 10.0 | 0.1 | 3 | −16.4 | 0.3 | 9.3 | 0.4 |
| 41 | Neptunea cumingii | Gas | 3 | −14.0 | 0.4 | 11.4 | 0.4 | ||||||||||
| 42 | Reishia bronni | Gas | 4 | −16.6 | 0.8 | 10.1 | 0.5 | 4 | −17.3 | 0.4 | 10.5 | 0.3 | 3 | −17.3 | 0.3 | 9.6 | 0.6 |
| Feeding Group | Artificial Macroalgal Bed | Natural Macroalgal Bed | Barren Ground Site | PERMANOVA | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| δ13C | δ15N | δ13C | δ15N | δ13C | δ15N | |||||||||||||
| n | Mean | SD | Mean | SD | n | Mean | SD | Mean | SD | n | Mean | SD | Mean | SD | Pseudo-F | p-Value | ||
| Herbivore | 26 | −16.7 | 1.6 | 8.0 | 1.6 | 25 | −16.8 | 2.6 | 8.1 | 1.1 | 16 | −17.1 | 1.3 | 7.4 | 0.7 | 1.12 | 0.376 | |
| Suspension feeder | 32 | −19.2 | 1.7 | 6.9 | 1.4 | 19 | −18.5 | 1.4 | 7.2 | 1.1 | 25 | −20.5 | 1.3 | 6.4 | 0.7 | 6.45 | 0.002 | |
| Deposit feeder | 13 | −16.1 | 3.5 | 7.2 | 1.1 | 7 | −15.0 | 2.5 | 5.9 | 1.2 | 1.94 | 0.153 | ||||||
| Omnivore | 13 | −15.9 | 4.7 | 8.1 | 1.1 | 12 | −15.3 | 2.3 | 8.7 | 0.8 | 9 | −13.7 | 1.0 | 7.0 | 0.5 | 2.84 | 0.046 | |
| Carnivore | 13 | −16.0 | 1.3 | 10.3 | 0.7 | 9 | −17.4 | 0.6 | 9.9 | 0.5 | 6 | −16.9 | 0.6 | 9.4 | 0.5 | 5.12 | 0.010 | |
| Isotopic niche areas | Artificial macroalgal bed | Natural macroalgal bed | Percentage overlap (%) | |||||||||||||||
| TA | SEAc | TA | SEAc | TA | SEAc | A/N | A/B | N/B | ||||||||||
| Herbivore | 20.69 | 8.27 | 22.24 | 8.97 | 6.14 | 3.09 | 70 | 54 | 39 | |||||||||
| Suspension feeder | 19.23 | 7.71 | 9.29 | 4.78 | 7.85 | 3.03 | 64 | 36 | 13 | |||||||||
| Deposit feeder | 19.11 | 13.60 | 4.82 | 4.04 | 14 | |||||||||||||
| Omnivore | 21.55 | 15.61 | 8.17 | 4.93 | 2.16 | 1.56 | 37 | 2 | 0 | |||||||||
| Carnivore | 3.67 | 1.72 | 1.50 | 1.00 | 1.13 | 1.10 | 13 | 18 | 30 | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, T.H.; Jang, J.-B.; Chun, C.-K.; Lee, Y.; Han, K.-S.; Yang, S.-J.; Park, H.J. Trophic Assessment of an Artificial Kelp Eisenia bicyclis Bed Off the Eastern Coast of Korea Based on Stable Isotope Analyses. Water 2023, 15, 1099. https://doi.org/10.3390/w15061099
Park TH, Jang J-B, Chun C-K, Lee Y, Han K-S, Yang S-J, Park HJ. Trophic Assessment of an Artificial Kelp Eisenia bicyclis Bed Off the Eastern Coast of Korea Based on Stable Isotope Analyses. Water. 2023; 15(6):1099. https://doi.org/10.3390/w15061099
Chicago/Turabian StylePark, Tae Hee, Jae-Bin Jang, Chan-Kil Chun, Youngkweon Lee, Kyu-Sam Han, Su-Jeong Yang, and Hyun Je Park. 2023. "Trophic Assessment of an Artificial Kelp Eisenia bicyclis Bed Off the Eastern Coast of Korea Based on Stable Isotope Analyses" Water 15, no. 6: 1099. https://doi.org/10.3390/w15061099
APA StylePark, T. H., Jang, J. -B., Chun, C. -K., Lee, Y., Han, K. -S., Yang, S. -J., & Park, H. J. (2023). Trophic Assessment of an Artificial Kelp Eisenia bicyclis Bed Off the Eastern Coast of Korea Based on Stable Isotope Analyses. Water, 15(6), 1099. https://doi.org/10.3390/w15061099