
Resource Use among Six Commercial Fish Species from the South-Eastern Gill Net Fisheries, Korea

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Stomach Content Analyses
2.3. Stable Isotope Analyses
2.4. Trophic Position
3. Results
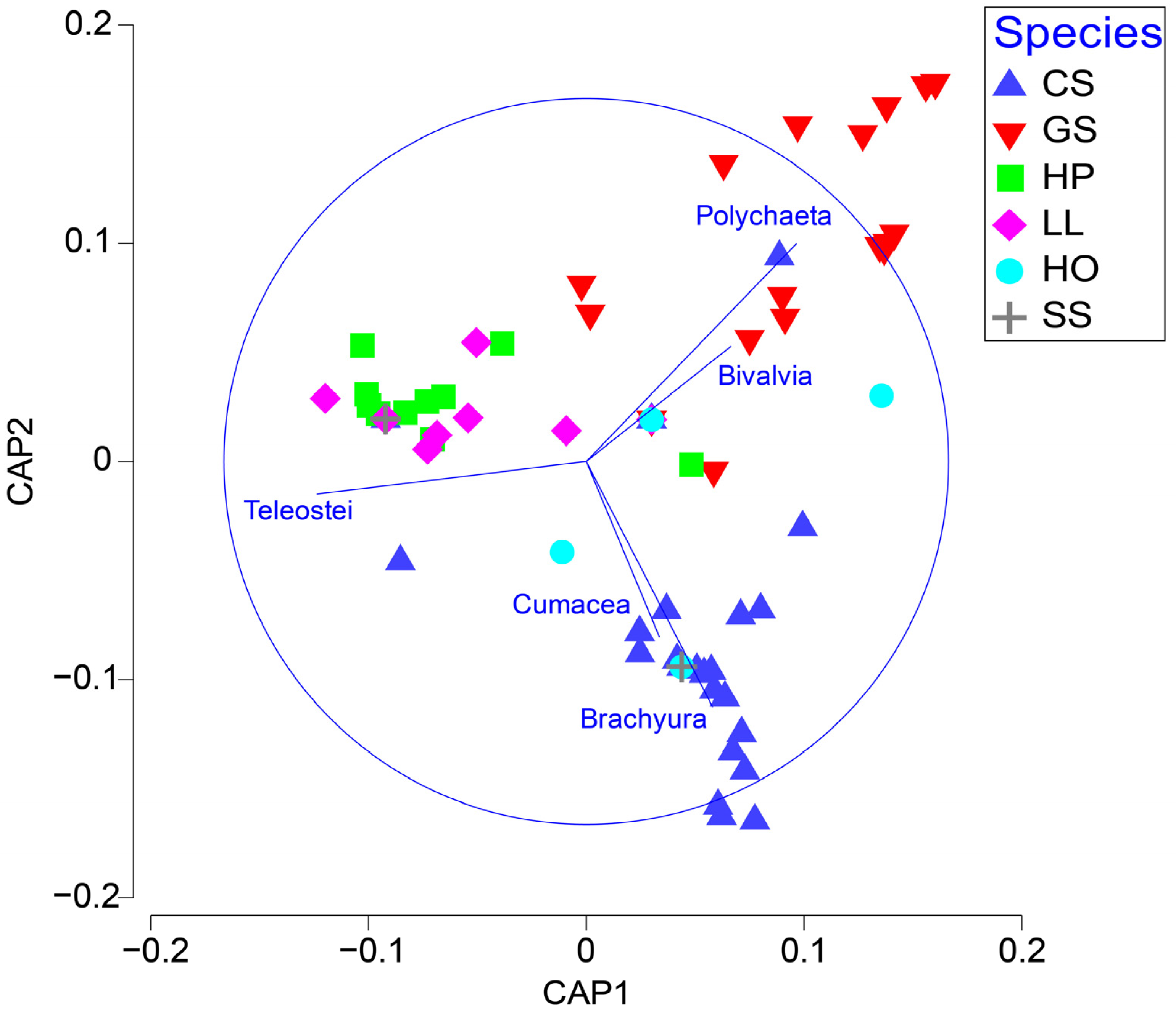
3.1. Trend in Dietary Composition
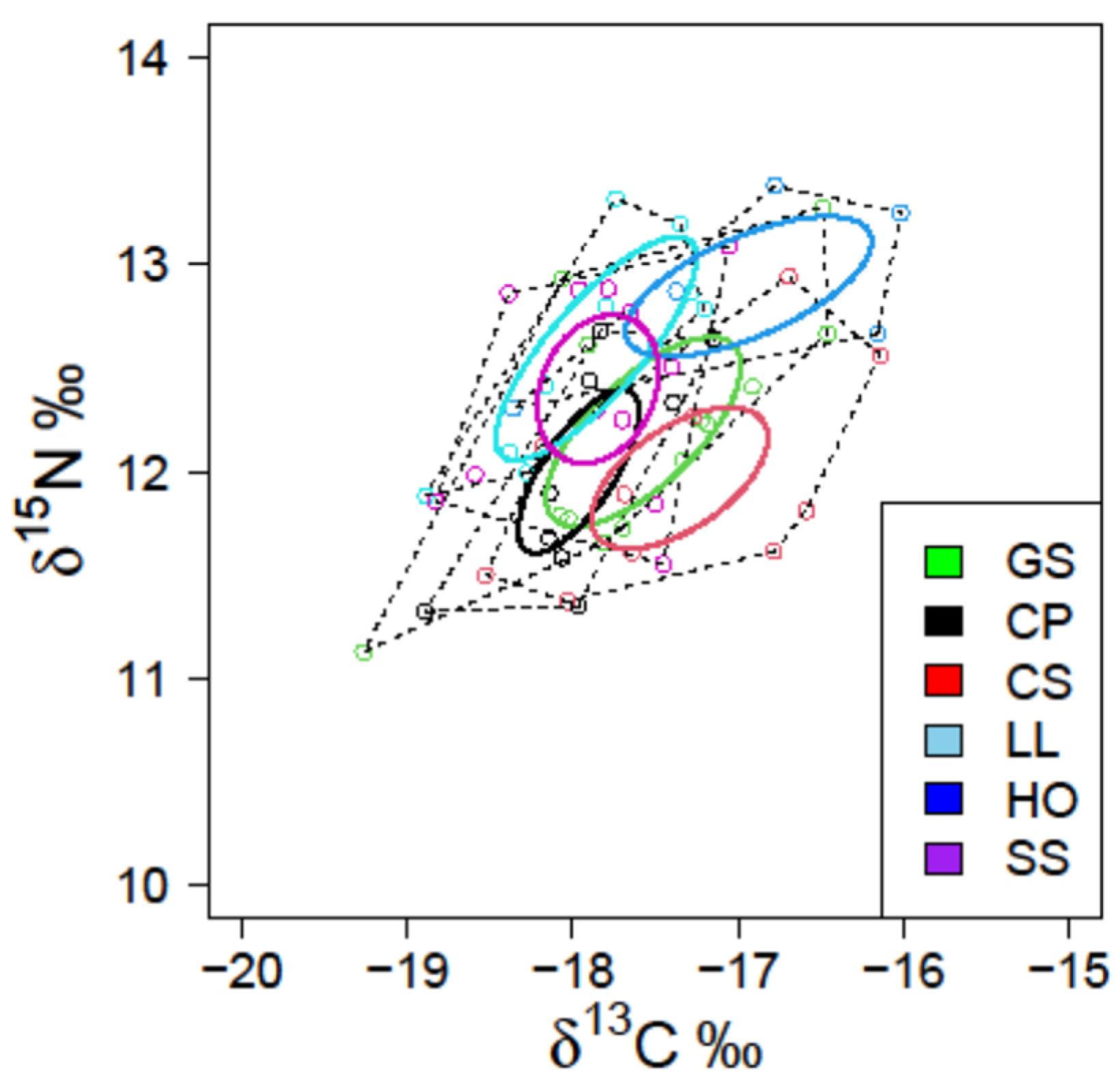
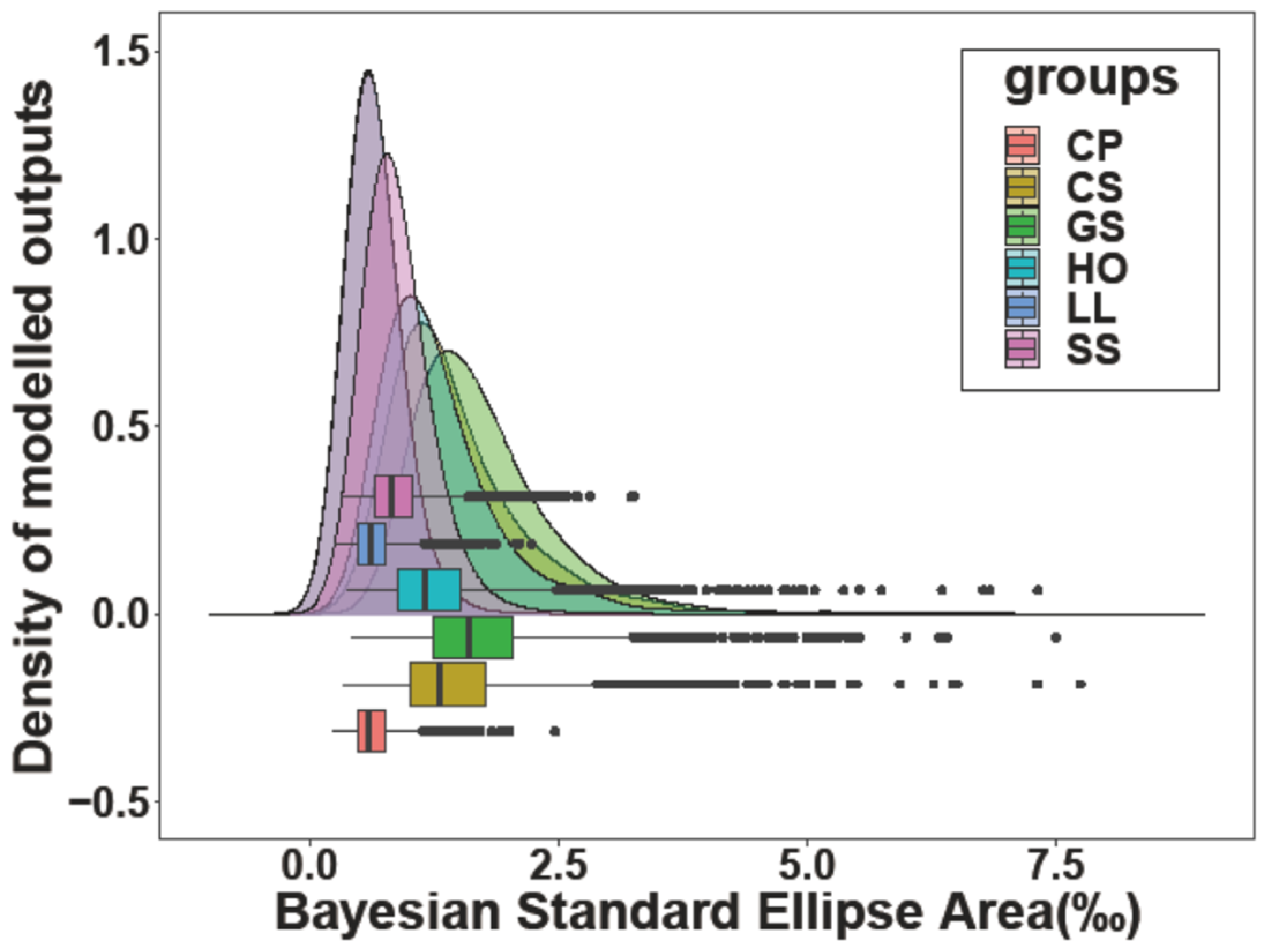
3.2. Stable Isotope Values
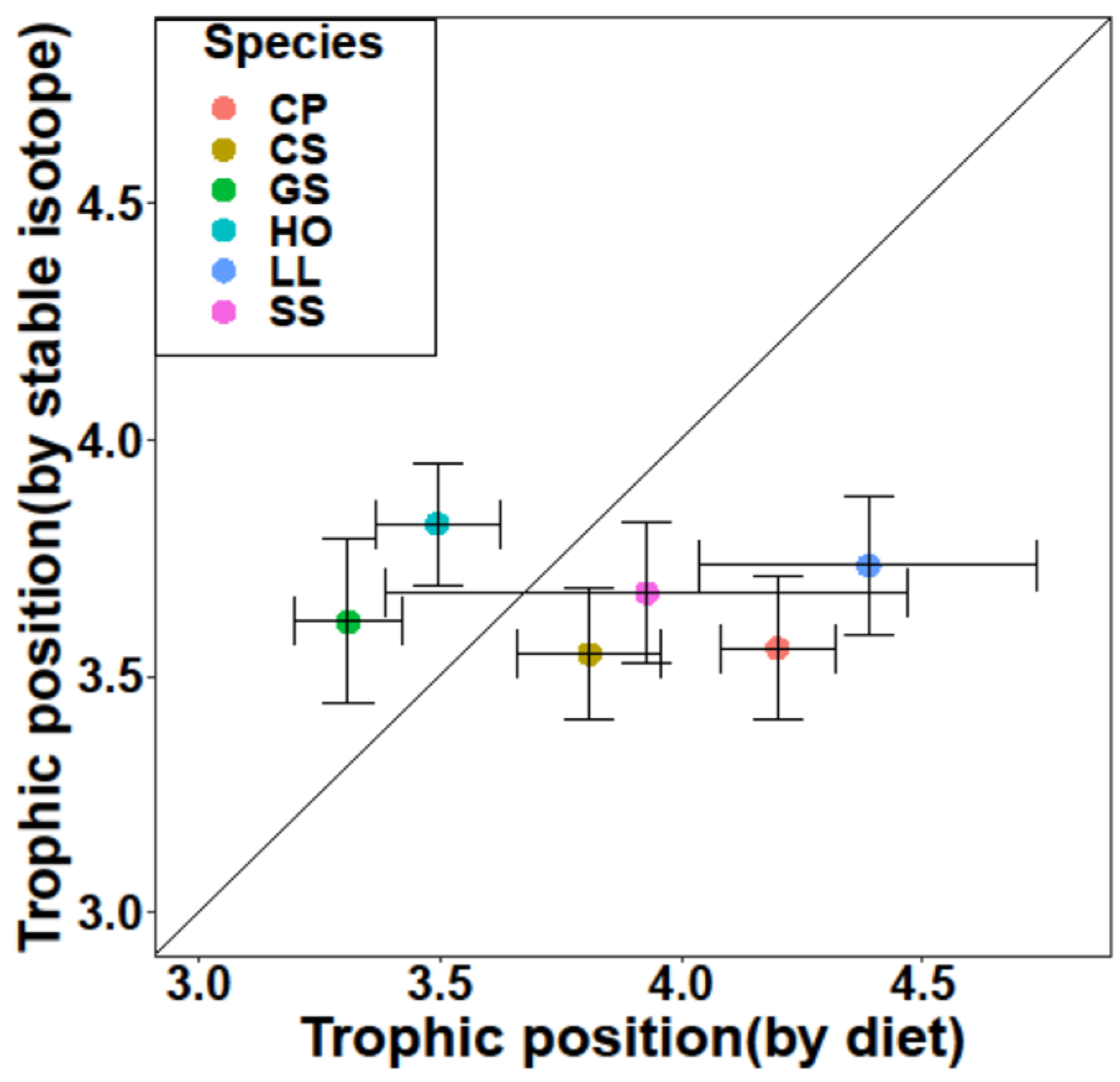
3.3. Trophic Position
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hussey, N.E.; MacNeil, M.A.; McMeans, B.C.; Olin, J.A.; Dudley, S.F.J.; Cliff, G.; Wintner, S.P.; Fennessy, S.T.; Fisk, A.T. Rescaling the trophic structure of marine food webs. Ecol. Lett. 2014, 17, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Froese, R.; Pauly., D. (Eds.) Fishbase; World Wide Web Electronic Publication. [Version 02/2022]; Available online: http://www.fishbase.org (accessed on 24 October 2022).
- Zanden, M.J.V.; Rasmussen, J.B. Variation in δ15N and δ13C trophic fractionation: Implications for aquatic food web studies. Limnol. Oceanogr. 2001, 46, 2061–2066. [Google Scholar] [CrossRef]
- Park, J.M.; Gaston, T.F.; Williamson, J.E. Resource partitioning in gurnard species using trophic analyses: The importance of temporal resolution. Fish. Res. 2017, 186, 301–310. [Google Scholar] [CrossRef]
- Amundsen, P.A.; Sánchez-Hernández, J. Feeding studies take guts–critical review and recommendations of methods for stomach contents analysis in fish. J. Fish Biol. 2019, 95, 1364–1373. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Tang, J.-P.; Su, L.-H.; Fan, J.-J.; Chang, H.-Y.; Wang, T.-T.; Wang, L.; Lin, H.-J.; Yang, Y. Fish feeding groups, food selectivity, and diet shifts associated with environmental factors and prey availability along a large subtropical river, China. Aquat. Sci. 2019, 81, 31. [Google Scholar] [CrossRef]
- Manko, P. Stomach Content Analysis in Freshwater Fish Feeding Ecology; University of Prešov; Prešov, Slovakia, 2016; Volume 116, p. 114. [Google Scholar]
- Park, J.M.; Kwak, S.N.; Huh, S.-H.; Han, I.-S. Diets and niche overlap among nine co-occurring demersal fishes in the southern continental shelf of East/Japan Sea, Korea. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 143, 100–109. [Google Scholar] [CrossRef]
- Park, J.M.; Huh, S.-H. Dietary habits and feeding strategy of the fivespot flounder, Pseudorhombus pentophthalmus in the southeastern coast of Korea. Ichthyol. Res. 2017, 64, 93–103. [Google Scholar] [CrossRef]
- Cortés, E. A critical review of methods of studying fish feeding based on analysis of stomach contents: Application to elasmobranch fishes. Can. J. Fish. Aquat. Sci. 1997, 54, 726–738. [Google Scholar] [CrossRef]
- Bearhop, S.; Adams, C.E.; Waldron, S.; Fuller, R.A.; MacLeod, H. Determining trophic niche width: A novel approach using stable isotope analysis. J. Anim. Ecol. 2004, 73, 1007–1012. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.-J.; Kao, W.-Y.; Wang, Y.-T. Analyses of stomach contents and stable isotopes reveal food sources of estuarine detritivorous fish in tropical/subtropical Taiwan. Estuar. Coast. Shelf Sci. 2007, 73, 527–537. [Google Scholar] [CrossRef]
- Cresson, P.; Ruitton, S.; Ourgaud, M.; Harmelin-Vivien, M. Contrasting perception of fish trophic level from stomach content and stable isotope analyses: A Mediterranean artificial reef experience. J. Exp. Mar. Biol. Ecol. 2014, 452, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Knickle, D.C.; Rose, G.A. Dietary niche partitioning in sympatric gadid species in coastal Newfoundland: Evidence from stomachs and CN isotopes. Environ. Biol. Fishes 2014, 97, 343–355. [Google Scholar] [CrossRef]
- Gibson, R.; Ezzi, I. Feeding relationships of a demersal fish assemblage on the west coast of Scotland. J. Fish Biol. 1987, 31, 55–69. [Google Scholar] [CrossRef]
- Fuita, T.; Kitagawa, D.; Okuyama, Y.; Ishito, Y.; Inada, T.; Jin, Y. Diets of the demersal fishes on the shelf off Iwate, northern Japan. Mar. Biol. 1995, 123, 219–233. [Google Scholar] [CrossRef]
- Labropoulou, M.; Papadopoulou-Smith, K.-N. Foraging behaviour patterns of four sympatric demersal fishes. Estuar. Coast. Shelf Sci. 1999, 49, 99–108. [Google Scholar] [CrossRef]
- Madurell, T.; Cartes, J.E. Trophic relationships and food consumption of slope dwelling macrourids from the bathyal Ionian Sea (eastern Mediterranean). Mar. Biol. 2006, 148, 1325–1338. [Google Scholar] [CrossRef]
- De la Morinière, E.C.; Pollux, B.; Nagelkerken, I.; Hemminga, M.; Huiskes, A.; van der Velde, G. Ontogenetic dietary changes of coral reef fishes in the mangrove-seagrass-reef continuum: Stable isotopes and gut-content analysis. Mar. Ecol. Prog. Ser. 2003, 246, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Dromard, C.R.; Bouchon-Navaro, Y.; Harmelin-Vivien, M.; Bouchon, C. Diversity of trophic niches among herbivorous fishes on a Caribbean reef (Guadeloupe, Lesser Antilles), evidenced by stable isotope and gut content analyses. J. Sea Res. 2015, 95, 124–131. [Google Scholar] [CrossRef]
- Yamada, U.; Shirai, S.; Irie, T. Names and Illustrations of Fishes from the East China Sea and the Yellow Sea: Japanese Chinese Korean; Overseas Fishery Cooperation Foundation: Tokyo, Japan, 1995; p. 288. [Google Scholar]
- Ryu, J.H.; Kim, P.-K.; Kim, J.K.; Kim, H.J. Seasonal variation of species composition of fishes collected by gill net and set net in the middle East Sea of Korea. Korean J. Ichthyol. 2005, 17, 279–286. [Google Scholar]
- Hong, B.-K.; Kim, J.-K.; Park, K.-D.; Jeon, K.-A.; Chun, Y.-Y.; Hwang, K.-e.; Kim, Y.-S.; Park, K.-Y. Species composition of fish collected in gill nets from Youngil Bay, East Sea of Korea. Korean J. Fish. Aquat. Sci. 2008, 41, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.H.; Kim, J.Y.; Kim, J.K.; Kim, J.B. Seasonal variation of species composition of fish in the coastal waters off Wolseong Nuclear Power Plant, East Sea of Korea by otter trawl survey. Korean J. Fish. Aquat. Sci. 2014, 47, 645–653. [Google Scholar] [CrossRef] [Green Version]
- Seong, G.C.; Ko, A.; Nam, K.M.; Jeong, J.M.; Kim, J.N.; Baeck, G.W. Diet of the Korean flounder Glyptocephalus stelleri in the coastal waters of the east sea of Korea. Korean J. Fish. Aquat. Sci. 2019, 52, 430–436. [Google Scholar] [CrossRef]
- Hyslop, E. Stomach contents analysis—A review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef] [Green Version]
- Da Silveira, E.L.; Semmar, N.; Cartes, J.E.; Tuset, V.M.; Lombarte, A.; Ballester, E.L.C.; Vaz-dos-Santos, A.M. Methods for Trophic Ecology Assessment in Fishes: A Critical Review of Stomach Analyses. Rev. Fish. Sci. Aquac. 2020, 28, 71–106. [Google Scholar] [CrossRef]
- Warwick, R.M. Environmental impact studies on marine communities: Pragmatical considerations. Aust. J. Ecol. 1993, 18, 63–80. [Google Scholar] [CrossRef]
- Clarke, K.; Gorley, R. Getting Started with PRIMER v7; PRIMER-E; Plymouth Marine Laboratory: Plymouth, UK, 2015; Volume 20, p. 18. [Google Scholar]
- Krebs, C.J. Ecological Methodology; Jarper & Row Publishers: New York, NY, USA, 1989. [Google Scholar]
- Peterson, B.J.; Fry, B. Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst. 1987, 18, 293–320. [Google Scholar] [CrossRef]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER—Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Syväranta, J.; Lensu, A.; Marjomäki, T.J.; Oksanen, S.; Jones, R.I. An empirical evaluation of the utility of convex hull and standard ellipse areas for assessing population niche widths from stable isotope data. PLoS ONE 2013, 8, e56094. [Google Scholar] [CrossRef] [Green Version]
- Pauly, D.; Froese, R.; Sa-a, P.; Palomares, M.; Christensen, V.; Rius, J. TrophLab Manual; Iclarm: Manila, Philippines, 2000. [Google Scholar]
- Pauly, D.; Froese, R.; Palomares, M.L. Fishing down aquatic food webs: Industrial fishing over the past half-century has noticeably depleted the topmost links in aquatic food chains. Am. Sci. 2000, 88, 46–51. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Kang, C.-K.; Choy, E.J.; Son, Y.; Lee, J.-Y.; Kim, J.K.; Kim, Y.; Lee, K.-S. Food web structure of a restored macroalgal bed in the eastern Korean peninsula determined by C and N stable isotope analyses. Mar. Biol. 2008, 153, 1181–1198. [Google Scholar] [CrossRef]
- Huh, S.-H.; Park, J.M.; Baeck, G.W. Feeding habits of bluefin searobin (Chelidonichthys spinosus) in the coastal waters off Busan. Korean J. Ichthyol. 2007, 19, 51–56. [Google Scholar]
- Kim, J.-B.; Kim, J.-Y.; Lee, D.-W.; Choi, J.-H. Feeding Habits of Bluefin Searobin Chelidonichthys spinosus around Jeju Island. Korean J. Fish. Aquat. Sci. 2011, 44, 378–382. [Google Scholar] [CrossRef]
- Park, J.; Huh, S.H.; Jeong, J.; Baeck, G. Diet composition and feeding strategy of yellow goosefish, Lophius litulon (Jordan, 1902), on the southeastern coast of Korea. J. Appl. Ichthyol. 2014, 30, 151–155. [Google Scholar] [CrossRef]
- Pushchina, O. Specific features of feeding of the Glyptocephalus stelleri and Acanthopsetta nadeshnyi in the northwestern Sea of Japan. J. Ichthyol. 2000, 40, 247–252. [Google Scholar]
- Tokranov, A. Specific features of distribution and some features of biology of Korean flounder Glyptocephalus stelleri (Pleuronectidae) in waters off Kamchatka in the Sea of Okhotsk. J. Ichthyol. 2008, 48, 759–769. [Google Scholar] [CrossRef]
- Park, K.-D.; Kang, Y.-J.; Huh, S.-H.; Kwak, S.-N.; Kim, H.-W.; Lee, H.-W. Feeding ecology of Sebastes schlegeli in the Tongyeong marine ranching area. Korean J. Fish. Aquat. Sci. 2007, 40, 308–314. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Li, Z.; Jin, X. Food composition and prey selectivity of Sebastes schlegeli. J. Fish. Sci. China 2014, 21, 134–141. [Google Scholar]
- Zhang, Y.; Xu, Q.; Xu, Q.; Alós, J.; Zhang, H.; Yang, H. Dietary Composition and Trophic Niche Partitioning of Spotty-bellied Greenlings Hexagrammos agrammus, Fat Greenlings H. otakii, Korean Rockfish Sebastes schlegelii, and Japanese Seaperch Lateolabrax japonicus in the Yellow Sea Revealed by Stomach Content Analysis and Stable Isotope Analysis. Mar. Coast. Fish. 2018, 10, 255–268. [Google Scholar]
- Zhang, R.; Liu, H.; Zhang, Q.; Zhang, H.; Zhao, J. Trophic interactions of reef-associated predatory fishes (Hexagrammos otakii and Sebastes schlegelii) in natural and artificial reefs along the coast of North Yellow Sea, China. Sci. Total Environ. 2021, 791, 148250. [Google Scholar] [CrossRef] [PubMed]
- Huh, S.; Baeck, G. Feeding habits of Hippoglossoides pinetorum collected in the coastal waters of Kori, Korea. Korean J. Ichthyol. 2003, 15, 157–161. [Google Scholar]
- Choi, H.C.; Huh, S.-H.; Park, J.M. Size-related and temporal dietary variations of Hexagrammos otakii in the mid-western coast of Korea. Korean J. Ichthyol. 2017, 29, 117–123. [Google Scholar]
- Gerking, S.D. Feeding Ecology of Fish; Academic Press: San Diego, CA, USA, 1994. [Google Scholar]
- Albouy, C.; Guilhaumon, F.; Villéger, S.; Mouchet, M.; Mercier, L.; Culioli, J.; Tomasini, J.; Mouillot, D. Predicting trophic guild and diet overlap from functional traits: Statistics, opportunities and limitations for marine ecology. Mar. Ecol. Prog. Ser. 2011, 436, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Norton, S.F. A functional approach to ecomorphological patterns of feeding in cottid fishes. In Ecomorphology of Fishes; Springer: Berlin/Heidelberg, Germany, 1995; pp. 61–78. [Google Scholar] [CrossRef]
- Fry, B. Stable Isotope Ecology; Springer: Berlin/Heidelberg, Germany, 2006; Volume 521. [Google Scholar]
- DeNiro, M.J.; Epstein, S. Influence of diet on the distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta 1978, 42, 495–506. [Google Scholar] [CrossRef]
- McConnaughey, T.; McRoy, C. Food-web structure and the fractionation of carbon isotopes in the Bering Sea. Mar. Biol. 1979, 53, 257–262. [Google Scholar] [CrossRef]
- Tieszen, L.L.; Boutton, T.W.; Tesdahl, K.G.; Slade, N.A. Fractionation and turnover of stable carbon isotopes in animal tissues: Implications for δ13C analysis of diet. Oecologia 1983, 57, 32–37. [Google Scholar] [CrossRef]
- Fry, B. Food web structure on Georges Bank from stable C, N, and S isotopic compositions. Limnol. Oceanogr. 1988, 33, 1182–1190. [Google Scholar] [CrossRef]
- Rossman, S.; Berens McCabe, E.; Barros, N.B.; Gandhi, H.; Ostrom, P.H.; Stricker, C.A.; Wells, R.S. Foraging habits in a generalist predator: Sex and age influence habitat selection and resource use among bottlenose dolphins (Tursiops truncatus). Mar. Mammal Sci. 2015, 31, 155–168. [Google Scholar] [CrossRef]
- Newsome, S.D.; Martinez del Rio, C.; Bearhop, S.; Phillips, D.L. A niche for isotopic ecology. Front. Ecol. Environ. 2007, 5, 429–436. [Google Scholar] [CrossRef]
- Gearing, J.N. The Study of Diet and Trophic Relationships through Natural Abundance 13C. In Carbon Isotope Techniques; Academic Press: San Diego, CA, USA, 1991; pp. 201–203. [Google Scholar]
- Macdonald, J.S.; Waiwood, K.G.; Green, R.H. Rates of digestion of different prey in Atlantic cod (Gadus morhua), ocean pout (Macrozoarces americanus), winter flounder (Pseudopleuronectes americanus), and American plaice (Hippoglossoides platessoides). Can. J. Fish. Aquat. Sci. 1982, 39, 651–659. [Google Scholar] [CrossRef]
- Polis, G.A.; Strong, D.R. Food web complexity and community dynamics. Am. Nat. 1996, 147, 813–846. [Google Scholar] [CrossRef]
- Azzurro, E.; Fanelli, E.; Mostarda, E.; Catra, M.; Andaloro, F. Resource partitioning among early colonizing Siganus luridus and native herbivorous fish in the Mediterranean: An integrated study based on gut-content analysis and stable isotope signatures. J. Mar. Biol. Assoc. UK 2007, 87, 991–998. [Google Scholar] [CrossRef] [Green Version]
- Sturbois, A.; Cozic, A.; Schaal, G.; Desroy, N.; Riera, P.; Le Pape, O.; Le Mao, P.; Ponsero, A.; Carpentier, A. Stomach content and stable isotope analyses provide complementary insights into the trophic ecology of coastal temperate bentho-demersal assemblages under environmental and anthropogenic pressures. Mar. Environ. Res. 2022, 182, 105770. [Google Scholar] [CrossRef] [PubMed]
- Trueman, C.N.; McGill, R.A.R.; Guyard, P.H. The effect of growth rate on tissue-diet isotopic spacing in rapidly growing animals. An experimental study with Atlantic salmon (Salmo salar). Rapid Commun. Mass Spectrom. 2005, 19, 3239–3247. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | CS | LL | GS | CP | HO | SS |
|---|---|---|---|---|---|---|
| Number of stomachs analyzed | 62 | 62 | 26 | 28 | 10 | 25 |
| Empty stomachs (%) | 33.3 | 41.9 | 19.2 | 50.0 | 50.0 | 68.0 |
| Diversity (H’) | 2.782 | 1.282 | 2.432 | 1.885 | 1.834 | 1.567 |
| MACRO ALGAE * | 0.10 | 0.36 | ||||
| POLYCHAETA | 0.34 | 37.31 | 13.08 | |||
| MOLLUSCA | ||||||
| Bivalvia | 1.03 | 11.28 | ||||
| Cephalopoda | 0.06 | 5.28 | 15.43 | 15.00 | ||
| CRUSTACEA | ||||||
| Malacostraca | ||||||
| Euphausiacea | 0.03 | |||||
| Stomatopoda | 0.05 | |||||
| Mysidacea | 0.03 | |||||
| Amphipoda | 0.13 | 3.79 | ||||
| Cumacea | 0.04 | |||||
| Decapoda unidentified | 0.77 | |||||
| Caridea | 21.56 | 9.19 | 30.80 | 20.22 | 20.00 | |
| Brachyura | 41.68 | 0.29 | 51.56 | 28.57 | ||
| Thoracica | 1.31 | |||||
| ECHINODERMATA | ||||||
| Crinoidea | ||||||
| CHORDATA | 28.57 | |||||
| Ascidiacea | ||||||
| Teleostei | 32.91 | 8.07 | ||||
| Unidentified material | 85.53 | 64.34 | 42.86 | |||
| Debris | 8.42 |
| CS | LL | GS | CP | HO | SS | |
| CS | - | 75.06 | 90.21 | 77.72 | 71.01 | 68.72 |
| LL | Teleostei Brachyura Caridea | - | 96.26 | 49.61 | 97.05 | 48.59 |
| GS | Brachyura Caridea Polychaeta | Teleostei Polychaeta Caridea | - | 90.09 | 86.83 | 99.66 |
| CP | Teleostei Brachyura Caridea | Teleostei Caridea Cephalopoda | Teleostei Caridea Polychaeta | - | 92.14 | 67.88 |
| HO | Brachyura Caridea Teleostei | Teleostei Brachyura Caridea | Brachyura Polychaeta Caridea | Teleostei Brachyura Caridea | - | 76.97 |
| SS | Teleostei Brachyura Caridea | Teleostei Brachyura Caridea | Teleostei Brachyura Polychaeta | Teleostei Brachyura Caridea | Teleostei Brachyura Caridea | - |
| CS | LL | GS | CP | HO | SS | |
|---|---|---|---|---|---|---|
| SEA | 1.016 | 0.524 | 1.019 | 0.507 | 1.015 | 0.807 |
| SEAC | 1.109 | 0.589 | 1.147 | 0.571 | 1.353 | 0.887 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, M.-G.; Lee, S.-H.; Kim, M.-J.; Kwak, S.-N.; Han, I.-S.; Park, J.-M. Resource Use among Six Commercial Fish Species from the South-Eastern Gill Net Fisheries, Korea. Water 2023, 15, 1146. https://doi.org/10.3390/w15061146
Kang M-G, Lee S-H, Kim M-J, Kwak S-N, Han I-S, Park J-M. Resource Use among Six Commercial Fish Species from the South-Eastern Gill Net Fisheries, Korea. Water. 2023; 15(6):1146. https://doi.org/10.3390/w15061146
Chicago/Turabian StyleKang, Min-Gu, Sang-Heon Lee, Myung-Joon Kim, Seok-Nam Kwak, In-Seong Han, and Joo-Myun Park. 2023. "Resource Use among Six Commercial Fish Species from the South-Eastern Gill Net Fisheries, Korea" Water 15, no. 6: 1146. https://doi.org/10.3390/w15061146
APA StyleKang, M. -G., Lee, S. -H., Kim, M. -J., Kwak, S. -N., Han, I. -S., & Park, J. -M. (2023). Resource Use among Six Commercial Fish Species from the South-Eastern Gill Net Fisheries, Korea. Water, 15(6), 1146. https://doi.org/10.3390/w15061146