

Diversity and Community Composition of Labyrinthulomycetes Protists in the Coastal Zone of Hainan Island, South China Sea



 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Water Sampling and Environmental Data
2.2. Quantitative PCR Analysis
2.3. High-Throughput Sequencing and Bioinformatics Analysis
2.4. Statistical Analyses
3. Results
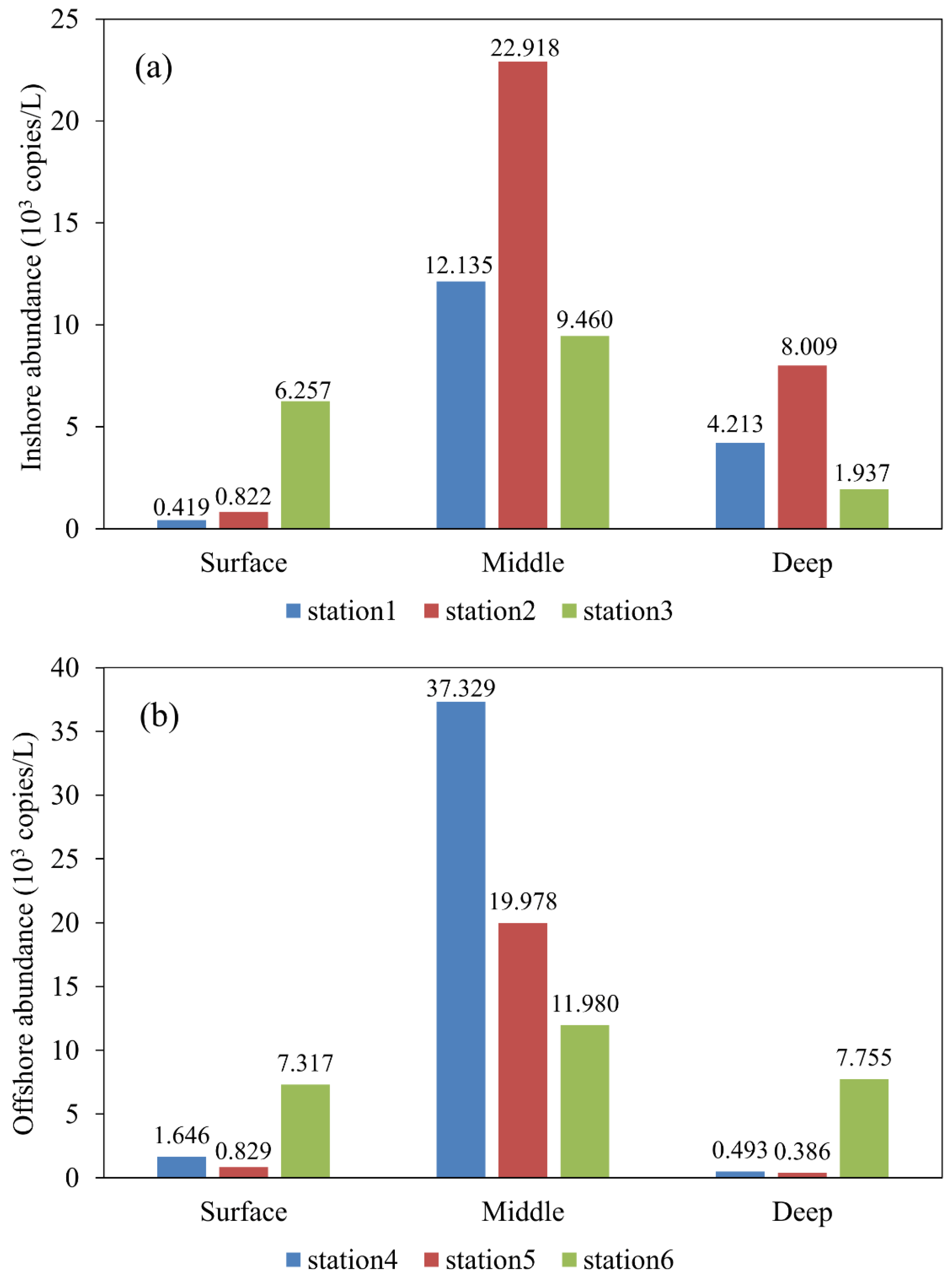
3.1. Abundance Pattern of Labyrinthulomycetes Protists
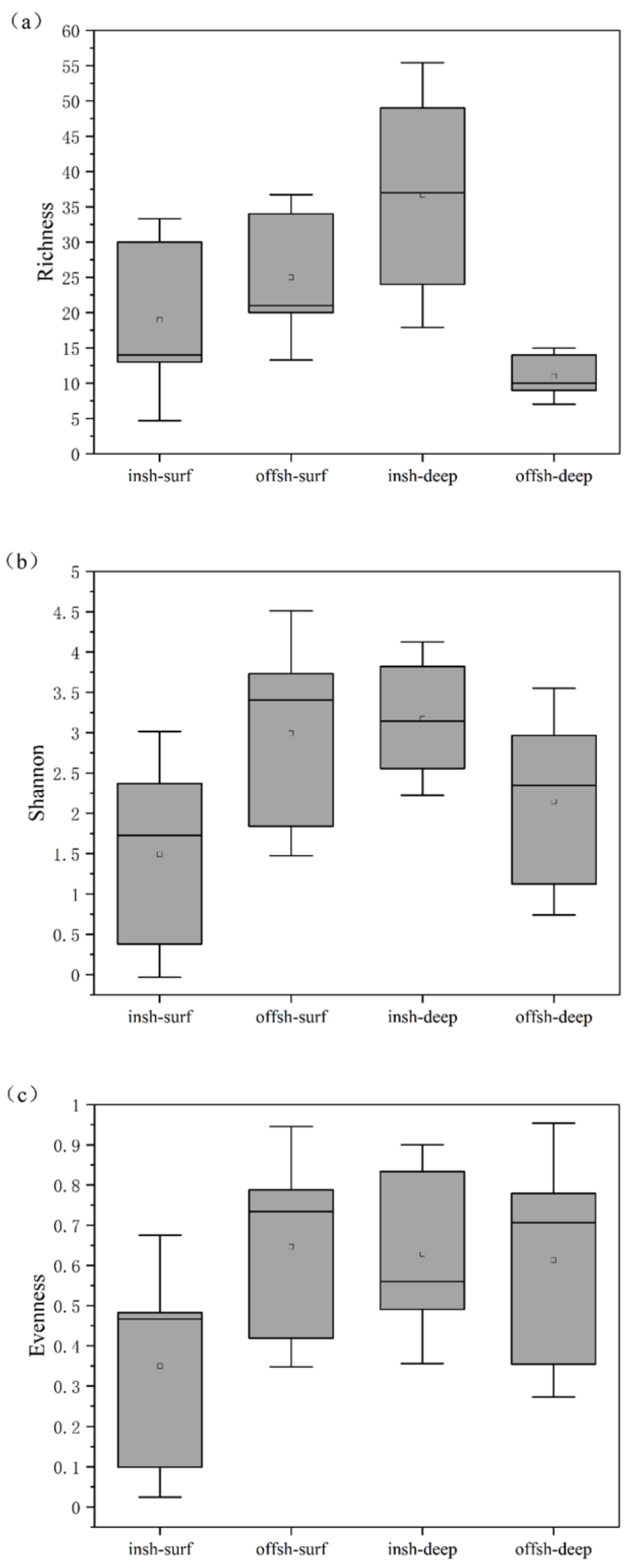
3.2. Diversity Patterns of Labyrinthulomycetes Protists
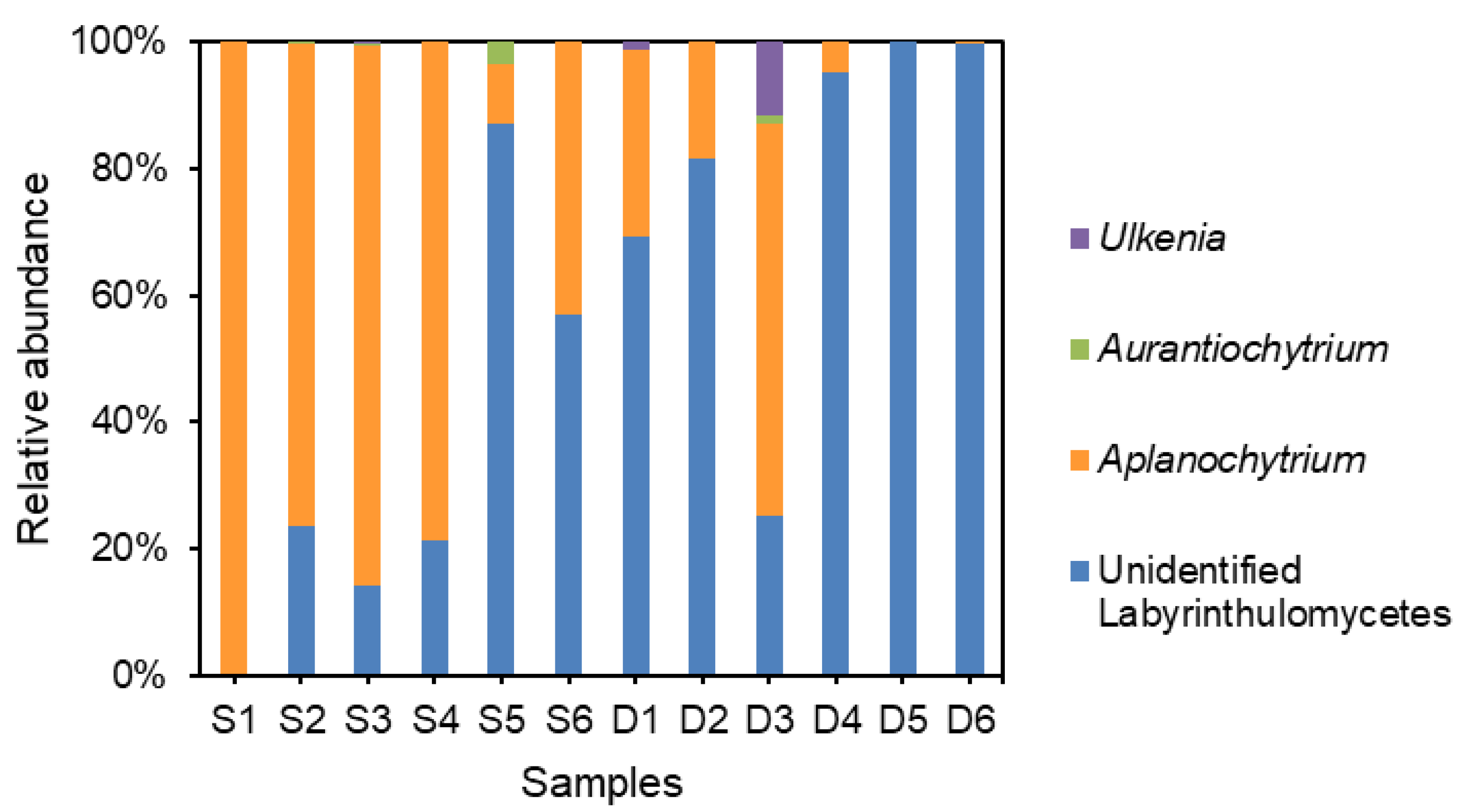
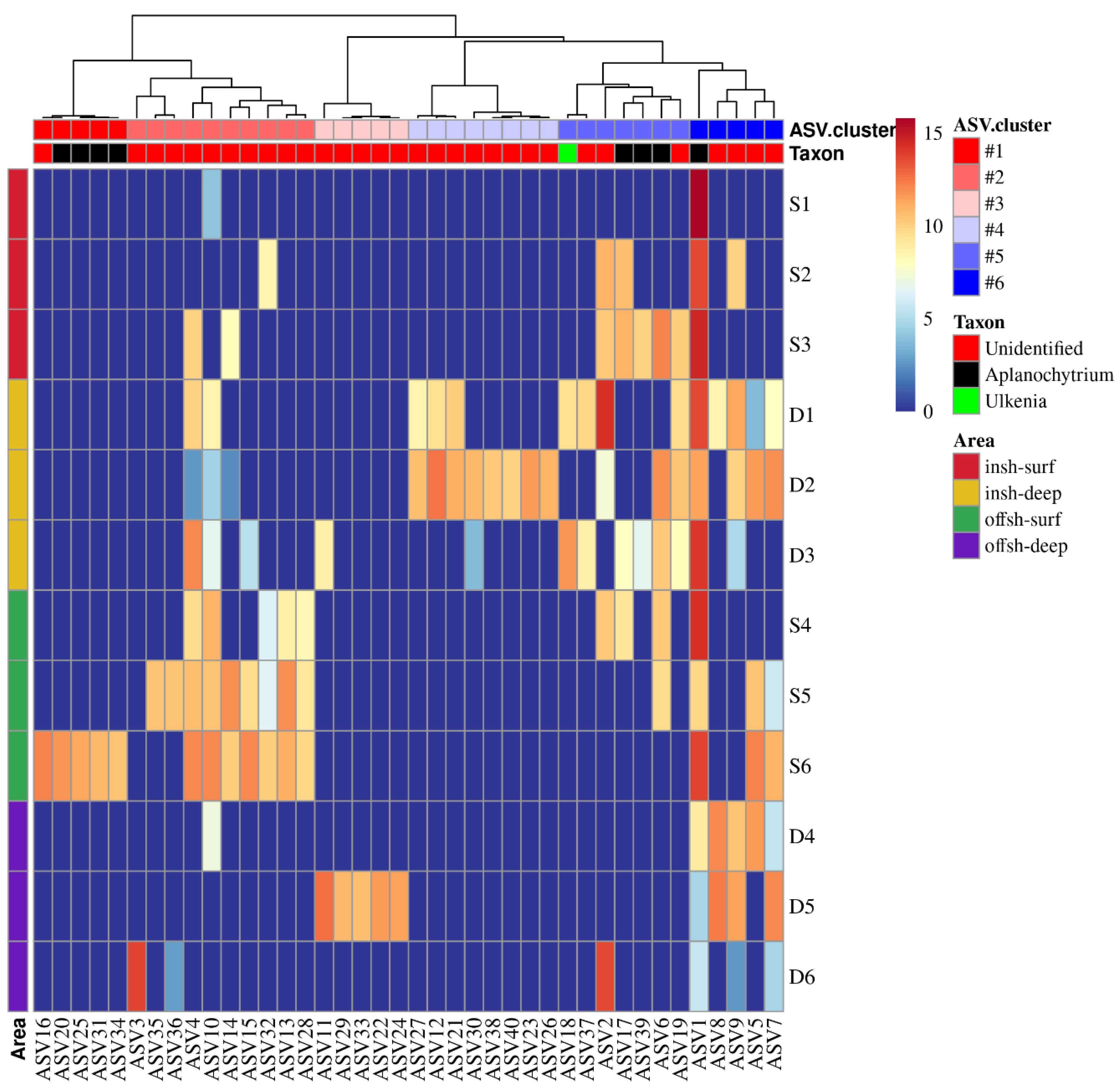
3.3. Community Structure of Labyrinthulomycetes Protists
3.4. Distribution Patterns of Dominant Labyrinthulomycetes Protists
3.5. Cooccurrence Network of Labyrinthulomycetes and Bacteria
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Caron, D.A.; Countway, P.D.; Jones, A.C.; Kim, D.Y.; Schnetzer, A. Marine protistan diversity. Ann. Rev. Mar. Sci. 2012, 4, 467–493. [Google Scholar] [CrossRef] [PubMed]
- Worden, A.Z.; Follows, M.J.; Giovannoni, S.J.; Wilken, S.; Zimmerman, A.E.; Keeling, P.J. Rethinking the marine carbon cycle: Factoring in the multifarious lifestyles of microbes. Science 2015, 347, 1257594. [Google Scholar] [CrossRef]
- Xie, N.; Sen, B.; Song, Z.; Zhao, Y.; Chen, Z.; Shi, W.; Zhang, Y.; Zhang, J.; Johnson, Z.I.; Wang, G. High phylogenetic diversity and abundance pattern of Labyrinthulomycete protists in the coastal waters of the Bohai Sea. Environ. Microbiol. 2018, 20, 3042–3056. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Singh, P.; Liang, Y.; Li, J.; Xie, N.; Song, Z.; Daroch, M.; Leng, K.; Johnson, Z.I.; Wang, G. Abundance and Molecular Diversity of Thraustochytrids in Coastal Waters of Southern China. FEMS Microbiol. Ecol. 2017, 93, fix070. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, X.; Liu, X.; Jiao, N.; Wang, G. Abundance and Novel Lineages of Thraustochytrids in Hawaiian Waters. Microb. Ecol. 2013, 66, 823. [Google Scholar] [CrossRef] [PubMed]
- Marchan, L.F.; Chang, K.J.L.; Nichols, P.D.; Mitchell, W.J.; Polglase, J.L.; Gutierrez, T. Taxonomy, ecology and biotechnological applications of thraustochytrids: A review. Biotechnol. Adv. 2018, 36, 26–46. [Google Scholar] [CrossRef]
- Nakai, R.; Naganuma, T. Diversity and Ecology of Thraustochytrid Protists in the Marine Environment. In Marine Protists: Diversity and Dynamics; Ohtsuka, S., Suzaki, T., Horiguchi, T., Suzuki, N., Not, F., Eds.; Springer: Tokyo, Japan, 2015; pp. 331–346. [Google Scholar]
- Singh, P.; Liu, Y.; Li, L.; Wang, G. Ecological dynamics and biotechnological implications of thraustochytrids from marine habitats. Appl. Microbiol. Biotechnol. 2014, 98, 5789–5805. [Google Scholar] [CrossRef] [PubMed]
- Taoka, Y.; Nagano, N.; Kai, H.; Hayashi, M. Degradation of Distillery Lees (Shochu kasu) by Cellulase-Producing Thraustochytrids. J. Oleo Sci. 2017, 66, 31–40. [Google Scholar] [CrossRef]
- Taoka, Y.; Nagano, N.; Okita, Y.; Izumida, H.; Sugimoto, S.; Hayashi, M. Extracellular Enzymes Produced by Marine Eukaryotes, Thraustochytrids. Biosci. Biotechnol. Biochem. 2009, 73, 180–182. [Google Scholar] [CrossRef]
- Nagano, N.; Matsui, S.; Kuramura, T.; Taoka, Y.; Honda, D.; Hayashi, M. The Distribution of Extracellular Cellulase Activity in Marine Eukaryotes, Thraustochytrids. Mar. Biotechnol. 2011, 13, 133–136. [Google Scholar] [CrossRef]
- Bongiorni, L.; Pusceddu, A.; Danovaro, R. Enzymatic activities of epiphytic and benthic thraustochytrids involved in organic matter degradation. Aquat. Microb. Ecol. 2005, 41, 299–305. [Google Scholar] [CrossRef]
- Damare, V.; Raghukumar, S. Morphology and Physiology of the Marine Straminipilan Fungi, the Aplanochytrids Isolated from the Equatorial Indian Ocean. Indian J. Mar. Sci. 2006, 35, 326–340. [Google Scholar]
- Takao, Y.; Tomaru, Y.; Nagasaki, K.; Sasakura, Y.; Yokoyama, R.; Honda, D. Fluorescence in situ hybridization using 18S rRNA-targeted probe for specific detection of thraustochytrids (Labyrinthulomycetes). Plankton Benthos Res. 2007, 2, 91–97. [Google Scholar] [CrossRef]
- Kimura, H.; Sato, M.; Sugiyama, C.; Naganuma, T. Coupling of thraustochytrids and POM, and of bacterio- and phytoplankton in a semi-enclosed coastal area: Implication for different substrate preference by the planktonic decomposers. Aquat. Microb. Ecol. 2001, 25, 293–300. [Google Scholar] [CrossRef]
- Ashley, K.E.; Crosta, X.; Etourneau, J.; Campagne, P.; Gilchrist, H.; Ibraheem, U.; Greene, S.E.; Schmidt, S.; Eley, Y.; Massé, G.; et al. Exploring the use of compound-specific carbon isotopes as a palaeoproductivity proxy off the coast of Adélie Land, East Antarctica. Biogeosciences 2021, 18, 5555–5571. [Google Scholar] [CrossRef]
- Liu, Y.; Singh, P.; Sun, Y.; Luan, S.; Wang, G. Culturable diversity and biochemical features of thraustochytrids from coastal waters of Southern China. Appl. Microbiol. Biotechnol. 2014, 98, 3241–3255. [Google Scholar] [CrossRef]
- Ueda, M.; Nomura, Y.; Doi, K.; Nakajima, M.; Honda, D. Seasonal dynamics of culturable thraustochytrids (Labyrinthulomycetes, Stramenopiles) in estuarine and coastal waters. Aquat. Microb. Ecol. 2015, 74, 187–204. [Google Scholar] [CrossRef]
- Bongiorni, L.; Mirto, S.; Pusceddu, A.; Danovaro, R. Response of benthic protozoa and thraustochytrid protists to fish farm impact in seagrass (Posidonia oceanica) and soft-bottom sediments. Microb. Ecol. 2005, 50, 268–276. [Google Scholar] [CrossRef]
- Xie, N.; Bai, M.; Liu, L.; Li, J.; He, Y.; Collier Jackie, L.; Hunt Dana, E.; Johnson Zackary, I.; Jiao, N.; Wang, G. Patchy Blooms and Multifarious Ecotypes of Labyrinthulomycetes Protists and Their Implication in Vertical Carbon Export in the Pelagic Eastern Indian Ocean. Microbiol. Spectrum 2022, 10, e00144-22. [Google Scholar] [CrossRef]
- Pan, J.; del Campo, J.; Keeling, P.J. Reference Tree and Environmental Sequence Diversity of Labyrinthulomycetes. J. Eukaryot. Microbiol. 2017, 64, 88–96. [Google Scholar] [CrossRef]
- Damare, V.; Raghukumar, S. Abundance of thraustochytrids and bacteria in the equatorial Indian Ocean, in relation to transparent exopolymeric particles (TEPs). FEMS Microbiol. Ecol. 2008, 65, 40–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Xie, N.; Bai, M.; Li, J.; Wang, G. Composition change and decreased diversity of microbial eukaryotes in the coastal upwelling waters of South China Sea. Sci. Total Environ. 2021, 795, 148892. [Google Scholar] [CrossRef] [PubMed]
- Collado-Mercado, E.; Radway, J.C.; Collier, J.L. Novel uncultivated labyrinthulomycetes revealed by 18S rDNA sequences from seawater and sediment samples. Aquat. Microb. Ecol. 2010, 58, 215–228. [Google Scholar] [CrossRef]
- Stokes, N.A.; Ragone Calvo, L.M.; Reece, K.S.; Burreson, E.M. Molecular diagnostics, field validation, and phylogenetic analysis of Quahog Parasite Unknown (QPX), a pathogen of the hard clam Mercenaria mercenaria. Dis. Aquat. Org. 2002, 52, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Murtagh, F.; Legendre, P. Ward’s Hierarchical Agglomerative Clustering Method: Which Algorithms Implement Ward’s Criterion? J. Classif. 2014, 31, 274–295. [Google Scholar] [CrossRef]
- Liu, X.; Xie, N.; Li, J.; Bai, M.; Sen, B.; Wang, G. Potential Contribution of Coastal Upwelling to Carbon Sink through Interaction between Cyanobacteria and Microbial Eukaryotes. Water 2022, 14, 3097. [Google Scholar] [CrossRef]
- Fortunato, C.S.; Herfort, L.; Zuber, P.; Baptista, A.M.; Crump, B.C. Spatial variability overwhelms seasonal patterns in bacterioplankton communities across a river to ocean gradient. ISME J. 2012, 6, 554–563. [Google Scholar] [CrossRef]
- Herlemann, D.P.R.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef]
- Bessudova, A.Y.; Sorokovikova, L.M.; Firsova, A.D.; Kuz’mina, A.Y.; Tomberg, I.V.; Likhoshway, Y.V. Changes in phytoplankton community composition along a salinity gradient from the lower Yenisei River to the Kara Sea, Russia. Bot. Mar. 2014, 57, 225–239. [Google Scholar] [CrossRef]
- Nielsen, D.L.; Brock, M.A.; Vogel, M.; Petrie, R. From fresh to saline: A comparison of zooplankton and plant communities developing under a gradient of salinity with communities developing under constant salinity levels. Mar. Freshw. Res. 2008, 59, 549–559. [Google Scholar] [CrossRef]
- Wang, S.; Sen, K.; He, Y.; Bai, M.; Wang, G. Riverine Inputs Impact the Diversity and Population Structure of Heterotrophic Fungus-like Protists and Bacterioplankton in the Coastal Waters of the South China Sea. Water 2022, 14, 1580. [Google Scholar] [CrossRef]
- Kimura, H.; Naganuma, T. Thraustochytrids: A neglected agent of the marine microbial food chain. Aquat. Ecosyst. Health Manag. 2001, 4, 13–18. [Google Scholar] [CrossRef]
- Bai, M.; Xie, N.; He, Y.; Li, J.; Collier, J.L.; Hunt, D.E.; Johnson, Z.I.; Jiao, N.; Wang, G. Vertical community patterns of Labyrinthulomycetes protists reveal their potential importance in the oceanic biological pump. Environ. Microbiol. 2022, 24, 1703–1713. [Google Scholar] [CrossRef]
- Xie, N.; Hunt, D.E.; Johnson, Z.I.; He, Y.; Wang, G. Annual Partitioning Patterns of Labyrinthulomycetes Protists Reveal Their Multifaceted Role in Marine Microbial Food Webs. Appl. Environ. Microbiol. 2021, 87, e01652-20. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, Y.; Honda, D. Nutritional intake of Aplanochytrium (Labyrinthulea, Stramenopiles) from living diatoms revealed by culture experiments suggesting the new prey-predator interactions in the grazing food web of the marine ecosystem. PLoS ONE 2019, 14, e0208941. [Google Scholar] [CrossRef]
- Raghukumar, S.; Damare, V.S. Increasing evidence for the important role of Labyrinthulomycetes in marine ecosystems. Bot. Mar. 2011, 54, 3–11. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Liu, X.; Xie, N.; Li, J.; Liu, X.; Sen, B.; Wang, G. Diversity and Community Composition of Labyrinthulomycetes Protists in the Coastal Zone of Hainan Island, South China Sea. Water 2023, 15, 738. https://doi.org/10.3390/w15040738
Song J, Liu X, Xie N, Li J, Liu X, Sen B, Wang G. Diversity and Community Composition of Labyrinthulomycetes Protists in the Coastal Zone of Hainan Island, South China Sea. Water. 2023; 15(4):738. https://doi.org/10.3390/w15040738
Chicago/Turabian StyleSong, Jing, Xiuping Liu, Ningdong Xie, Jiaqian Li, Xianhua Liu, Biswarup Sen, and Guangyi Wang. 2023. "Diversity and Community Composition of Labyrinthulomycetes Protists in the Coastal Zone of Hainan Island, South China Sea" Water 15, no. 4: 738. https://doi.org/10.3390/w15040738
APA StyleSong, J., Liu, X., Xie, N., Li, J., Liu, X., Sen, B., & Wang, G. (2023). Diversity and Community Composition of Labyrinthulomycetes Protists in the Coastal Zone of Hainan Island, South China Sea. Water, 15(4), 738. https://doi.org/10.3390/w15040738